
Abstract
Homegardens are reservoirs of genetic resources. When native plant populations are compromised, traditional management of homegardens can be an effective means of conserving species and enhancing food security. Hylocereus undatus (dragonfruit) is a key species in homegardens but is rarely found in wild populations in the Yucatan. We analyzed management practices and genetic diversity of dragonfruit grown in homegardens in three climate regions (West, Central, and East) in the state of Yucatan, Mexico. Dragonfruit presence and reproductive phenology were documented via 2,660 surveys applied in seventeen municipalities. Eighty-nine semi-structured interviews were conducted to document dragonfruit management and use. Seventy-one specimens were collected among nine populations to characterize genetic diversity with DNA Inter Simple Sequence Repeats (ISSRs) markers. Dragonfruit was found in 12.4% of homegardens, propagated via cuttings (98%), received minimal agricultural management, and flowered and fruited in different seasons among regions. Its fruits were consumed fresh or as a beverage and different parts of the plants were used to treat dysentery, kidney stones, hair loss, and gastritis. Genetic diversity among regions was moderate (PLP [polymorphic loci] = 25.1 to 53.3%; He [expected heterozygosity] = 0.075 to 0.138), but the polymorphism information content (PIC) values were at the most 20% lower than those estimated from an ex situ germplasm collection of Hylocereus spp. A Bayesian assignment analysis identified two genetic lineages (K = 2): one in the West and East regions and another in the West and Central regions. Genetic differentiation Jaccard indexes among accessions from all homegardens were similar to those found among wild accessions from Mexico. We conclude, therefore, that homegardens in Yucatan are important in conserving the regional biocultural heritage of dragonfruit.
Keywords
Introduction
Traditional agroecosystems function as genetic resource reservoirs when wild populations of native species have declined due to deforestation (Bhagwat et al. 2008; Sharma and Vetaas 2015). Conservation of native species may therefore fall to agroforestry systems, which are largely managed by Indigenous groups (Boege 2008; Casas et al. 2016; Sharma and Vetaas 2015; Vera V et al. 2017). The Maya homegarden is an example of an agroforestry system in which plant and animal production has not only provided subsistence to rural Indigenous populations for centuries but has also helped conserve native species (Colunga-García Marín and Zizumbo-Villarreal 2004; De Clerck and Negreros-Castillo 2000; Jimenez-Osornio et al. 2005; Mariaca-Méndez et al. 2010; Rico-Gray et al. 1991).
The Yucatan Peninsula is a priority region for genetic resource conservation because the millennia-old management systems of the Yucatec Maya, including homegardens, have resulted in enhanced agrodiversity (Boege 2008). However, as a result of deforestation of dry tropical forests, epiphytes are threatened in the Yucatan Peninsula, especially in the Yucatan state (Ellis and Porter-Bolland 2008; Olmsted and Juarez 1996; Porter-Bolland et al. 2015). Growing epiphytes in Yucatec Maya homegardens could, thus, play a central role in epiphyte conservation in the region.
Dragonfruit (Hylocereus undatus), known as pitahaya in Spanish, is an epiphytic cactus grown for centuries for its edible fruit and traditional medicinal properties (Bravo-Hollis and Sánchez-Mejorada 1978; Colunga-García Marín and Zizumbo-Villarreal 2004). Native to Mesoamerica (Bravo-Hollis and Sánchez-Mejorada 1978; García-Rubio et al. 2015), it has the potential to be highly productive in tropical and subtropical climates (Montesinos-Cruz et al. 2015). It appeals to international markets (Shelef et al. 2016) and its phytochemical traits make it apt for use in the food and pharmaceutical industries (Jeronimo et al. 2017; Mohamed et al. 2018). Several studies of dragonfruit have characterized its fruit and stem morphological variations and examined the effect of agricultural practices on fruit production in commercial plantations throughout Mexico (reviewed by Ortiz-Hernández et al. 2012). However, information about the management of dragonfruit in the Yucatan Peninsula remains limited (Andrade et al. 2006; Herrera-Krings 2016; Jhibran et al. 2003; Manzanero-Acevedo et al. 2014; Yam Noh 2002), even though this region has the largest commercial production of dragonfruit in Mexico (Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación 2017).
Dragonfruit grown in homegardens plays an important role in Yucatec Mayan food security and health, more broadly. In 2008, for instance, dragonfruit production in homegardens was approximately three tons/ha on 50 hectares (Becerril et al. 2009). Dragonfruit fruit, roots, and flowers are used as medicines for their hypoglycemic, diuretic, cardiovascular tonic, antiseptic, and wound healing properties (Ankli et al. 1999; Argueta-Villamar et al. 1994; Mendieta and Del Amo 1984). Hylocereus undatus has a high nutritional value and relatively low amounts of soluble sugars. Its low sugar content is especially important because of the prevalence of child and adult obesity and diabetes in the Yucatan state associated with the consumption of high-caloric drinks and processed foods (Barquera et al. 2013; Hall et al. 2017; Loria et al. 2018).
This study documents several aspects of the ethnoecology of dragonfruit in Maya homegardens as a way to promote its conservation. Specifically, we document the prevalence, management, and use of dragonfruit in three climatic regions in the state of Yucatan to assess if the species is underutilized (Padulosi and Hoeschle-Zeledon 2004). We compare these results to the reproductive phenology of dragonfruit in commercial plantations to find if there is more diversity in the Maya homegardens than in commercial operations. We also analyze the genetic variability of H. undatus in homegardens and compare it to that of existing germplasm collections of the species to evaluate the role of homegardens in dragonfruit conservation. The results of this study demonstrate the cultural importance of H. undatus and the role of homegardens as biocultural reservoirs for this species.
Materials and Methods
Hylocereus undatus (Cactaceae), known to the Maya as chakam, sakam, or chak wob (Mariaca-Méndez et al. 2010), is an epiphyte with long, thin triangular stems. Its large, white flowers bloom at night, generally from May to October. The fruit is normally round with a bright red to pink peel and scale-like protrusions; fruit is produced from June to November (Castillo-Martínez et al. 2005).
The species is cultivated in homegardens in the states of Campeche, Quintana Roo, and Yucatan (Flores-Guido 2012). It grows wild in high and low dry tropical forests and evergreen tropical forests, and occasionally grows on xerophilic scrub (Cálix de Dios 2004; García-Rubio et al. 2015; Ortiz-Hernández et al. 2012). Hylocereus undatus produces fruit both by cross-pollination and self-pollination (Valiente-Banuet et al. 2007), although self-incompatible accessions have been reported in Quintana Roo (Castillo-Martínez et al. 2005). Fruit production is greater when produced by cross-pollination than with self-pollination (Nerd et al. 1999; Valiente-Banuet et al. 2007; Weiss et al. 1994). Because the flowers are chiropterophilous, their most common visitors are bats (Leptonycteris curasoae and Choeronycteris Mexicana) and sphingid moths (Sphinx ligustri) (Nerd and Mizrahi 1997; Valiente-Banuet et al. 2007; Weiss et al. 1994). Indeed, the species' main pollinator in Yucatan is reported to be Sphinx ligustri (Herrera-Krings 2016). Several local populations of H. undatus may be extinct in the state of Yucatan, which has been deeply deforested (Ellis and Porter-Bolland 2008; Olmsted and Juarez 1996).
Study Site
The state of Yucatan is located in the Yucatan Peninsula biogeographic province (Espinosa-Organista et al. 2008). Its elevation gradient ranges from 0 to 400 meters above sea level (masl). According to the Köppen classification modified for Mexico (García 1973), a humid tropical climate (three climate variants of type A) predominates, although an area of dry climate (two climate variants of type B) is present along the northwest coast (Figure 1). Vegetation cover today is mostly for agriculture and livestock and includes some remnants of the original dry tropical forest, semi-evergreen tropical forest, and evergreen tropical forest (Ellis and Porter-Bolland 2008; Porter-Bolland et al. 2015).
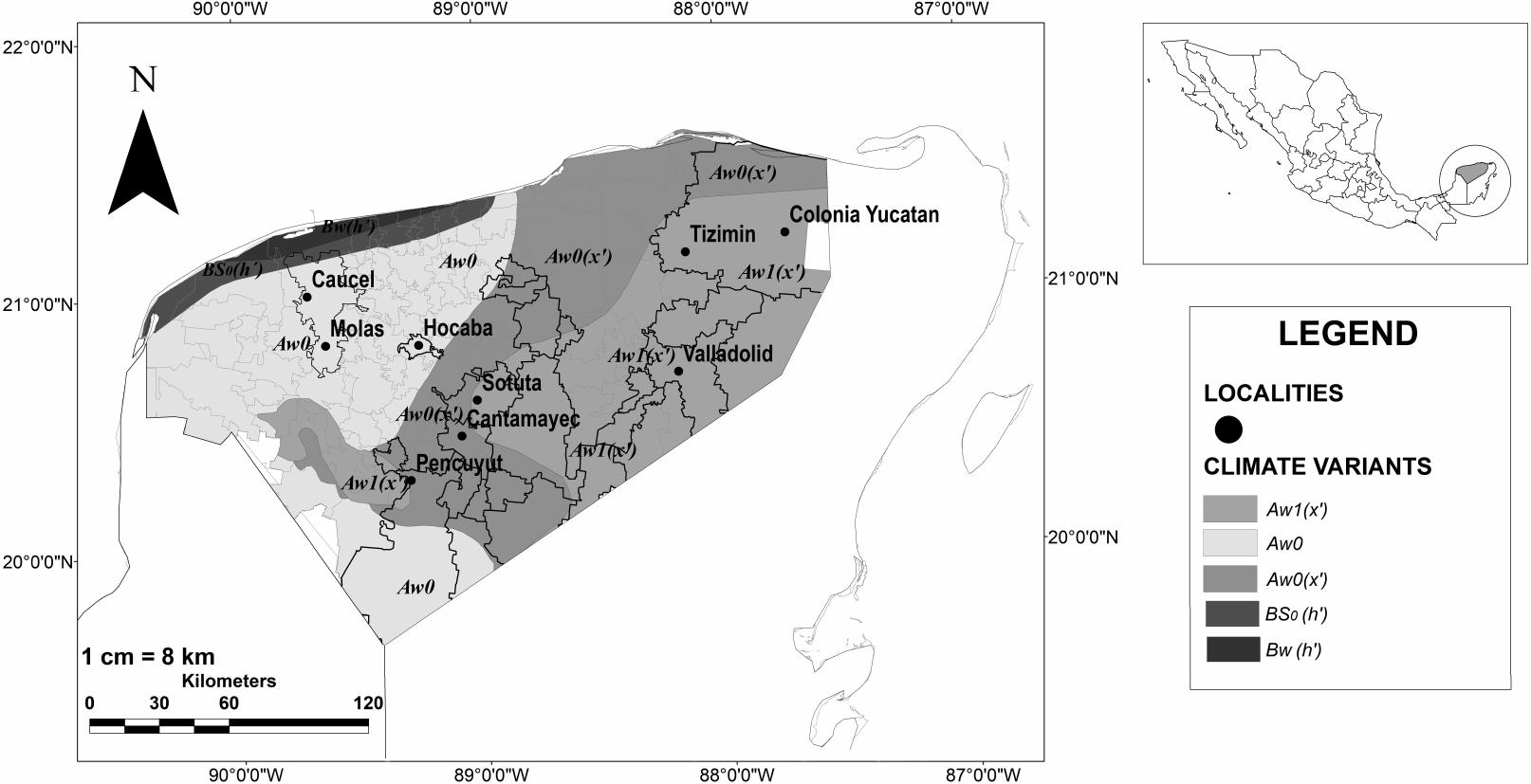
Study area: Yucatan state in Mexico (inset map), municipalities of Yucatan state where surveys were done (outlined with solid lines), and communities in the state of Yucatan where stems of Hylocereus undatus were collected for molecular analyses (black dots) by climate variant regions West Aw0, Central Aw0(x′), and East Aw1(x′).
All families in the rural communities of the region have homegardens that are structured similarly to the native forest and contain many of the same species (Colunga-García Marín and Zizumbo-Villarreal 2004; De Clerck and Negreros-Castillo 2000). At least 387 species of plants are present in the Maya homegardens, representing one-sixth of the native flora reported for the Yucatan Peninsula (Herrera 1992). The 22 core species (mainly trees) present in the homegardens all have at least one use (Jimenez-Osornio et al. 2005). Homegardens are richly complex agroecosystems in which cultural, social, ecological, agronomic, and economic factors converge to maintain a large diversity of animal and plant species that meet the food, health, and recreation necessities of the family (Jimenez-Osornio et al. 2005). Maya homegardens, thus, represent a dynamic system in which adaptation processes of native and introduced plant and animal species are continually taking place; they are sites where agricultural diversity has been and continues to be maintained (De Clerck and Negreros-Castillo 2000; Jimenez-Osornio et al. 2005).
Region Classification
For analytic purposes, data were grouped into three regions (West, Central, and East) based on climate variants within the type A climate (Figure 1). Climatic variant distribution was based on meta-data provided by the biodiversity page of the Comisión Nacional de Biodiversidad (CONABIO) (CONAIBO and Gárcia 1998). The West and Central regions contain two tropical climate variants that are the driest of the subhumid variants (P/T ≥ 43.2mm/°C): subhumid with summer rains and subhumid with summer rains and a 10.2% winter rain proportion (García 1973; Orellana et al. 2010; Figure 1). The East region is a variant of tropical subhumid with summer rains, intermediate between the subhumid variants (P/T < 43.2 mm/°C and ≤ 55.3 mm/°C) with a 10.2% winter rain proportion (García 1973; Orellana et al. 2010; Figure 1). The differences between these climate variants suggest a rainfall gradient that increases from west to east.
H. undatus Prevalence and Management in Family Homegardens
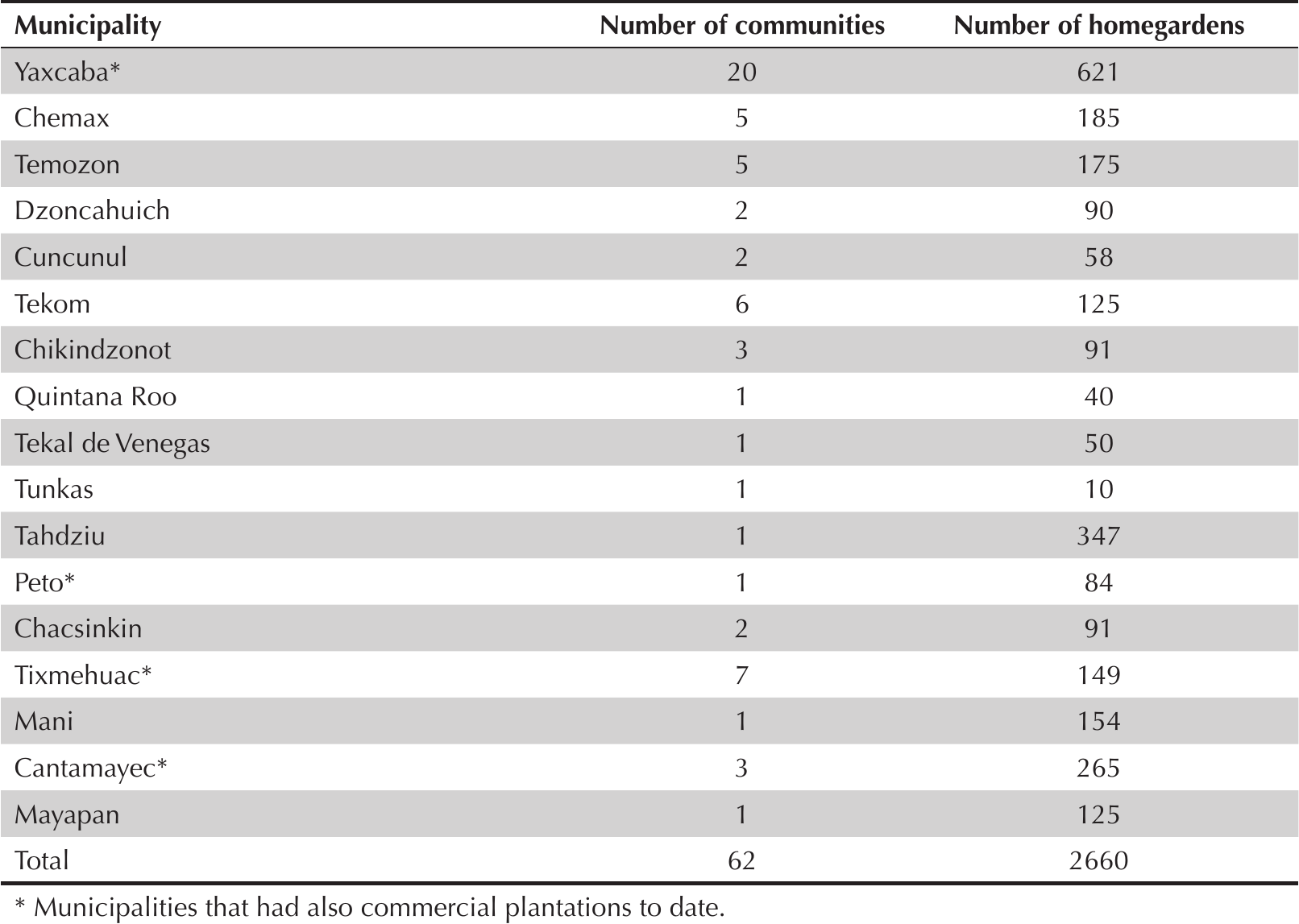
To characterize the prevalence and management of dragonfruit, field work was conducted in two stages. During the first stage, data on presence/absence, origin, and flowering and fruit production seasons of H. undatus in homegardens were collected by including four questions on H. undatus management on a survey given to 2,660 owners of homegardens that were participating in the Secretaría de Desarrollo Social (SEDESOL) “Programa Producción Social Familiar de Traspatio” (Gobierno del Estado de Yucatán 2016) in 17 municipalities in the state of Yucatan (Table 1; Figure 1). The survey questions were in Spanish, but a Yucatec Maya translator assisted in asking the questions when necessary.
Number of communities and homegardens surveyed in the municipalities of the state of Yucatan.
The four questions [and possible responses] were: 1) Do you have dragonfruit in your homegarden? [Yes/No]; 2) Where did you get the plant? [It was a gift/ I bought it/It was already there/Other]; 3) What season does it flower? [Dry (March to June)/Rainy (summer July to October)/ Northwinds (autumn and winter rains November to February)]; and 4) What season does it produce fruit? [Dry/Rainy/ Northwinds]. Results were analyzed to evaluate whether there were regional variations in the frequencies of: 1) the presence of dragonfruit cultivation in homegardens; 2) plant origin; 3) flowering season; and 4) fruit production season. Significant χ2 results were processed with a residuals analysis to identify which frequencies were above or below expected values (Haber-man 1973).
Based on the relative abundance of cultivated H. undatus in homegardens, six communities across the three regions were chosen for the second stage of field work: West (Caucel, Hocabá, and Molas), Central (Cantamayec and Sotuta), and East (Yaxcabá). In 2015, a total of 89 home gardeners were interviewed using semi-structured interview questions. Of these, 60 had H. undatus in their homegardens. In addition, we interviewed 29 people who were referred to us by the first group as people having experience in the management of H. undatus. The interview guide was aimed at obtaining descriptions of the management and use of dragonfruit (see Supplement 1). Because the number of communities differed between regions, as did the number of interviews held per community, the results were pooled for an overall analysis for the entire state.
To identify if dragonfruit varieties in homegardens may have had an extended reproductive phenology, data on the frequency of flower and fruit production in the Yucatan state were compared to those reported for commercial/industrial plantations of H. undatus in other Mexican states (Castillo-Martínez and Ortiz-Hernandez 1994; Martínez-Ruiz et al. 2017; Osuna-Enciso et al. 2016). The results of the management practices interviews were compared to those obtained from a similar survey of Campeche homegardens (Manzanero-Acevedo et al. 2014).
Genetic Variability
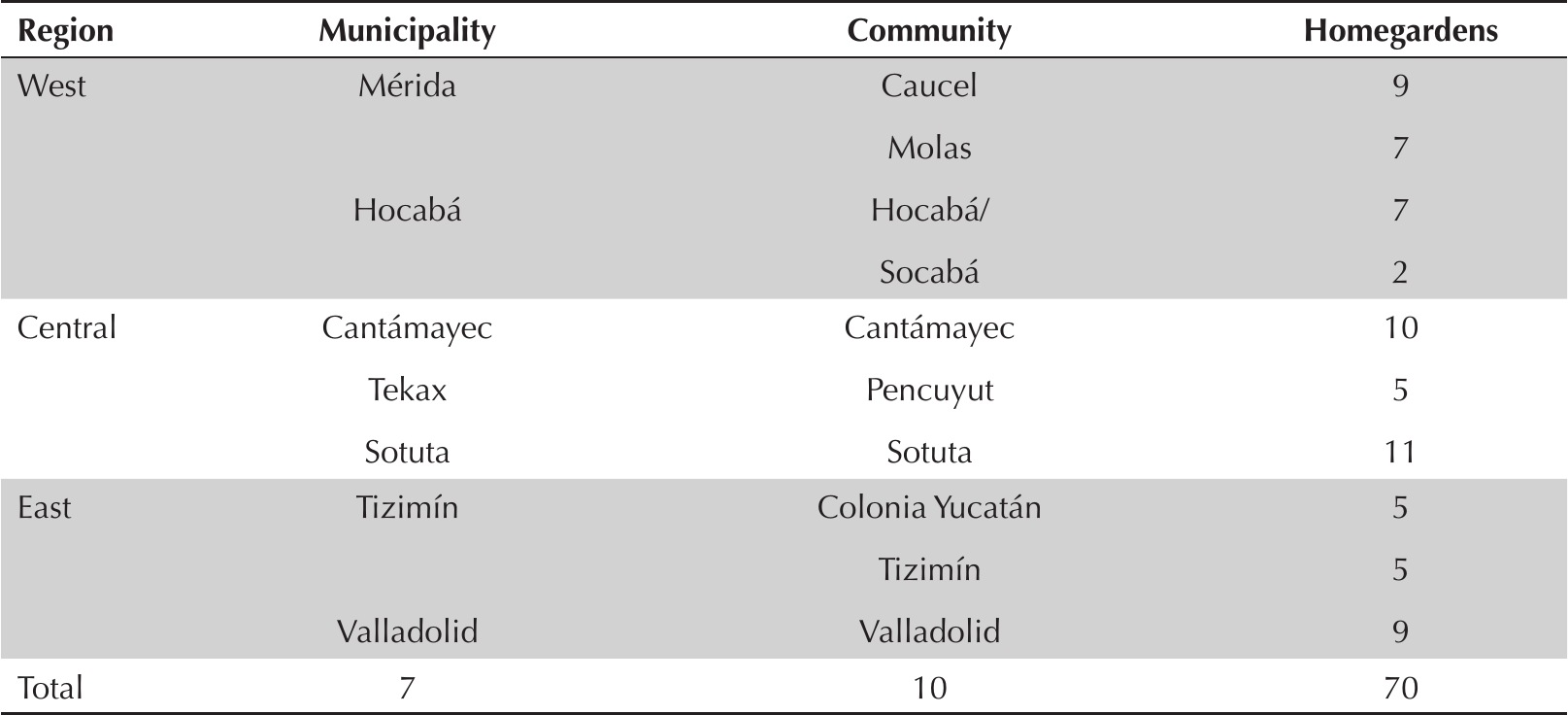
Genetic variability was characterized for three communities in each region: West (Caucel, Hocabá, Molas), Central (Cantamayec, Pencuyut, Sotuta), and East (Colonia Yucatán, Tizimín, Valladolid). The nine communities yielded a total of 70 homegardens where H. undatus was cultivated (Table 2). When permitted by owners, 5 cm-long cuttings of H. undatus were collected. The distal portion of each cutting was dissected in 1 cm-thick discs; these were cut into small cubes (0.25 cm3) to be stored at -20 °C until DNA extraction.
Number of homegardens where stems were collected for molecular analysis in different communities of the municipalities of the state of Yucatan.
DNA Purification and Amplification
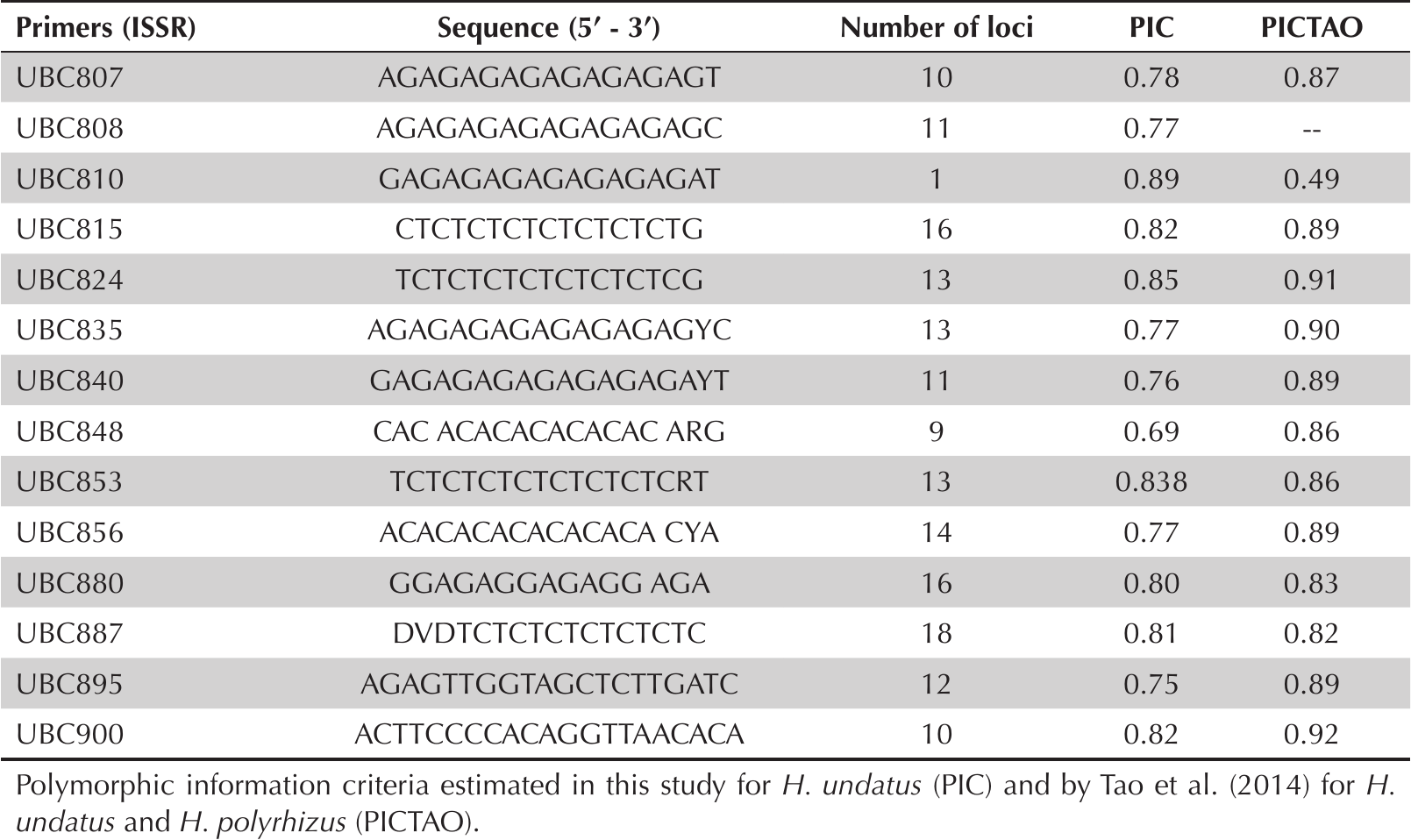
DNA was purified following the CTAB extraction method as modified for Cactaceae (Falcón and Valera 2007). Following the protocols described in Supplement 2, genomic DNA was amplified for previously reported DNA Inter Simple Sequence Repeats (ISSRs) (Tao et al. 2014; Table 3).
Sequence of thirteen ISSRs primers from the set 9 designed by the University of British Columbia (UBC) for molecular marker amplification in Hylocereus and Polymorphic information criteria (PIC).
Statistical Analysis
Each identified band was considered a locus. A total of 167 bands were amplified for the fifteen primers and presence/ absence data recorded for each locus (Table 3). Statistics obtained for populations and regions included: percentage of polymorphic loci (PLP), expected heterozygosity (He) (Vekemans 2002), and qII statistic (Holsinger et al. 2002). Additionally, polymorphic information criteria (PIC) was estimated for loci as described by Tao et al. (2014). Genetic relationships were inferred using a Bayesian grouping approach (Pritchard et al. 2000). Consensus dendrograms were obtained for Nei's genetic distances among populations and regions (Nei 1978) as described by Tel-Zur et al. (2004) and for Jaccard coefficient among individuals as described by Legaria-Solano et al. (2005). Detailed information of estimations for statistics is included in Supplement 2.
The PIC values were compared to those from a germplasm collection of 51 cultivated lines and wild accessions of H. undatus and H. polyrhizus (Tao et al. 2014). The ranges of genetic distances were compared to those obtained for germplasm collections of: i) 230 cultivated lines and wild accessions of eight species and hybrids from them (Pagliaccia et al. 2015); and ii) wild populations of H. undatus throughout its distribution in Mexico (Legaria-Solano et al. 2005).
Results
H. undatus Prevalence and Management in Family Homegardens
From the initial survey of owners of homegardens (N = 2660), H. undatus was grown by 330 of them; of that 12.40 %, 1.65% were found in the West, 7.66% in the Central, and 3.08% in the East region. This interregional variation (χ2 = 10.44; p = 0.0054) in dragonfruit cultivation from the initial survey (N = 2660) can be explained by lower than expected frequency of dragonfruit cultivation in the East region (9.62%), with higher values in the West (13.01%) and Central (14.01%) regions. The actual distribution of H. undatus in the Yucatan state suggests that the most adequate climate variant is the driest of the subhumid variants (P/T = > 43.2mm/°C), and the East region is the least suitable because it has the wettest climate variant. However, the proportion of origins of the H. undatus plants in the homegardens was constant across the three regions (χ2 = 8.38; P = 0.2116): 36.23% as a gift; 10.80% purchased; 42.86% present previously, and 10% other.
Interregional variations were observed for flowering season (χ2 = 29.20; p < 0.0001) and fruit production season (χ2 = 89.32; p < 0.0001). The differences in reported flowering season were due to differences between expected and observed frequencies. For example, there was an excess of reports of flowering during the northwinds season in the Central region and during the rainy season in the East region, but a deficiency of reports of flowering during the northwinds season in the East region (Figure 2a). Other discrepancies included an excess of reports of fruit production during the rainy season in the Central and East regions and a deficiency of reports of flowering during the northwinds season in all three regions (Figure 2b).
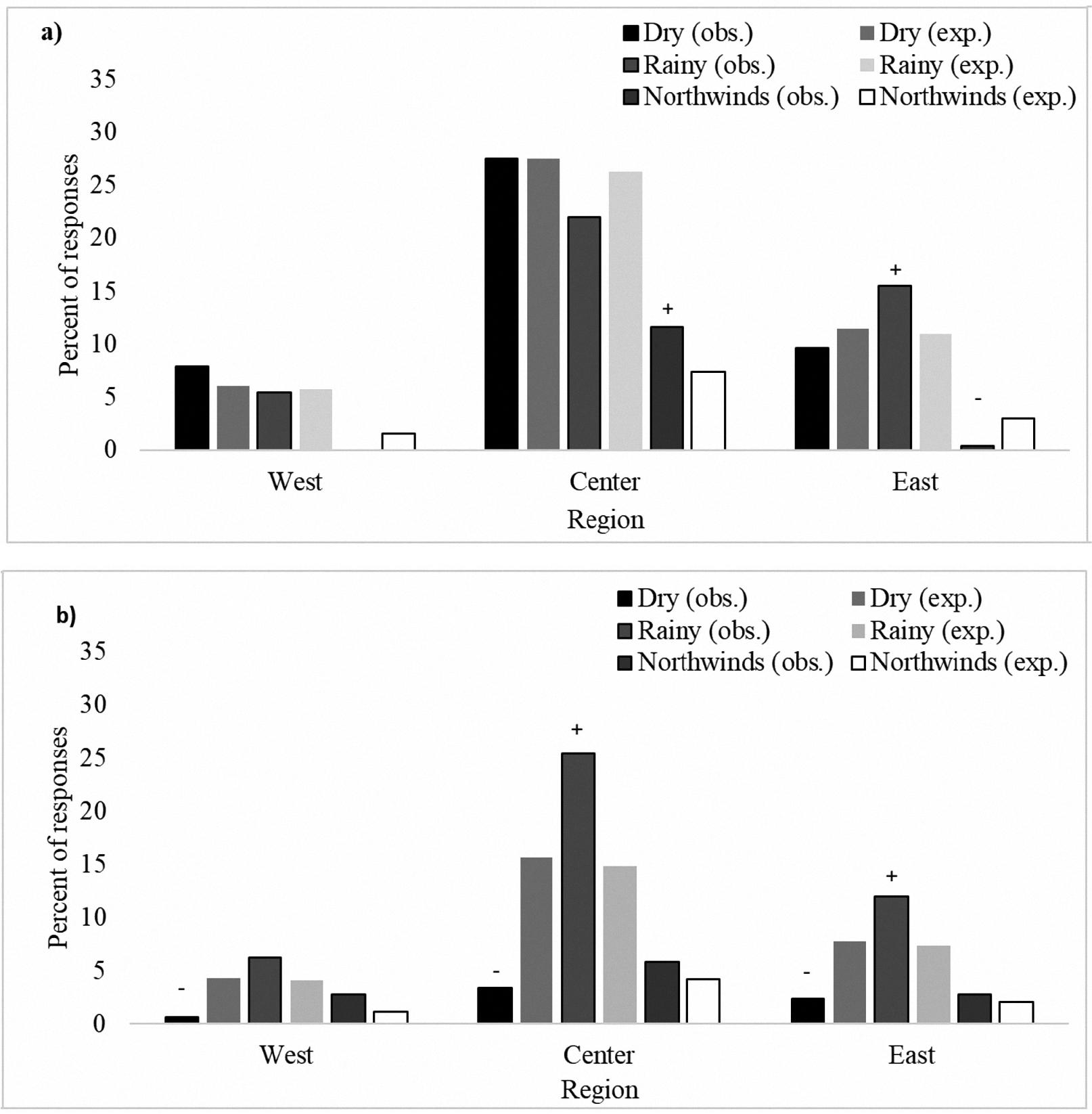
Observed (obs.) and expected (exp.) reported frequencies per season for a) flowering and b) fruit production (N = 2660) in West, Central, and East regions of the state of Yucatan.
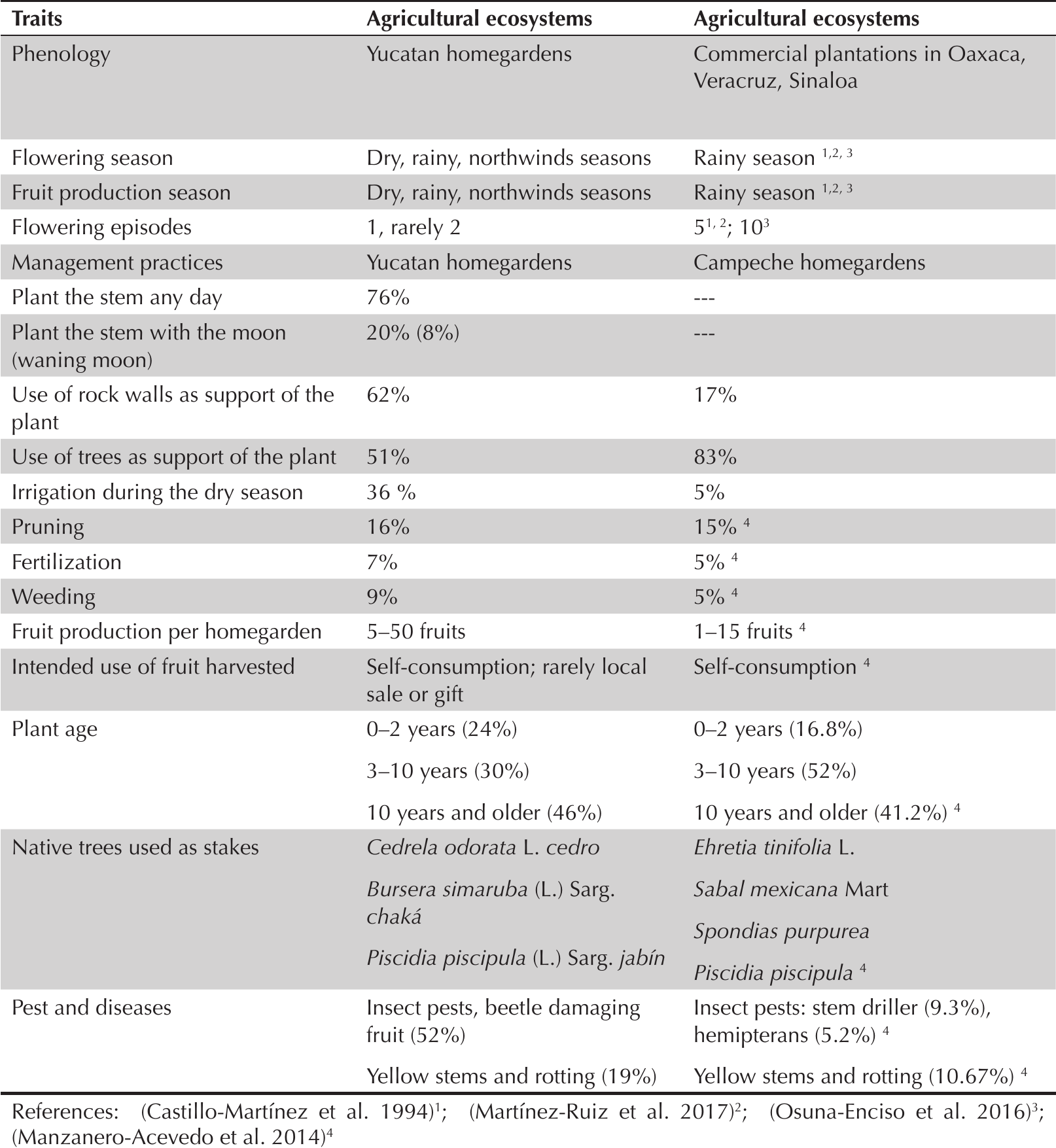
Our results demonstrate a predominance of flowering and fruit production in the rainy season, which largely coincides with the flowering and fruit phenology of dragonfruit growing in commercial plantations of Oaxaca, Veracruz, and Sinaloa (Castillo-Martínez and Ortiz-Hernández 1994; Martínez-Ruiz et al. 2017; Osuna-Enciso et al. 2016). However, while flowering episodes are more frequent in the commercial plantations, they are restricted to the rainy season. This may be compared to Yucatan homegardens, where owners report flower and fruit production also occurring during dry and northwind seasons (Table 4). H. undatus plants that bloom and produce fruits out of the flowering peak provide food security to homegarden owners, as the fruits are still available to them when commercial production is low and market prices high in Yucatan.
Comparisons of the phenology variation in flowering and fruit production seasons; flowering season and management practices of cultivation of Hylocereus undatus in Yucatan homegardens and other agricultural systems in Mexican states.
Of the 89 interviewees, 89% spoke Yucatec Maya and identified themselves as Mayan (see demographic questions in Supplement 1). This indicates that homegardens including dragonfruit are mainly managed by this ethnic group.
Only one dragonfruit plant was cultivated in most of the Yucatan homegardens, probably due to its extended epiphyte growth habit. However, some interviewees said they had more than thirty plants. Of the interviewed home gardeners, most (55%) had acquired cuttings by asking for them, 20% from a neighbor, and 2% from another community. These results bring out the importance of the traditional exchange of plants for the homegarden owners. Also worth mentioning is that dragonfruit propagation in the homegardens of Yucatan is not exclusively via cuttings, since 38% of owners reported plants in their garden that had not been planted or were preexisting. Therefore, Mayan homegardens may maintain plants that were originally propagated by natural vectors or had been cultivated by older generations and left in the homegardens.
Dragonfruit management practices in Yucatan homegardens varied widely, though differed only slightly from the management practices reported by Manzanero-Acevedo et al. (2014) for 384 homegardens in the state of Campeche (Table 4). Most Yucatan producers seem aware that dragonfruit requires only minimal management when grown in homegardens.
All those interviewed said they consumed the fruit fresh or as a beverage. A small number (2%) consumed it in January (northwinds season) and April (dry season); the rest did so during the rainy summer season (June to October). This seems to be due to the variation in reproductive phenology and because the plants mostly had only one flowering episode (Table 4). A popular fruit widely used in fruit drinks, 13% of the interviewees described dragonfruit as “healthy” and containing “vitamins” and “nutrients.” Due to its wide acceptance, dragonfruit beverages constitute a healthier alternative to commercial sodas for Yucatec Mayan families. While 83% of the interviewees recognized the medicinal properties of different parts of the dragonfruit plant, only 19% noted that it is used for curing a particular disease. Seven out of the 89 recognize it can be used to cure kidney stones, four out the 89 to cure dysentery, five out of the 89 to cure and prevent hair loss, and three out the 89 to cure gastritis.
Genetic Variability
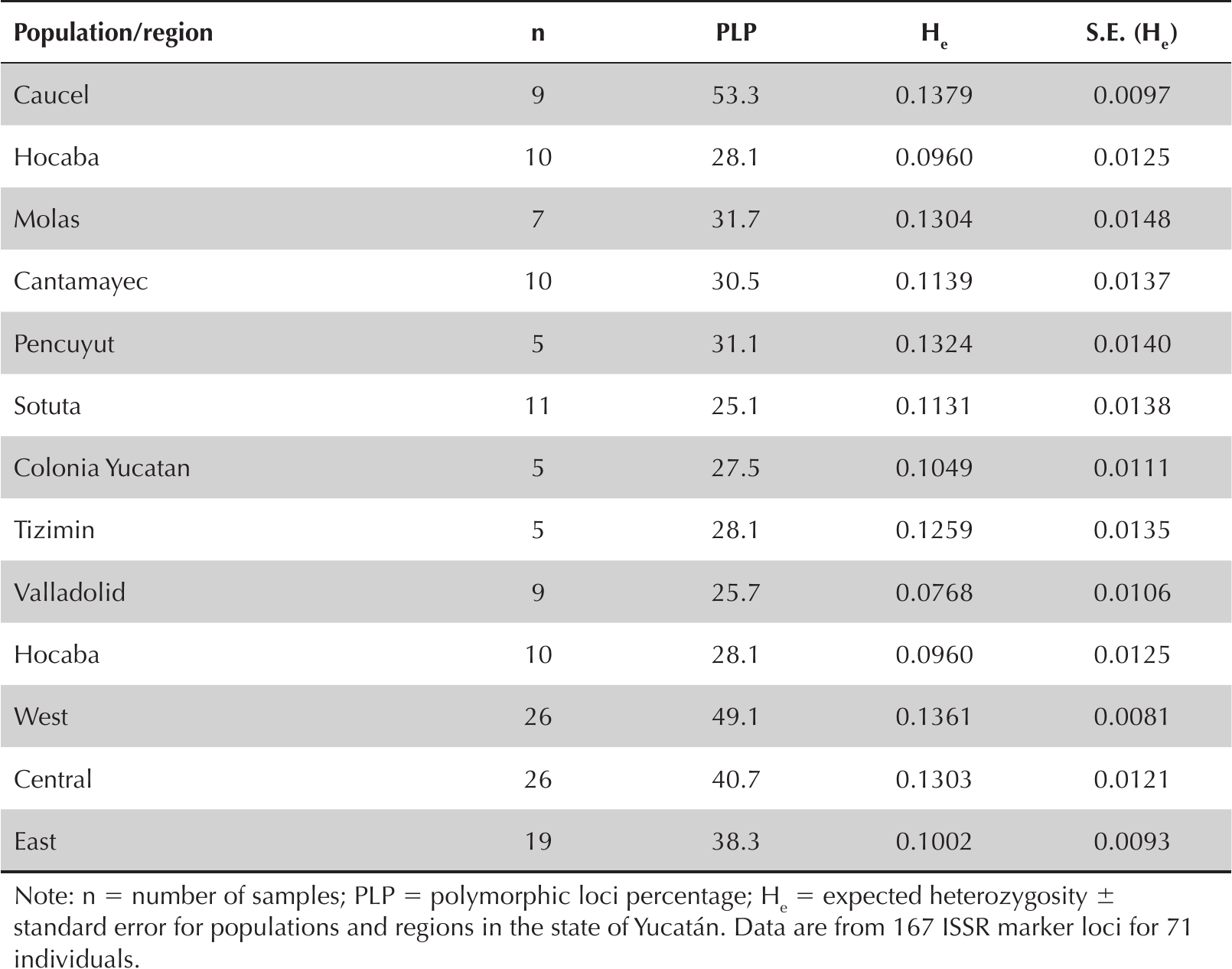
The genetic diversity for H. undatus under cultivation was moderate, with an allelic diversity (PLP) ranging from 25% to 32%. However, at Caucel, it was almost double (53%), indicating that double forms of alleles can be found in that community. Between regions, PLP ranged from 38.3% in the East to 49.1% in the West, which suggest that the West region contains slightly more diverse allele forms than the East region. Among the populations, He was highest at Caucel, Molas, and Pencuyut, intermediate at Cantamayec, Sotuta, and Tizimín, and lowest at Colonia Yucatán, Hocabá, and Valladolid, suggesting that more homozygous genotypes are found in the latter communities (Table 5). This parameter was higher in the East than in the West and Central regions, suggesting that West and Central regions tend to have more homozygous genotypes of the species (Table 5).
Hylocereus undatus genetic diversity statistics per population and region.
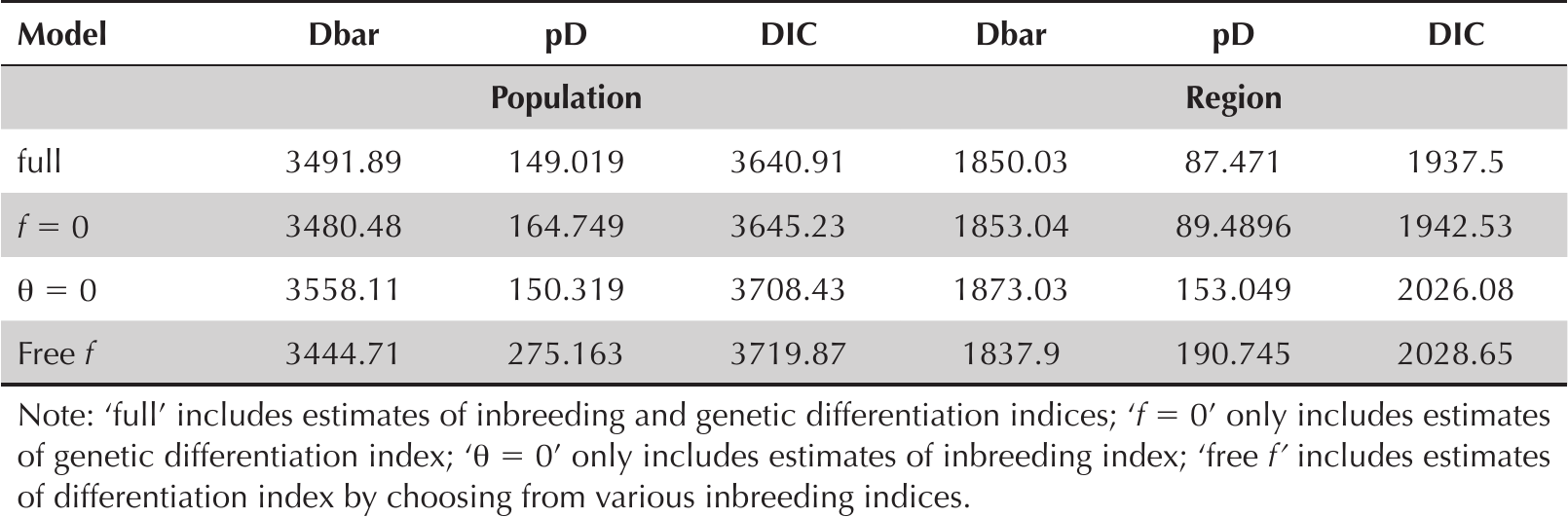
On the other hand, the genetic variability displayed by the individual plants in the homegardens is comparable and sometimes higher than for those maintained in ex situ germplasm collections. The PIC values estimated in this study were at most 20% lower than those recorded from the Chinese germplasm collection for the wild and cultivated accessions from H. undatus and H. polyrhizus (Table 3). Genetic differentiation was slightly lower between regions (qII = 0.0034 ± 0.0025) than between populations (qII = 0.0049 ± 0.0031). The f = 0 model provided the best fit to the data grouped by population, suggesting that inbreeding within populations is avoided, while the full model had the best fit for those grouped by region, suggesting that there may be characteristic inbred lines in each region (Table 6). The Bayesian assignment analysis identified two different genetic groups (K = 2). Most of the collected plants were assigned to the first group with a probability greater than 0.9 (Colonia Yucatán, Hocabá, Tizimín, Valladolid); the remainder were assigned to the second group (Cantamayec, Molas, Sotuta), which largely coincides with the dendrograms obtained for genetic distances among individuals, populations, and regions (Figure 3). At least two genetic variants, coinciding with the two documented genetic groups, were present in the homegardens of Yucatan; those in the Central and West regions are of special interest. Among the individuals of these two groups, genetic differentiation ranges from 0.002–0.8465 (Figure 3c), values that are comparable to those estimated in the analysis of genetic differentiation of Mexican wild accessions from the entire distribution of H. undatus in the country (Legaria-Solano et al. 2005). Genetic differentiation values in this study were higher than those recorded from a reference germplasm collection of the genus maintained at the University of California, in which the distances from a group of genetically improved variants and a group of wild accessions of dragonfruit were estimated in 0.1 changes using AFLP molecular markers (Pagliaccia et al. 2015). These comparisons highlight the importance of homegardens as in circa situm reservoirs of genetic diversity, which is larger than that maintained in ex situ germplasm collections.
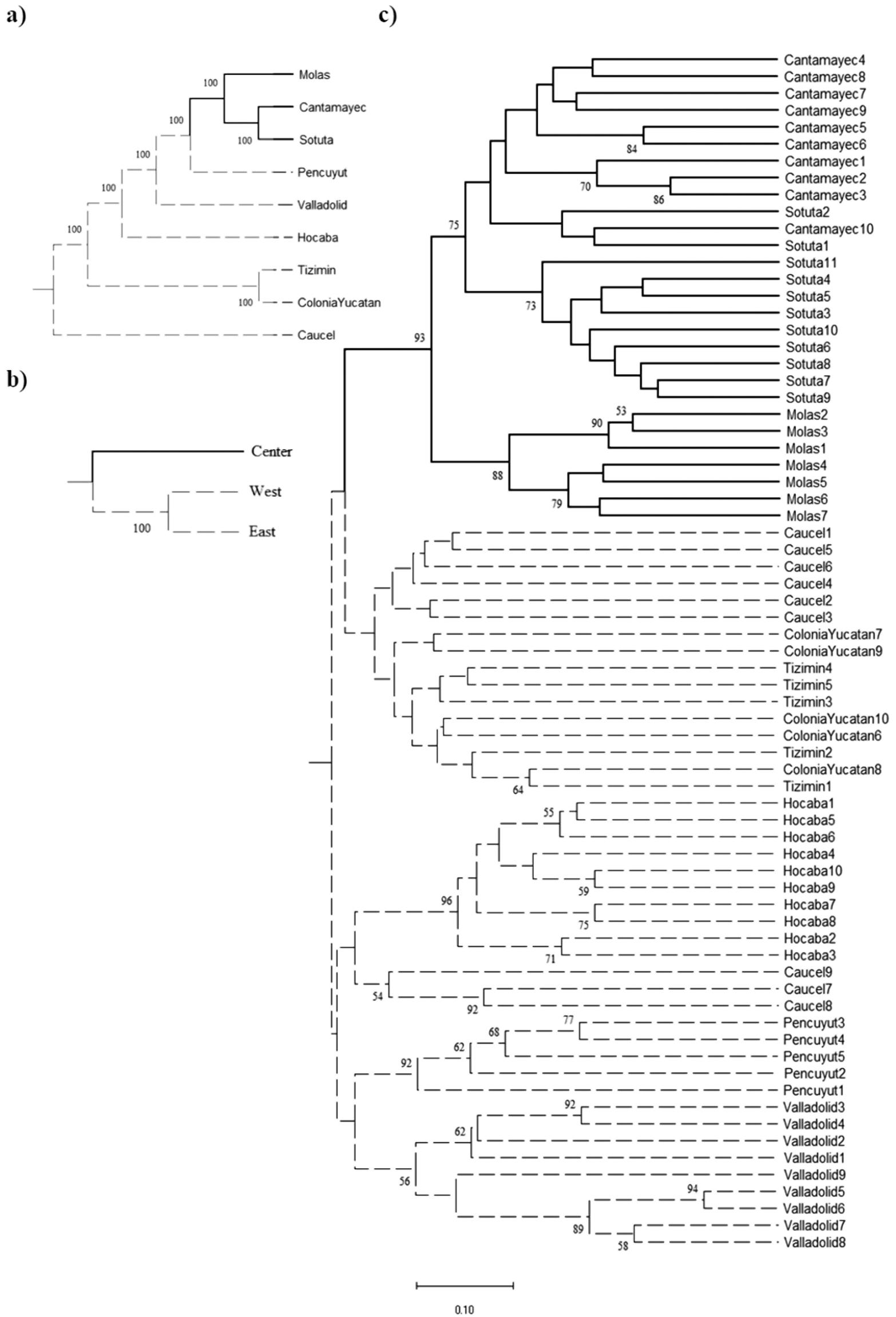
Nearest neighbor dendrogram for Nei genetic distances between a) populations and b) regions; c) UPGMA dendrogram for Jaccard coefficients among individuals of Hylocereus undatus from the state of Yucatan. Branch shading corresponds to Bayesian assignment to Group 1 (dashed line) or Group 2 (solid line).
Data fit values for Hylocereus undatus allelic frequencies (Dbar), parameters (pD), and deviance information criterion (DIC) for selection among four models.
Discussion
The results indicate that H. undatus is cultivated in Yucatan homegardens, although agricultural management is low. The plants are mainly used for alimentary and medicinal purposes. The variation in flowering and fruit production seasons between climate regions, as reported by interviewees, suggests that some homegarden varietals bloom and produce fruit earlier and later than those from commercial plantations. Also, the species varies in prevalence of genetic group, resulting in genetic structuring between the populations and climate regions where collections were made. This means homegardens maintain equal or larger genetic variability than ex situ germplasm collections. Taken together, these results indicate that homegardens are reservoirs of H. undatus diversity and that cultivation under traditional management is maintained because of the extensive use of dragonfruit as food and medicine.
When native species are not only cultivated less frequently but also consumed less often, the loss of the species within Mayan homegardens and traditional knowledge about their management may be hastened (Montañez-Escalante et al. 2015; Moo-Aldana et al. 2017; Rivas 2003). A relatively low frequency cultivation of H. undatus in seventeen municipalities of Yucatan (12.4%), and particularly in the East region of the state, contrast with the 26% frequency reported in the neighboring state of Quintana Roo (De Clerck and Negreros-Castillo 2000). This fact suggests that this species may be underutilized in the Yucatan state. However, its frequency in homegardens is higher than that of other species that were once as common as dragonfruit, including Pouteria campechiana (known as canister), Diospyros digyna (known as tauch), Parmentiera aculeate (known as pepino kat), and a traditional variety of Carica papaya (known as papaya mamey) (Montañez-Escalante et al. 2015). While many underutilized species are excluded from the agendas of policy makers (Padulosi and Hoeschle-Zeledon 2004), the government began providing incentives to introduce dragonfruit to commercial plantations in Yucatan in 1996 (Meráz Alvarado et al. 2003; Rodríguez-Canto 2000). The Fondo Nacional de Apoyos para las Empresas en Solidaridad (FONAES) program supported dragonfruit cultivation in ten Yucatan municipalities: four in the West region (Dzidzantún, Halachó, Kopomá, Mérida), three in the Central region (Buctzotz, Dzilam González, Tahdziu), and three in the East (Temozón, Quintana Roo, Espita) (Meráz Alvarado et al. 2003; Rodríguez-Canto 2000). To date, dragonfruit is grown commercially in eleven municipalities in the West (Cansahcab, Dzidzantún, Halachó, Hunucmá, Kinchil, Samahil, Seyé, Suma, Temax, Tetiz, Yobain), nine in the Central region (Akil, Cantamayec, Dzilam González, Muna, Otzkutzcab, Peto, Ticul, Tixmehuac, Tzucacab), and one in the East (Yaxcabá) (Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación 2017).
We documented dragonfruit under cultivation in homegardens in four of these municipalities (Table 1). Cultivation in homegardens may be displaced when more productive commercial varieties are adopted over traditional ones (Rivas 2003; Whitney et al. 2017), after which traditional knowledge of their use becomes compromised (Montañez-Escalante et al. 2015; Moo-Aldana et al. 2017). The potential for loss of cultivated dragonfruit from homegardens is of concern because the diverse management practices in Mayan homegardens contribute to the diversity of cultivated dragonfruit varietals and, thus, increase its availability throughout the year. Early- and late-blooming and fruit-bearing varieties of dragonfruit could be selected in homegardens to further enhance variations in reproductive phenology and improve overall fruit production and availability for consumption, contributing to food security. Whether the presence of commercial plantations compromises the existence of traditional cultivation in homegardens deserves future research.
Implementation of programs supporting management of native fruit crops in homegardens provides another opportunity for the conservation of species, such as H. undatus, that are widely cultivated and consumed by the Yucatec Mayans. This is significant considering the documented reduction of homegarden size (Jiménez-Osornio et al. 2018) and even their abandonment due to biological and social factors, such as emigration of rural populations in Yucatan and the availability of fruits from commercial plots (Montañez-Escalante et al. 2014). Such phenomena represent potential hazards for the cultivation of H. undatus and fruit trees in general (Montañez-Escalante et al. 2014).
Conservation of dragonfruit in Mayan homegardens fosters diversity of the agroecosystem and maintainence of desirable phenotypes for genetic improvement of the species. Due to the epiphytic nature of dragonfruit, homegarden cultivation preserves the biotic interactions with the tree species that are used to stalk the plant and enhances the overall diversity of the agroecosystem. Therefore, conservation of epiphytes may increase the value of homegardens as conservation sites in general, as has been recorded for another epiphyte Laelia anceps in the Montaña of the Guerrero region (Herrera-Cabrera et al. 2018). Furthermore, conserving and managing dragonfruit native varieties may also be important, as they represent the genetic resources for breeding programs that look to extending the seasonal availability of fruit production under low-input agronomic requirements and preserving the genetic variability of the species.
Traditional management of H. undatus maintains comparable and even higher levels of genetic variability than that of ex situ germplasm collection. Genetic bottlenecks are common in cultivated plants, particularly in those propagated clonally, which result in reduced genetic diversity (Miller and Gross 2011). However, genetic variability is promoted when genetic material is exchanged among homegarden owners from different communities or recruited by sexual reproduction from preferentially cross-pollinated species (Hamrick et al. 1992). Dragonfruit populations exhibited moderate genetic diversity (ca. 50% lower) compared to that estimated for other cacti such as Stenocereus thurberi and Rhipsalis dissimilis (Bustamante et al. 2016; Giovanoni 2014), suggesting that the species might be going through a genetic bottleneck. Nevertheless, the exchanges of cuttings (ca. 50% of cuttings were obtained as gifts or bought) and the cultivation of plants originating via outcrossing in homegardens contribute to the genetic variability found in H. undatus from Maya homegardens. Therefore, Maya homegardens should be valued as reservoirs of dragonfruit genetic resources in circa situm compared to ex situ germplasm collections, as has been documented for the preservation of both native and exotic crops around the world (Galluzzi et al. 2010; Heraty and Ellstrand 2016) and for Annona squamosa in Mayan homegardens (Salazar et al. 2010).
The importance of H. undatus for food security, combined with its variability in traditional use and management, reproductive phenology, and genetics, suggests that Yucatan Mayan homegardens should be considered sites of conservation in circa situm. This native species may be locally extinct in the wild because the state of Yucatan has been deeply deforested (Ellis and Porter-Bolland 2008). Therefore, due to its importance in the traditional food and health system of the Yucatec Mayans, its conservation in Mayan homegardens should be encouraged.
Footnotes
Acknowledgments
The research reported here was financed by a grant to MMF from the Consejo Nacional de Ciencia y Tecnología (Ciencia Básica 169336-B), to JJO from Secretaría de Desarrollo Social Yucatán (Propuesta para la instalación y operación de las Agencias de Desarrollo Humano Local, como estrategia para erradicar la pobreza en Yucatán), and by an undergraduate scholarship awarded to AH (578607) from the Consejo Nacional de Ciencia y Tecnología. The authors thank the owners of the homegardens for their support of this research and Dana Lepofsky, Alex McAlvay, Jaida Samudra, and one anonymous reviewer for their valuable comments on the original manuscript and the editorial team of Journal of Ethnobiology.