
Abstract
Archaeologists have long envisioned direct encounters between Paleoindians and megafauna of the Last Glacial-Interglacial Transition (LGIT, 15–11.5 cal BP). Debate continues regarding the role that these Paleoindian hunters played in the extinction event(s). Archaeologists, paleontologists, and paleobiologists have proposed that Paleoindians proved to be very effective hunters who employed darts and spears tipped with razor-sharp, chipped-stone projectile points. These weapons are assumed to have been capable of inflicting mortal wounds and death as a result of massive blood loss. Few archaeologists, however, have considered the possible use of hunting poisons, as well as the implications of poison use for past procurement tactics and present-day archaeological research. This paper explores the feasibility of poison hunting by Paleoindians—specifically those derived from Aconitum spp. or monkshood—as well as the possible material correlates of this technology that might be observed in the archaeological record.
Introduction
Archaeologists have focused considerable attention on the weapon technology of Clovis hunters since the early discoveries of carefully flaked stone points in direct association with the remains of now extinct megafauna. The Clovis period is thought to have had a maximum time span of 450 years (13,250–12,800 cal BP) and a minimum span of 200 years (13,125–12,925 cal BP) (Fiedel 2015:14; Waters and Stafford 2007). Frison (2004:43) has presented some of the most thorough discussions of Clovis period hunting weaponry; he stated that “Clovis hunters possessed well-designed, well-made, and well-maintained weapons and tools, and developed systematic and effective large mammal procurement strategies.” In keeping with Frison's extensive research, many archaeologists, paleoecologists, and others assume that the deaths of targeted prey animals resulted primarily from mortal wounds and blood loss. They, too, assume that Clovis projectile points themselves were intrinsically lethal or “weapons of mass destruction” (e.g., Collins 2007; Fiedel and Haynes 2004; Frison 1989, 1991, 1998; Frison and Bradley 1999; Gorman 1972; Haynes 1966; Haynes 1991, 1999a, 1999b, 2002; Lister and Bahn 1994; Pearson 2001; Storck 2004; Tankersley 2002). It is important to point out that these chipped stone weapon points are quite small. Clovis points associated with mammoth (Mammuthus primigenius) kills (e.g., Blackwater Draw, Lehner 1 and 2, Miami, and Naco) range from 3–13 cm in length and from 2.5–3.5 cm in maximum width (Gorman 1972; Haury et al. 1953, 1959).
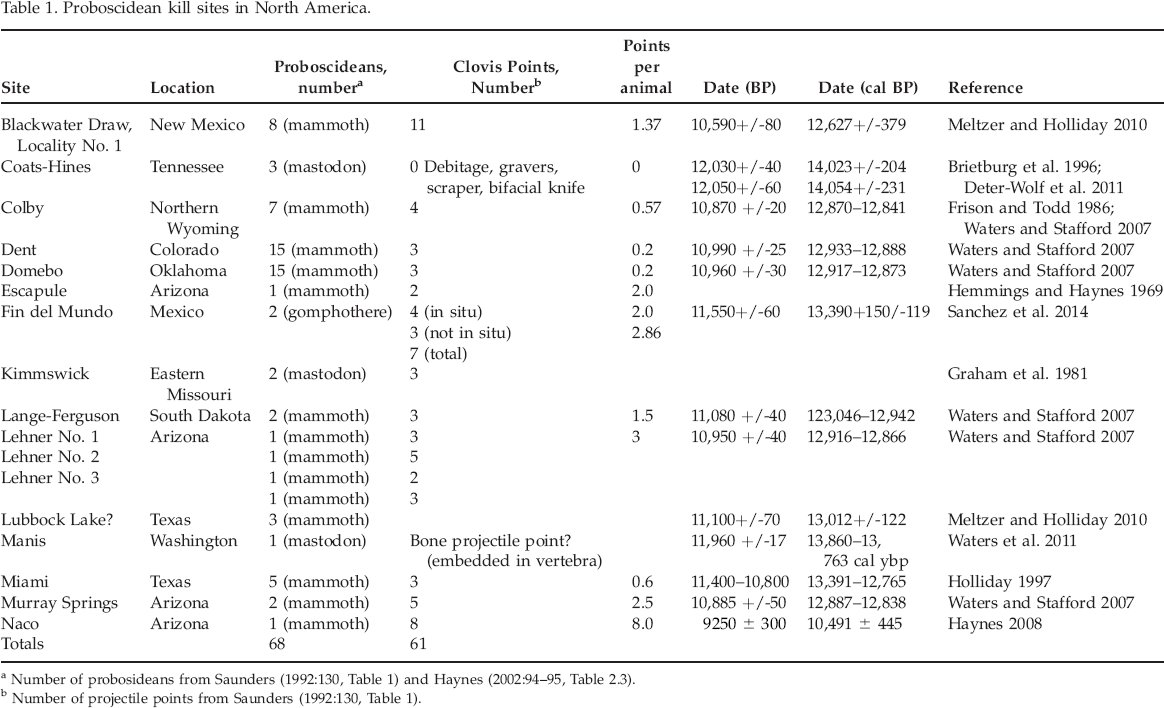
Archaeologists have arrived at several different estimates of the number of Clovis-age proboscidean kill sites in North America. Grayson and Metzler (2002, 2003) concluded that there are 14 Clovis proboscidean kill locations that include 15 Clovis components. Other archaeologists have use of less conservative criteria and concluded that there are 26 Clovis mammoth and mastodon (Mammut americanum) kill sites (Surovell and Waguespack 2008; Waguespack and Surovell 2003). In both cases, archaeologists consider that the most significant criteria include the direct association of projectile points (principally chipped stone points) and evidence for butchering (dismemberment and cut marks on bone). At present, there are 15 Clovis-age proboscidean kill sites (Table 1) that have been investigated in North America and northern Mexico (Grayson and Meltzer 2003, 2015; Sanchez et al. 2014; Surovell and Waguespack 2008). The Coats-Hines mastodon kill site in central Tennessee and Fin del Mundo in Mexico have been added to this list. Sixty-eight proboscideans are represented at these sites and a total of 61 Clovis projectile points were found associated with these remains. The number of projectile points associated with proboscideans varies from less than one to eight per animal. These proboscidean kills were carried out within a 200–450 year interval within the Last Glacial-Interglacial Transition (LGIT; 15,000–11,500 BP) (Haynes 2002; Haynes and Eiselt 2004; Surovell and Waguespack 2008). A number of archaeologists attribute the extinction of 17 to 35 genera of Late Pleistocene mammals to early Paleoindians—particularly the so-called Clovis people (e.g., Fiedel 2009:21; Waguespack 2014; cf. Grayson and Meltzer 2015; Meltzer 2015).
Proboscidean kill sites in North America.
Archaeologists continue to assume that Paleoindians hunted megamammals solely with spears and darts tipped with chipped-stone points, in part because several field experiments have demonstrated that replicated atlatl darts tipped with chipped-stone points or fire-hardened wood points can successfully penetrate the carcasses of African elephants (Loxodonta africana) (Butler 1980; Frison 1989; Huckell 1982). Also, their assumptions about the efficacy of early Paleoindian hunting weaponry appear to be bolstered by historic accounts of elephant hunting in Africa. It is suggested here, however, that such descriptions of historic elephant hunters and their weapons should be examined further.
The purpose of this paper is to explore the feasibility of an alternative hunting technology that could have made use of plant-based poisons during the LGIT between 15,000–11,500 years ago. Discussions of early Paleoindian hunting weapon systems to date have given very little attention to the possible use of poisons. There are three significant questions that must be asked at this point regarding the feasibility of the use of hunting poisons by Paleoindians. First, were there suitable poisons available that possessed the potential for killing megamammals? Second, how might archaeologists determine the lethal dosage of such a poison? And, third, would it have been possible to deliver a lethal dose of poison with Paleoindian hunting weapons (e.g., thrusting spears, throwing spears, or atlatl darts)? Once effective hunting poisons have been identified, it is necessary to make use of physiological and pharmacological concepts to calculate the lethal dosages required to kill Columbian (Mammuthus columbi) and woolly mammoths (Mammuthus primigenius), American mastodons (Mammut americanum), and gomphotheres (Cuvieronius sp.). Initially, however, it is helpful to briefly review the ethnographic record regarding elephant hunting in Africa and the cross-cultural use of hunting poisons, which highlights how elephants have been hunted and the feasibility of poison use.
Elephant Hunting in the Ethnographic Record
Historical accounts of elephant hunting by indigenous groups in Africa describe hunting tactics that employed thrusting spears, throwing spears, bow and arrows, and harpoon down-falls (Lagercrantz 1934; Macphail 1930). Poison hunting of elephants will be discussed in detail later. Bandala hunters in southern Sudan worked cooperatively to chase and hamstring bull elephants (Macphail 1930). Hunters dispatched bull elephants with thrusting spears once the hind legs had been disabled (Macphail 1930:281). Nuer hunters of Sudan attacked elephants with throwing spears (Howell 1945).
The Mbuti Pygmies of the Ituri Forest relied upon two primary tactics for killing elephants. First, the net hunters of the Epulu District of the Central Ituri used a spear with an attached cord as a harpoon. This device inflicted multiple injuries upon the elephant as it ran through the forest and the attached cord caught on trees and undergrowth (Schebesta 1941). Second, the Mbuti are well-known for their daring and perilous methods for elephant hunting. Hunters would silently stalk elephants as they traveled through the dense forest. In such cases, the hunters may have covered themselves with elephant dung before they approached the animal. They would then crawl beneath the elephant and plunge a razor-sharp, 30–50 cm long iron spear point and a wooden shaft (ca. 75 cm long) deep into its stomach just behind the rib cage (Janmart 1952:146–147; Putnam 1948:331). The hunter then stood motionless in the nearby bush to avoid detection. According to such accounts, the wounded animal would flee and later succumb to peritonitis within two to three days (Putnam 1948:331). Pygmies also resorted to hamstringing elephants. Once they located the dead elephant, pygmy groups would establish a short-term camp near the carcass where they remained until all of the meat was consumed (Turnbull 1968:138).
It should be pointed out that historic elephant hunting employed thrusting and throwing spears, as well as knives and axes (for hamstringing elephants) made from iron (Carrington 1959; Laub 1992:106). The iron spear points were two and one-half to four times as long as most prehistoric Clovis chipped-stone points, exhibited broad blades with razor-sharp cutting edges, and did not fracture or break while in use. There is little comparison, therefore, between historic iron projectile points and those fashioned by Paleoindians. As Saunders (1992:129) states, “In short, modern practices of elephant procurement are an impoverished analogy for an understanding of how a Pleistocene lifeway or phenomenon was enacted.” Thus, poisons may have been critical for hunting large mammals with stone-tipped projectiles.
Poison Hunting and the Ethnographic Record
Hunting poisons have been utilized in Africa, India, Southeast Asia, China, Japan, Melanesia, and the Americas (Bisset 1966, 1976, 1979, 1981, 1992; Bisset and Woods 1966; Cheney 1926; Hoffman 1891; Jett 1970; Jones 2007; Lewin 1923; Mines 1908; Neuwinger 1996, 1998; Perrot and Vogt 1913; Rambo 1978; Schapera 1925; Schebesta 1941; Woodburn 1970). Hunting poisons were used at least as early as 24,000 years ago at Border Cave in South Africa (d'Errico et al. 2012). This poison was the protein, ricin, derived from castor beans (Rincinus communis) (d'Errico et al. 2012:13218; cf. Evans 2012). Old World hunting poisons are typically plant-based, fast-acting cardiotoxins. Most poisons are derived from phytotoxins including strychnine (Strychnos sp.), ouabain (Acokanthera sp.), pro-inflammatory diterpenes (Hippomane sp., Euphorbia sp., and Sapium sp.), and aconitine (Aconitum sp.).
Central and South America hunters (e.g., Choco, Cuna, Embera, and Naonama) applied batrachotoxin from the skin of tropical frogs (e.g., Phyllobates terribilis) to poison the tips of blowgun darts (Alto 2011; Patocka et al. 1999; Saporito et al. 2007). In tropical South America, poisons containing muscle relaxants like tubocurarine or curarine were used to hunt birds, monkeys, and sloths in the canopy, as well as peccaries on the forest floor (Bisset 1992). The San and G/wi Bushmen of the Kalahari Desert in present-day Botswana and Namibia were able to kill gemsbok (Orynx sp.) and hartebeest (Alcelaphus sp.) with short, unfletched arrows propelled by small, simple bows (Lee 1979; Silberbauer 1981). Bushmen arrows were tipped with small metal points and the foreshafts were coated with poison derived from the larvae of beetles (Diamphidia sp. and Blepharida sp.), the beetle Polyclada flexuosa, and a number of phytotoxins (Koch 1958; Shaw et al. 1963; Woollard 1986).
Heizer (1938, 1943) published several of the earliest anthropological studies of hunting poison used in northern temperate and subarctic environments to kill seals, sea lions, walruses, and whales. Such phytotoxins were derived from wolfsbane (Aconitum napellus), larkspur (A. delphinifolium), and A. maximum (Heizer 1943:443–445). The Ainu of Japan used arrows treated with the same poison(s) to hunt bear (Ursus arctos), Sika deer (Cervus nippon), and sea mammals (Irimoto 2002; Ishikawa 1962; von Siebold 1878).
Travelers' accounts and ethnographic studies from Africa and India have described various poisons used for hunting elephants (Frazer 1889; Frazer and Tillie 1899; Holman 1967; Parker and Amin 1983). Laidlaw (1909) described the use of spears by Bini (Edo) hunters in southern Nigeria to hunt elephants. The spears had barbed iron points coated with strophanthin poison. Frazer (1889) and Frazer and Tillie (1899) provided an early description of the Wakamba and Wanyika who hunted elephants in the Tsavo and Taita highlands of Kenya with bow and arrows. The Liangulu (Wasanya, Waliangulu, or Waata), who lived in the Tsavo highlands, also hunted elephants with extremely powerful long bows and metal-tipped poison arrows (Holman 1967; Parker and Amin 1983). The elephant hunting bows were made from heartwood staves more than five feet long (1.52 m) and had draw weights that ranged between 120–170 lbs (54.4–77.1 kg) (Parker and Amin 1983:34).
The Liangulu boiled the bark, roots, and wood of the Acokanthera tree to form a thick, dark residue that contained ouabain, a cardiac glycoside (Neuwinger 1996:74–80; Parker and Amin 1983:36). They then applied 8 to 15 grams of this paste to the arrow foreshafts (Laidlaw 1909:354; Parker and Amin 1983:39). Such amounts were more than 70 times the lethal dosage for an elephant weighing 5,443 kg (12,000 lbs; Parker and Amin 1983:30). Neuwinger (1996:78) states that “Acokanthera arrow poisons, kept in a dry place, will keep their effect for decades.” Hunters then used their powerful bows and poisoned arrows to target the gut (small intestines) and also the base of the ear, not the heart and lungs, so that the poison spread via intraperitoneal and intravenous routes through the elephant's body (Holman 1967:40). Holman (1967:87) states, “Well shot with good poison, an elephant might collapse after running two hundred yards….” Larger prey like eland (Taurotragus oryx; 837 kg), gemsbock (Oryx gazelle; 145 kg) and elephant (10,000 kg) died more quickly than smaller animals like zebra (Equus zebra; 234 kg) and warthog (Phachochoerus aethiopicus; 28 kg).
Aconitine—A Possible Paleoindian Hunting Poison
Few archaeologists and anthropologists have mentioned the use of poisons for hunting by Paleoindians in the Americas (cf. Haury et al. 1959; Haynes 2002; Jones 2007; Wedel 1986). Interestingly, Haury et al. (1959:29) briefly mentioned the possible use of hunting poisons in their description of the Clovis-age Lehner mammoth kill site in southeastern Arizona. Wedel (1986:59) suggested that the earliest Americans may have made use of poisons like water hemlock (Cicuta sp.), prairie larkspur (Delphinium sp.), monkshood (Aconitum sp.), nightshade (Solanum sp.), and pokeberry (Phytolacca sp.) to enhance the effectiveness of thrown darts or spears used to debilitate or kill megafauna. North American archaeologists have not seriously considered hunting poisons perhaps because they associate their use with tropical and sub-tropical environments found in Africa, India, Asia, as well as Central and South America.
The plant poisons used by the Wakamba and the Liangulu in the highlands of southern Kenya to kill elephants were not available to Paleoindian populations in North America. Aconite poison, however, was available throughout North America as well as Asia and Europe (Bisset 1976; Heizer 1938, 1943). In fact, aconite derived from various species of Aconitum was used extensively by many indigenous groups as a hunting poison throughout northwestern India, Tibet, Burma, China, Eastern Russia, Sakhalin, Japan, Siberia, the Aleutians Islands, southwestern Alaska, and Kodiak Island (Bisset 1976; Heizer 1938, 1943). Consequently, aconite is a good candidate for a hunting poison possibly used by hunters in Asia during the Late Glacial Period and later in North America during the LGIT.
In North America, Brink (1982:14) states that Aconitum is distributed throughout circumboreal arctic and alpine environments and in high altitude mesic habitats of mountain ranges that stretch from British Columbia to Mexico. In western North America (Brink 1982:21), Aconitum species are confined to isolated mountain ranges or moisture islands between 1,219–3,659 m (4,000–12,000 ft) above mean sea level. Brink (1982:21) states, “During glacial periods the climate of this area was probably more suitable than it is now for arctic and alpine plants and Aconitum probably enjoyed a wider distribution at low elevations.”
Aconite and its constituent alkaloids are among the most thoroughly investigated poisons, as well as pharmaceuticals, in the world (e.g., Allen 1929; Cash and Dunstan 1903; Fatovich 1992; Fujita et al. 2007; Ishikawa 1962; Munch 1938; Murayama et al. 1991; Puri 1974; Srivastava 2010; Swanson 1938). Aconite poison is derived from Aconitum species that contain highly toxic alkaloids including hypaconitine, masaconitine, jesaconitine, and aconitine (Bisset 1976, 1979, 1981; Husa 1938; Stern 1954). Aconitine is the most significant toxic alkaloid in aconite poison and it acts as both a neurotoxin and a cardiotoxin (Bisset 1981:296; Mosto and de Landoni 1990). One gram of fresh Aconitum root contains 2 to 20 mg aconite (Mofenson and Caraccio 2001). Unlike many African hunting poisons, poisons made from Aconitum do not have to be boiled. Once suitable plants were found, the preparation of poisons did not require a great deal of time or complex technology (Bisset 1976, 1979). The Chinese crushed Aconitum roots and allowed the extracted juice to evaporate from bowls for seven days to form a crystalline alkaloid (probably aconitine) that looked like brown sugar. The Ainu combined aconite with toxic marsh marigold (Caltha palustris) and mugwort (Artemesia sp.) to produce surku, a very toxic arrow poison that could be stored for five months (Bisset 1976:90–91; Heizer 1938:361).
Aconite was applied to arrow foreshafts, as well as the projectile points themselves. The Ainu of Japan used arrows with bamboo points dipped in aconite (wolfsbane) and coated with pine resin (Irimoto 2002; Von Siebold 1878; Watanabe 1972:31, 37). The Hokkaido and Sakhalin Ainu placed thick aconite paste along the concave side of a bamboo point and then sealed it with pine resin (Bisset 1976:94). After the paste was applied, they dipped the arrow points in pine gum (Bisset 1976:98). Arrow foreshafts were also wrapped with five centimeters of cotton thread, soaked in aconite, and coated with carbon black produced by smudge fires fueled with straw (Bisset 1979:337).
Calculating Lethal Dosages for Extant Mammals
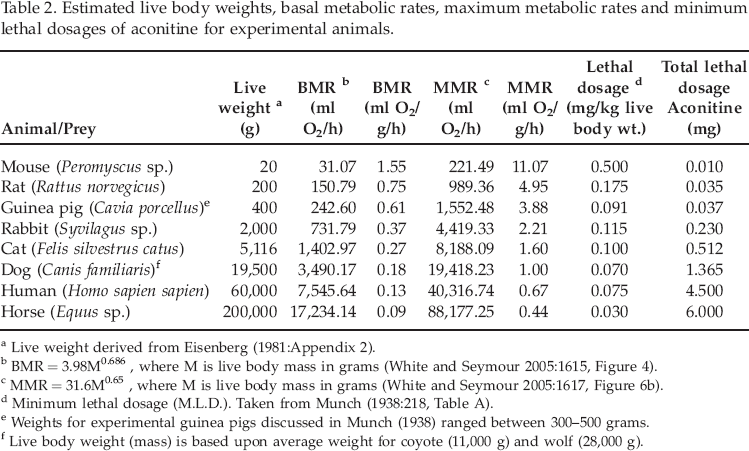
Early research has determined the toxicological properties of the constituent alkaloids of aconite through use of laboratory mice, rats, guinea pigs, rabbits, cats, dogs, and horses (e.g., Cash and Dunstan 1903; Mosto and de Landoni 1990; Munch 1938). Heizer (1943) used data from these early laboratory investigations in his study of aconite hunting poison. Using a simple proportionality based upon lethal dosage and body weight, he calculated an average lethal subcutaneous dosage (LD50) of aconite equal to 0.13 mg/kg live body weight for mammals (Heizer 1943:440). Heizer (1943) then utilized this average lethal dosage to calculate lethal dosages for marine mammals including seals, walruses, sea lions, and whales.
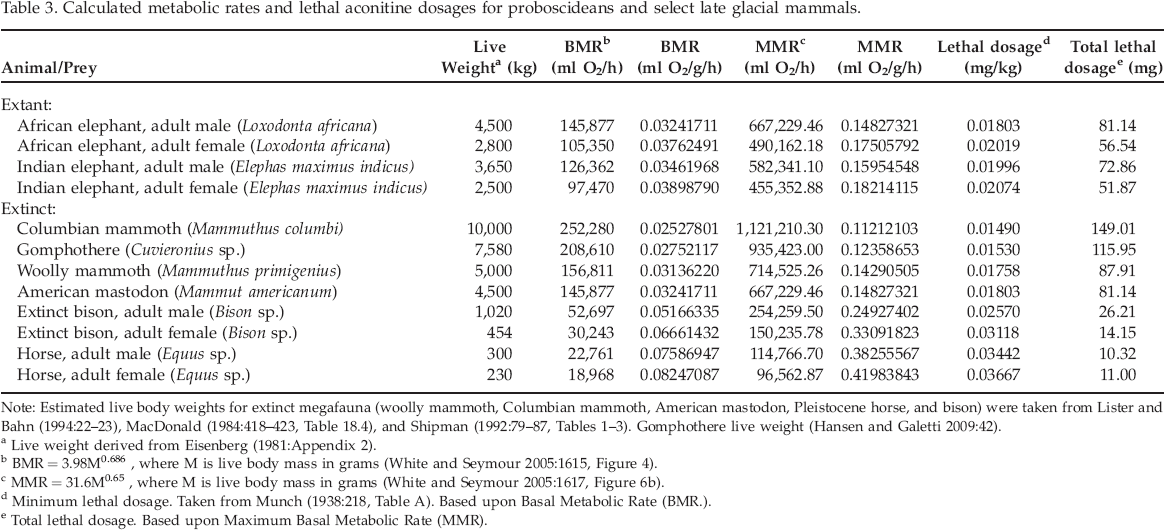
Heizer's (1938, 1943) calculations are incorrect because they do not consider the interrelationships between animal body size, basal metabolic rate, and detoxification capabilities (Norris 1998; Schmidt-Nielsen 1984). His calculation does not take into consideration toxicokinetics and toxicodynamics that examine how the body processes toxins as well as how toxins affect the body, respectively (Tozer and Rowland 2007; Travis et al. 1990:285). Chemicals introduced into the body are generally metabolized at different rates as a function of live body weight or mass. For mammals, then, basal metabolic rate is, in general, inversely related to body size (Kleiber 1932; McNab 1988; White and Seymour 2003, 2005). Resting or basal metabolic rate for mammals can be calculated using the equations provided by White and Seymour (2005:1615, 1617). Average live body weights were based upon data provided by Eisenberg (1981:464–471, Appendix 2). The equation for calculating basal metabolic rate of mammals is: (BMR) = 3.98 (Mb) 0.686+/-0.014; where BMR is expressed as milliliters of oxygen per hour (ml O2/h) and body mass Mb is measured in grams (g). If mammals are cold stressed, as might be expected for mammoths during severe winters, the maximum metabolic rate (MMR) can be calculated using the following equation: MMRc = 31.6Mb0.65+/-0.05.
Data derived from the previous investigations of aconitine toxicity by Cash and Dunstan (1903) and Munch (1938) were used to calculate the relationship(s) between lethal dosages of aconitine and metabolic rates of select prey animals. In some cases, Cash and Dunstan (1903) provided live body weights and corresponding lethal dosages for aconite and aconitine. Both BMR and MMR are expressed as milliliters of oxygen used per hour (ml O2/h), as well as milliliters of oxygen utilized per unit live body weight in grams (ml O2/g/h) were calculated for the laboratory animals using the equations provided by White and Seymour (2005) (see Table 2). In turn, two measures of lethal dosage (mg/kg live body weight) and total lethal dosage (mg/total live weight) were also calculated based on the laboratory data (Table 2). The relationship (y = −0.000412 + 0.294x) between lethal dosage (y in mg/kg live body weight) and basal metabolic rate (BMR) is shown in Figure 1. The relationship (y = 19.6x −2.46) between total lethal dosage (mg) and basal metabolic rate (BMR) for the experimental animals is presented in Figure 2.
Estimated live body weights, basal metabolic rates, maximum metabolic rates and minimum lethal dosages of aconitine for experimental animals.
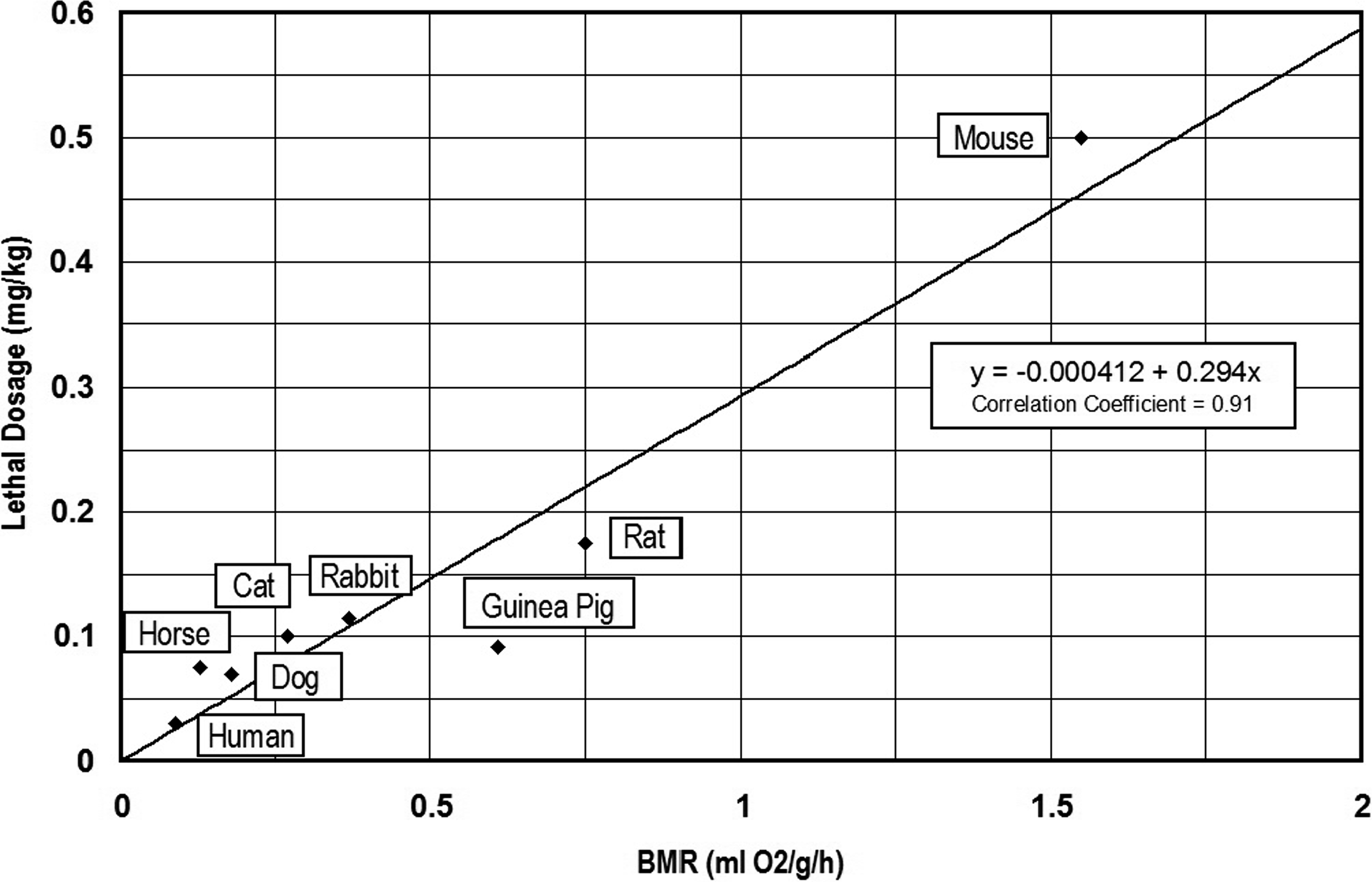
Relationship between basal metabolic rate (BMR) and lethal dosage (mg/kg) of aconitine for experimental animals.
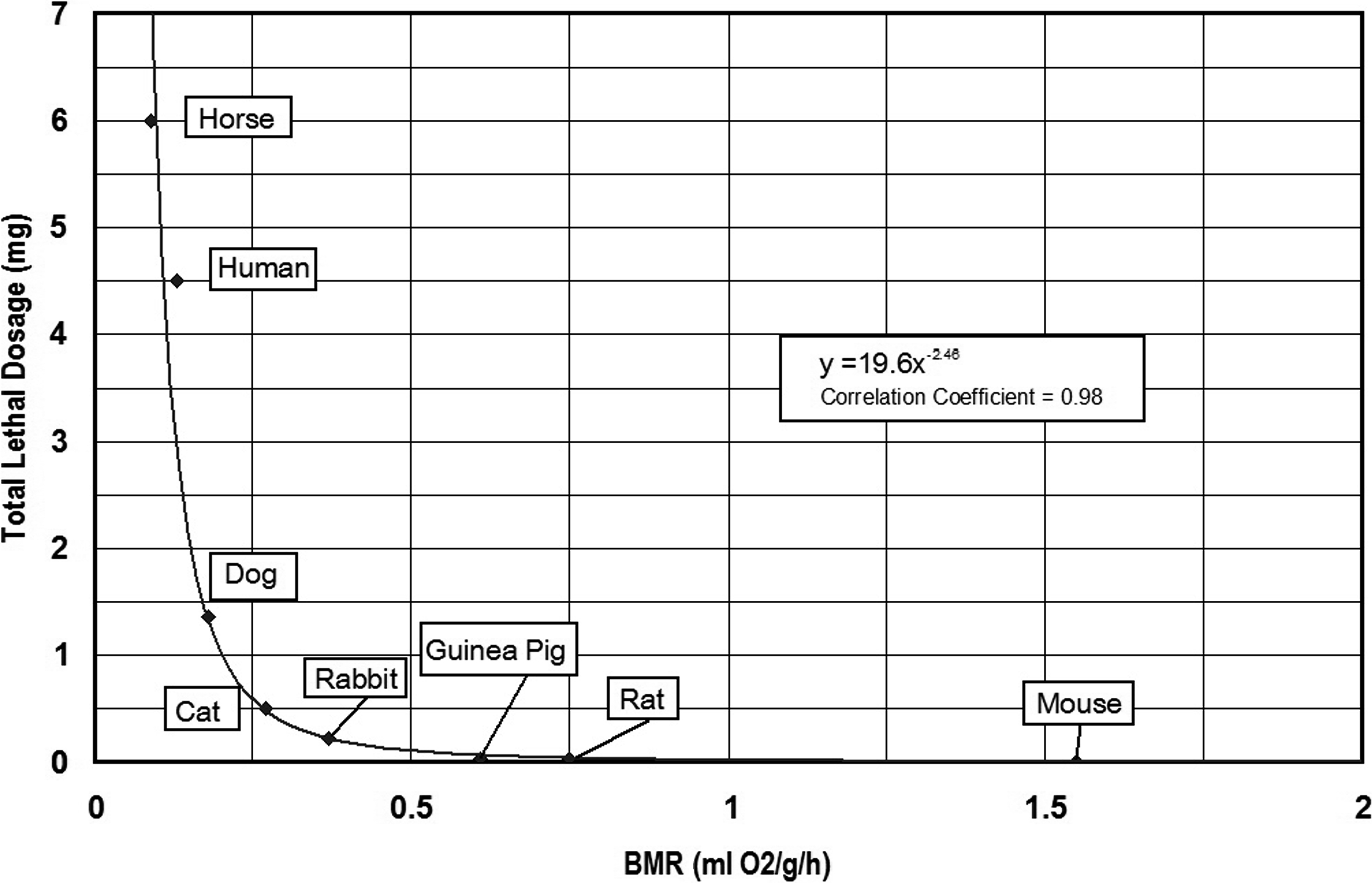
Relationship between basal metabolic rate (BMR) and total lethal dosage (mg) of aconitine for experimental animals.
Calculating Lethal Dosages for Proboscideans
In order to determine the lethal dosages of poison required to kill now extinct proboscideans, it is necessary to determine resting or basal metabolic rates. Estimates of live body weights for mammoths, mastodons, gomphotheres, and other possible LGIT prey are based upon a number of sources (Christiansen 2004; Klein 1969:222; Lister and Bahn 1994:22–23; MacDonald 1984:418–423, Table 18.4; Shipman 1992:79–87, Tables 1–3). Morphological characteristics of extinct proboscideans are important to consider when thinking about hunting tactics and weaponry. Columbian mammoths stood ca. 3.2–3.5 m high at the shoulders, whereas woolly mammoth and mastodon exhibited shoulder heights of 3–3.5 m and 2.75–3 m, respectively (Christiansen 2004). Shipman (1992:79–81) demonstrates that estimated live weights for mammoth vary considerably based upon allometric calculations using humerus length versus least midshaft circumference because their short, stocky bodies probably reflect their adaptations to very cold winters (Shipman 1992:84–85).
Calculated metabolic rates and lethal aconitine dosages for proboscideans and select late glacial mammals.
I calculated the BMR and MMR for these animals (Table 3) using equations provided by White and Seymour (2005:1615,1617). Estimated basal metabolic rates were used to calculate lethal dosages (mg/g live body wt) and total lethal dosages (mg/live body wt) (Table 3; Figures 3 and 4) for both extant and extinct proboscideans as well as bison (Bison sp.) and horse (Equus sp.). Lethal dosages of aconitine were then calculated for extant African and Indian elephants (Elephas maximus indicus), as well as Columbian mammoth (Mammuthus columbi), woolly mammoth (Mammuthus primigenius), American mastodon (Mammut americanum), and gomphothere (Cuvieronius) using the regression equations based upon data from previous animal experiments. Estimated total lethal dosages required to kill proboscideans both modern and extinct, range from ca. 52 mg to 149 mg aconitine. An adult female Indian elephant and the American mastodon weighing 2,500 kg would require an equivalent lethal dosage of 81 mg aconitine (Table 3). A woolly mammoth weighing 5,000 kg would require 88 mg of aconitine if its metabolic rate was near the resting or basal level during the warm season. In this case, one might visualize an 88 mg total lethal dose of hunting poison as equivalent to 293 grains of salt or sodium chloride (ca. 0.3mg/grain). Using Heizer's subcutaneous lethal dosage of aconite (0.13 mg/kg live body weight), the lethal dosage for a 5,000 kg mammoth would equal 650 mg, which is more than seven times the lethal dosage based upon Heizer's approach. The lethal dosage of aconitine necessary for an adult Columbian mammoth weighing 10,000 kg would equal ca. 149 mg. Other prey animals such as adult Pleistocene horses (Equus sp.; female and male) could be dispatched with 10–11 mg of aconitine and adult bison (female and male) would require lethal dosages of 14 mg and 26 mg, respectively.
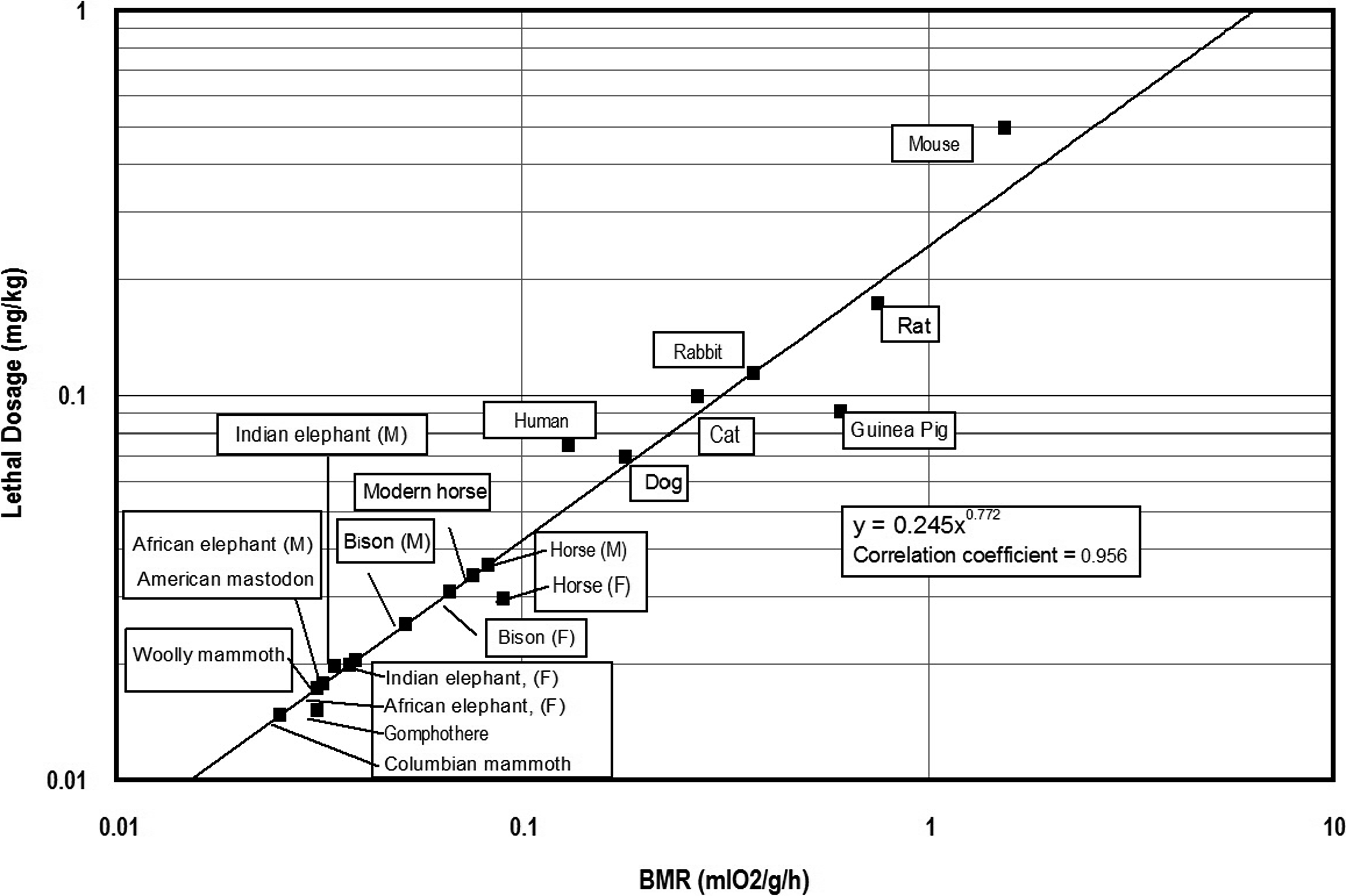
Relationship between basal metabolic rate (BMR) and lethal dosage (mg/kg) of aconitine for experimental animals, extinct and extant proboscideans, and select extinct mammals.
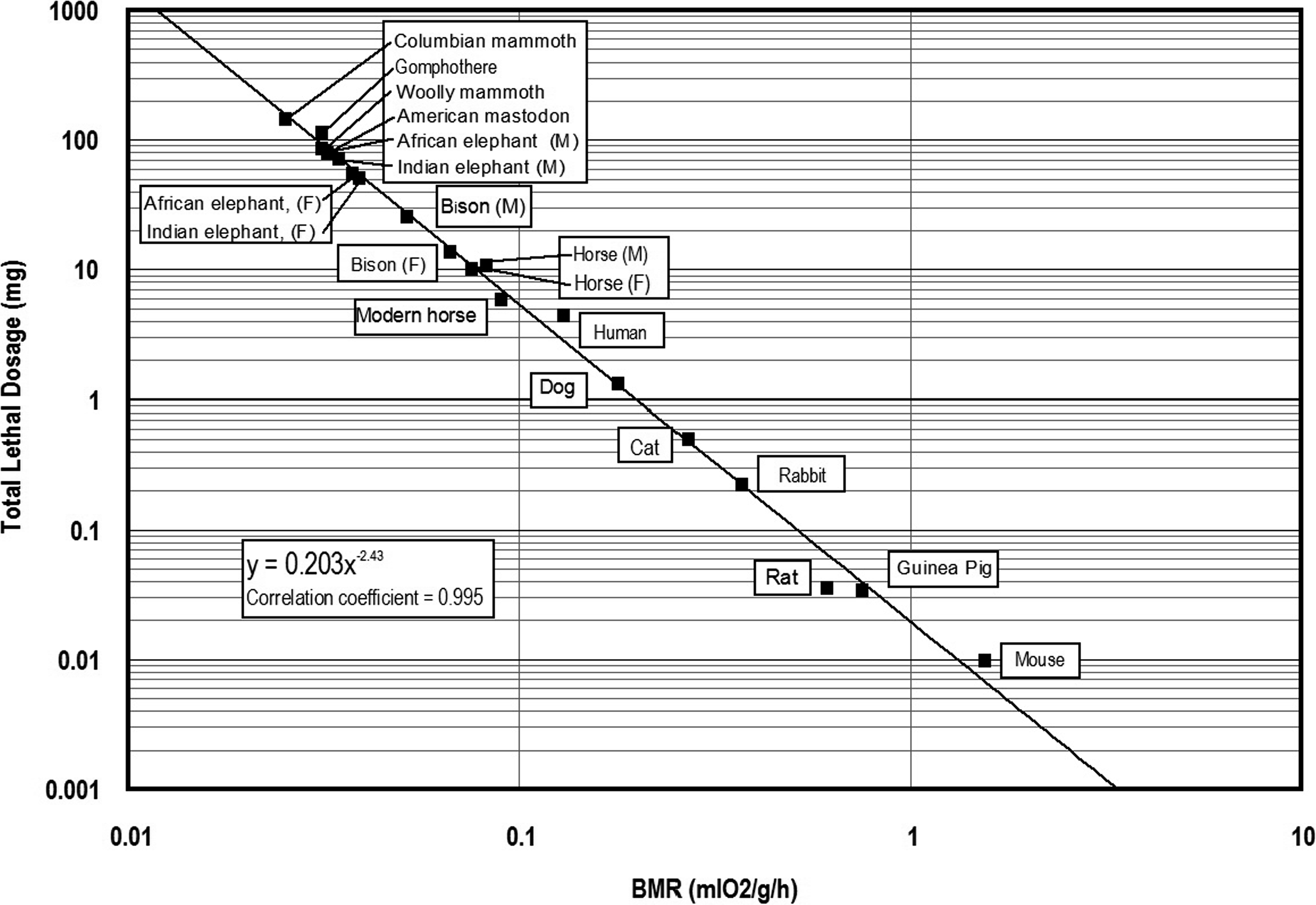
Relationship between basal metabolic rate (BMR) and total lethal dosage (mg) of aconitine for experimental animals, extinct and extant proboscideans, and select extinct mammals.
It is interesting to note that lethal dosages of hunting poisons may vary seasonally as a function of air temperature. Severe winter climatic conditions during the LGIT would have affected mammalian metabolism. Cold-induced maximum metabolic rate (MMRc) can be calculated using the equation: MMRc = 31.6Mb0.65+/-0.05, where Mb equals body mass in grams (see White and Seymour 2005:1617; Figure 6b). The MMRc for the adult woolly mammoth is more than 4.5 times its basal metabolic rate (BMR). Consequently, elevated metabolic rates as a function of cold stress would have necessitated higher lethal dosages of aconitine (474 mg) when mammoths were hunted during the winter. In this case, a lethal dosage of 474 mg might be visualized as equivalent to 1,520 grains of salt or sodium chloride. One might expect, then, to observe lethal dosages of hunting poison to vary in environments characterized by marked seasonal fluctuations in air temperature as well as wind chill factors.
Discussion
Archaeologists have generally assumed that Early Paleoindian hunters directly confronted and killed megamammals using stone-tipped darts and spears. Death, in such cases, is presumed to have resulted from massive blood loss 1 . As mentioned, archaeologists have demonstrated in a series of field experiments that atlatl darts with fire-hardened tips or chipped stone points can penetrate the hides and flesh of modern elephant carcasses (Butler 1980; Frison 1989; Huckell 1982). Early Paleoindian weaponry including fluted chipped stone projectile points and foreshaft assemblies would have been sufficient to have introduced lethal doses of phytotoxins such as aconitine. Possible configurations of Paleoindian hunting weapons and possible applications of poison are provided in Figure 5. The chipped stone points (i.e., Clovis, Cumberland, and Folsom) exhibit channels or flutes on both faces that facilitated the insertion of freshly sharpened tips into a slotted foreshaft. Additionally, hunting poison could be applied to the concavities in the point as well as to the foreshaft. Ethnographic accounts for the Waata elephant hunters of Kenya and others suggest that most of the poison, in fact, would have been applied to the foreshafts of complex weapons.
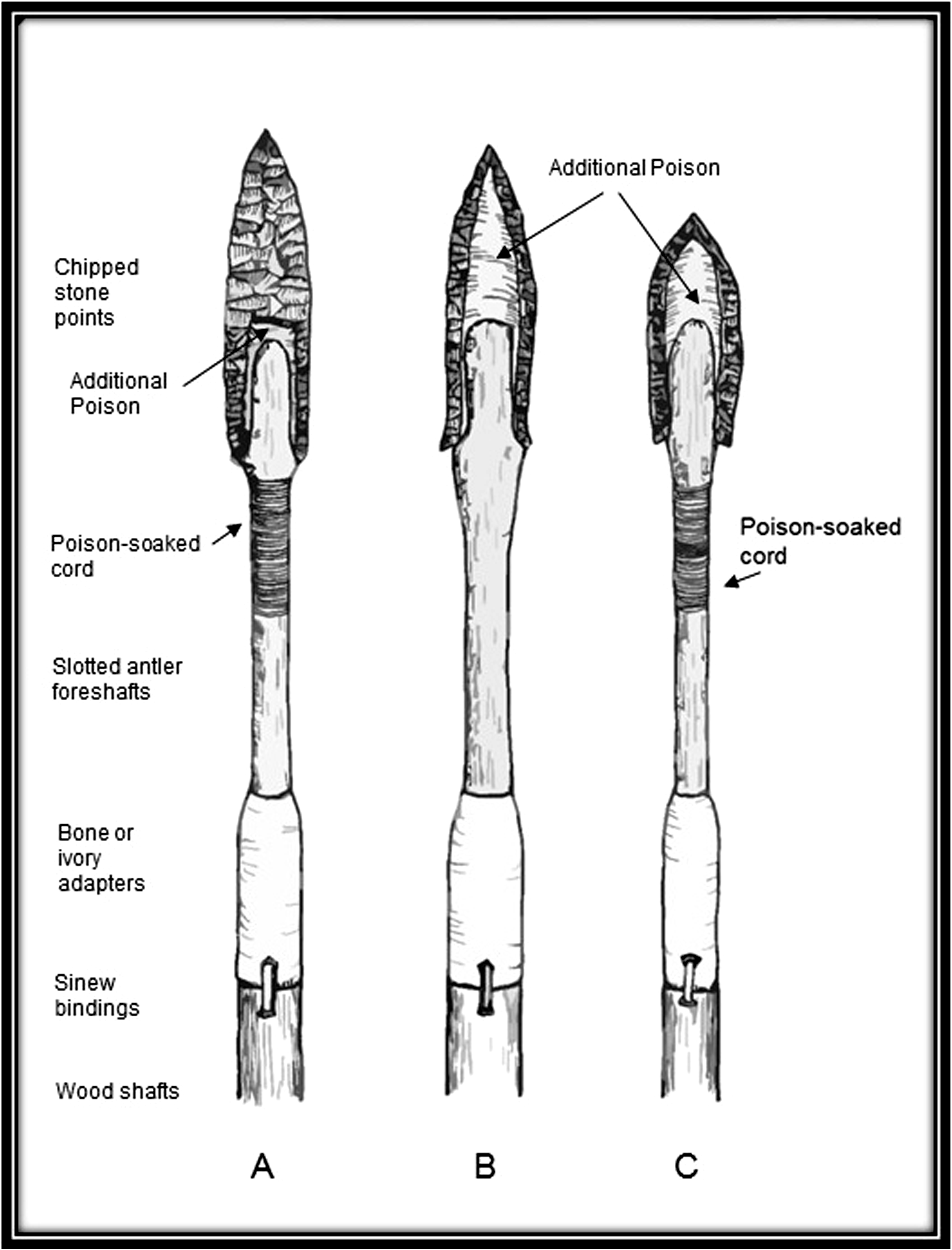
Possible hafting configurations and poison applications for Paleoindian chipped stone projectile points (A, Clovis; B, Cumberland; C, Folsom).
Bifacial, chipped stone weapon points have been found in direct association with proboscidean remains but archaeologists have yet to discover these projectile points actually embedded in bones 2 . On the other hand, there are sites that contain megafauna remains and chipped stone points that exhibit impact damage. For example, Clovis points were found directly associated with bison remains (Bison antiquus - B. occidentalis) at Murray Springs in Arizona (Hemmings 2007). Excavations in Area 4 yielded two complete Clovis points, four Clovis points with distal impact fractures, three Clovis points with one or more ears missing, and three points that exhibited transverse snaps (Hemmings 2007:123). Photographs of the Clovis points associated with mammoth remains recovered from both the Naco site (n = 8; one point exhibits missing ear) and the Lehner site (n = 13; one point exhibits distal impact fracture) in Arizona reflect minimal damage (Haury et al. 1953, 1959).
Ethnographic studies demonstrate that phytotoxins can greatly enhance the effectiveness of hunting weaponry including blowgun darts, arrows, and spears. Use of hunting poisons also reduces the risks associated with direct encounters with both terrestrial and marine mammals. And, they reduce pursuit costs once muscle relaxation, paralysis, and/or cardiopulmonary failure begins. Use of ethnopharmacology, concepts of pharmacokinetics and pharmacodynamics, and animal physiology involving body weight and metabolic rates provide a means for calculating lethal dosages for phytotoxins—specifically aconitine. One may, then, use these calculations of lethal dosages of poison to assess the feasibility of alternative hunting tactics by Early Paleoindians. The estimated dosages of aconitine required to kill a Columbian mammoth, woolly mammoth, and mastodon equal 149 mg, 88 mg, and 81 mg, respectively. Consequently, one-quarter teaspoon (3,000 mg) of aconitine could kill 20 Columbian mammoths, 34 woolly mammoths, or 37 mastodons. Parenthetically, Bisset (1976:94) states that the Ainu on Hokkaido would apply 600–700 mg of aconite poison to each arrowhead.
Ethnographic accounts conclude that hunting poisons do not adversely affect the consumption of meat and animal products. Marine mammals killed by circumpolar peoples, as well as bear dispatched by the Ainu with aconite, could be eaten without posing a threat to the consumers (Bisset 1976, 1979, 1981; Heizer 1938, 1943). In China, hunters used poisoned arrows and “the flesh around the wound was cut out” (Bisset 1979:355). The Semai Senoi, who used root bark from Strychnos species for dart poison, also cut away flesh around the dart wound before eating the meat (Bisset 1966:184). Lloyd (1897:405) states that African arrow poison extracted from the vines of Strophanthus hispidus posed no threat to those who consumed their prey. The Waata cut away the flesh that surrounded a poison arrow wound prior to consuming any of elephant flesh (Parker and Amin 1983:40). Given the small amount of poisons required to kill large mammals, the poison would have been greatly diluted once it entered the blood stream. For example, if a hunter managed to introduce 88 mg aconitine into the body of an adult male mammoth weighing 5,000 kg, the poison would then be diluted within body tissues as well as approximately 328 liters (86.65 gals) of blood.
Conclusion
At present, archaeologists have not observed any evidence for Early Paleoindian poison hunting. Yet archaeologists have not looked for such physical evidence until very recently. Similarly, archaeologists had not examined early Paleoindian projectile points for traces of protein-residue (blood) until relatively recently as well (Duke 2015; Loy and Dixon 1998; Seaman et al. 2008). Confirmation of the prehistoric use of hunting poisons may prove to be difficult but not impossible. Possible material correlates of early Paleoindian poison hunting might include:
The occurrence of chemical residue of aconitine (and/or other alkaloids) found on chipped stone weapon points as well as foreshafts (possibly the mammoth ivory rods associated with Clovis Period kill sites and caches). Weapon points can be kept relatively small when used in conjunction with hunting poisons. The chipped stone projectile points literally serve as the “leading edge” of the weapon and they provide an entry point for a long, thin foreshaft with a larger surface area coated with poison. Recent research by Valentina Borgia (Cambridge University) with Michelle Carlin (forensic chemist at Northumbria University) and Huw Barton (University of Leicester) suggest that chemical residues of hunting poisons may be identified on prehistoric hunting implements using liquid chromatography-mass spectrometry (LC-MS) (University of Cambridge 2015). At present, there is little available information regarding the length of time and physical conditions necessary for the preservation of aconitine and similar alkaloids
3
. State-of-the-art forensic methods used to detect such poisons can be expanded to include gas chromatography-mass spectrometry (GS-MS) and vibrational or Fourier-Transformed Infrared (FTIR) spectroscopy (see Baranska and Schulz 2009; Gajalakshmi et al. 2011; Sun et al. 2012; Yoshioka et al. 1996). Bradfield et al. (2015) caution that, “Working with unknown, decayed material from archaeological contexts where analysts have to rely on oxidative by-products, rather than the original compounds, further increases the difficulty in the accurate identification of substances.” Given such difficulties with biochemical markers, researchers have begun to rely upon DNA-based and molecular markers (see Srivastava et al. 2010). Srivastava et al. (2010:421) point out that analyses meant to identify biochemical markers are complicated by plant genetics and environmental factors. They continue, “This problem can be surmounted by using DNA-based markers and molecular markers…. DNA markers have now become a well-liked means for identification and authentication of medicinal plants….Also these DNA-based markers are less affected by age, physiological condition and other environmental factors” (Srivatsava et al. 2010:421). The presence of Aconitum pollen and starch particles on projectile points and foreshafts indicating the use of hunting poisons (University of Cambridge 2015). Pollen and starch granules representing the toxic alkaloids used for hunting poison may have been introduced into residential sites if the unprocessed plants (flowers and tubers) were initially transported to such locations. Pollen would be expected at locations where flowers were introduced and starch granules would more likely occur where Aconitum tubers were processed. It is important to point out that hunting poisons may not have been made in residential sites. Ethnographic accounts suggest that hunting poisons may, instead, have been produced at isolated logistical sites. Merker (1910) describes the use of hunting poisons on arrows and spears by the Wandor of East Africa, who used poison on their arrows and spears. He states, “To brew the poison two Wandor [unknown] obo always go a few thousand meters away from the camp into the bush” (Merker 1910:326). Similarly, with regard to the Strophanthus arrow poisons used by the Kabre in north Togo, Neuwinger (1996:609) states, “The preparation takes place in the bush.” And, among the Kamba of Kenya, the “medicine man” established their poison-making workshops along rivers some distance from settlements so that the “children should be kept away from the deadly poison [Acokanthera]” (Neuwinger 1996:66). Traces of pitch, resin, and/or carbon black used ethnographically to seal poisoned projectile points and foreshafts found on Early Paleoindian weapons. The Li-su of the Yunan and Sichuan provinces of China mixed aconite poison(s) with resin or vegetable gum before it was applied to arrow points and foreshafts (Bisset 1979:357). Ainu hunters on Hokkaido commonly made use of sealants such as pine resin and carbon black. Such treatments probably served to protect surfaces coated with lethal dosages of aconite-based hunting poison(s). Projectile points (chipped stone, bone, antler, or ivory) found in proximity to the gut and areas near the base of the ears of proboscideans at kill sites. As mentioned, ethnographic accounts of poison arrow hunting by the Liangulu (Waata) of southern Kenya suggest that these were preferred targets for historic elephant hunters using poison. Previously-discovered proboscidean kill sites should be carefully examined in order to identify possible projectile points of bone, antler, and ivory that may have been employed to deliver lethal or debilitating doses of hunting poisons.
For more than 175 years, naturalists, paleontologists, and archaeologists have assumed that prehistoric hunters killed mammoths, mastodons, gomphotheres, bison, and other prey using spears and darts that severely damaged critical organs and induced massive blood loss. Archaeologists, in particular, have relied upon the co-occurrence of carefully flaked projectile points and extinct animal remains to substantiate Paleoindian presence. Researchers have assumed that cooperative teams of hunters would have been required to mortally wound proboscideans and other megafauna during high risk, direct encounters. Yet, if hunting poisons were used during the LGIT, and possibly earlier during the Late Glacial Maximum in Asia, archaeologists may now be in a position to reassess hunting weaponry, hunting tactics and strategies, as well as previously unexplored aspects of Early Paleoindian knowledge related to ethnobotany and ethnopharmacology.
Footnotes
Acknowledgments
I would like to acknowledge the encouragement and support offered by Drs. Mary Ann Powell and Priscilla Grew. Comments by the late Norman C. Bisset as well as his extensive research on hunting poisons have served as a major source of inspiration. Also, I would like to thank Michael Jochim and Bradley T. Lepper for their earlier comments and encouragement. I am particularly grateful to Robert K. Hitchcock for directing me toward published and unpublished ethnographic literature—particularly with respect to the Waata (Liangulu) of Kenya. Finally, I would like to thank my wife Dr. Beth R. Ritter for her editorial advice. This paper is dedicated to Beth and my son Ian.
1
Total blood volume (Vb in liters) in relation to live body weight (M in kg) of mammals can be calculated using the equation, Vb = 65.6 M1.02 (Schmidt-Nielsen 1984). The blood volume, therefore, of a woolly mammoth weighing 5,000 kg equals 389 liters. Blood loss ranging between 30–33 percent can be life-threatening and blood loss more than 40–50 percent is fatal. Consequently, mortal wounds to a woolly mammoth would equal 156–194 liters (41–51 gal) of blood.
2
Wilmsen and Roberts (1984:171–172) describe an unfluted projectile point (distal fragment) that was found embedded within a cervical vertebra of a Bison antiquus at the Lindenmeier Site.
3
Aconitine is not water soluable. Toxicokinetic studies of human victims of aconitine poisoning indicate that it cannot be detected in human blood after 24 hrs (Fujita et al. 2007). The half-life of aconitine in the human body ranges between 5.8–15.4 h. After four half-lives, more than 94 percent of the alkaloid has been eliminated from the body (Yoshioka et al. 1996).