
Abstract
Herbaceous wetland plant resources have been widely cultivated and utilized by Indigenous peoples throughout North America since at least the early Holocene. Archaeologists and ethnographers, along with traditional knowledge holders, have documented and reconstructed deep histories of interaction between human communities and coastal plants that provide dietary carbohydrates, medicinal compounds, and craft-fiber. On the Florida peninsula, as elsewhere, paleoethnobotanical researchers face challenging preservation conditions and, despite the ubiquity and vastness of coastal wetlands, the resident flora are conspicuously underrepresented in the archaeological record. In this study, we work toward the recognition of wetland plant use on the Gulf Coast of Florida by integrating analyses of archaeo-molluscan, microfaunal, and palynological assemblages from stratified shell-midden deposits at a village and civic-ceremonial center occupied across the first millennium AD. We identify four particular herbaceous wetland plants as likely subsistence, medicinal, and technological resources. In a brief discussion, we propose that coastal wetland flora likely played key roles within late-Holocene maritime resource intensification, civic-ceremonial aggregation, and village-coalescence.
Introduction
Herbaceous coastal wetland plants have been utilized by Native American peoples as valuable subsistence and technological resources since at least the early Holocene (Hoffman et al. 2016; Nicholas 1998; Turner 2014) and appear commonly in ethnohistoric records from Florida (Baber 1942; Worth 2014), the greater southeastern U.S. (Harvard 1895; Whitford 1941), and farther afield (see Deur and Turner 2005). Paradoxically, coastal wetland plants are woefully underrepresented in the archaeological record and macroscopic remnants are preserved only under the most favorable conditions, such as within wet-sites, sealed hearths, rock shelters, and caves (Erlandson et al. 2019; Lyons et al. 2018; Tuross et al. 1994; Vellanoweth et al. 2003). In the frequently inundated, well-drained, quartz-sand sediments of Florida's coastal strand, macroscopic archaeobotanical remains of starchy corms, rhizomes, and stem-fibers are very rarelypreserved.Thus, whileancientcoastal peoples in Florida inhabited ecosystems dominated by marshes and swamps dense with edible and technologically useful flora, there is practically no archaeological evidence for the harvesting, processing, or consumption of these plants; they “remain invisible in the archaeological record to date” (Hutchinson et al. 2016:66).
In this study, we propose that environmental archaeologists may circumvent unsuitable conditions for macrobotanical preservation at coastal village sites and recognize past traditions of plant use by analyzing classes of small durable bycatch that accompanied plant resources from harvesting loci to storage and processing areas, where they accumulated within cultural soils. To support this proposition, we synthesize zooarchaeological and archaeobotanical data from excavation units and core samples at the Crystal River site (8CI1)—a village and civic-ceremonial center on the north peninsular Gulf Coast of Florida that was occupied across the first millennium AD. Within Crystal River's stratified midden record, we identify: 1) conspicuous concentrations of wetland-associated silicate sponge spicules that indicate the transport of wetland vegetation and/or associated soils from local marshes and swamps to village activity areas; 2) a characteristic assemblage of small, marsh-dwelling gastropods collected incidentally as bycatch during plant-harvesting; and 3) fossil pollens from a set of wetland forbs that enable the identification of likely subsistence, technological, and medicinal resources.
Extant Archaeobotanical and Bioarcheological Evidence
Archaeological research on Florida's Gulf Coast first gained broad anthropological attention at the close of the nineteenth century, following F. H. Cushing's ([1897] 2000) excavation of painted wooden masks, canoe paddles, fishing nets, and other botanical artifacts from water-logged deposits at Key Marco. Paradoxically, the following half-century of archaeological work in the region would foreground ceramic and lithic technologies and largely neglected the roles of plant resources in ancient Indigenous lifeways. Working with immense volumes of shell and bone at shell-terraformed village sites, twentieth century archaeologists cast ancient Gulf Coast societies as sedentary fisherfolk or affluent foragers (sensu Moseley 1975; Sahlins 1968) that depended fundamentally upon the natural productivity of the region's fisheries (Goggin and Sturtevant 1964; Marquardt 1992; Widmer 1988). The persistence of this perspective has led to a constructive regional emphasis on past sea-level dynamics and the capture of estuarine fauna, but has also perpetuated the faulty assumption that Precolumbian Gulf Coast peoples were not cultivators and deemphasized the importance of ancient plant use traditions.
Countering these trends, Newsom and Scarry's (2013) comprehensive work with macrobotanical remains at South Florida sites has identified a suite of floral resources that were widely used along Florida's coastal strand (Supplementary Table 1). In southwest Florida, these resources included “quasi-domesticate” varieties of papaya and squash/gourd, suggesting that late-Precolumbian peoples of the region “were cultivators who purposefully maintained distinct cultivars and other valuable plant resources” (Newsom and Scarry 2013:298–299). Macrobotanical evidence for the cultural importance of wetland forbs within ancient Gulf Coast lifeways is considerably more limited (Newsom and Scarry 2013:288–289); however, the use of marsh and swamp-understory plants is suggested by stable isotope analyses of human remains from Precolumbian Gulf Coast mortuary mounds (Hutchinson et al. 2016).
Previous research with starch grain and phytolith residues has illuminated the use of archaeologically elusive floral resources such as Zamia sp., Manihot sp., and Maranta sp. by Precolumbian peoples in the Bahamas (Berman and Pearsall 2008), the larger Caribbean (Pagan-Jimenez 2011), and throughout the neo-tropics (Dickau et al. 2007). No successful phytolith or starch grain work from coastal Precolumbian contexts in Florida has been published and preservation issues abound in this region due to sandy, well-drained soils, the general fragility of starch granules, and the dissolution of phytoliths within frequently inundated alkaline sediments. Attempts by specialists, and by the authors, to isolate and analyze starch grains and phytoliths from cultural soils at the Crystal River site encountered serious preservation issues consistent with mastication by sand grains and dissolution (Cummings and Varney 2013, 2015).
Archaeology of the Crystal River Site (8CI1)
The Crystal River site is situated on the north peninsular Gulf Coast of Florida—a region known as Florida's Big Bend—that extends northward 300 km from the central Gulf Coast barrier island chain to Apalachee Bay on the panhandle coast (Tanner 1960). In this region, siliciclastic-sediment starved streams and an exceptionally low topographic gradient have produced vast expanses of low-energy coastal marshes that open to the Gulf of Mexico (Hine et al. 1988; Hutton 1986). These coastal marshes are organized along topographic and salinity gradients. Freshwater marsh and swamp understory taxa proliferate near spring heads and frequently include cattails (Typha sp.), freshwater sedges (e.g., Rhynchospora sp.), emergent aquatic herbs (e.g., Saururus cernuus, Pontederia cordata, Sagittaria sp.), and ferns (e.g., Acrostichum sp.). Brackish zones support dense expanses of sawgrass (Cladium mariscus) and other generalist sedges (Eleocharis sp., Cyperus sp.). Brackish zones that experience greater marine influence are dominated by broad swaths of black needle rush (Juncus romerianus) with fringes of smooth cordgrass (Spartina alternaflora) along the channels of tidal creeks. These coastal marshes are dotted with wooded islands and peninsulas that denote topographic highs of the underlying limestone substrate, and—almost without exception—contain Precolumbian archaeological deposits (Sassaman et al. 2017; Williams et al. 2007).
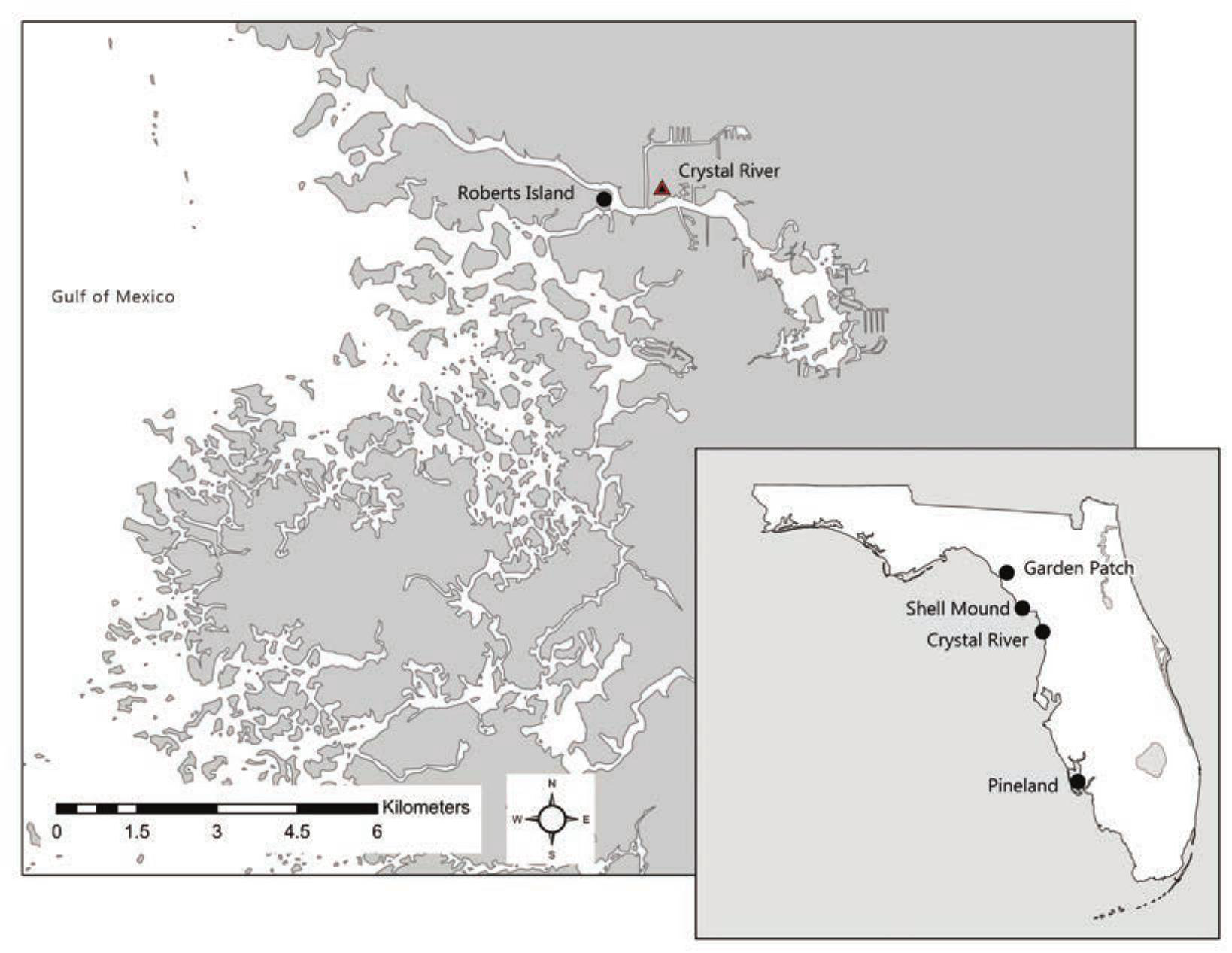
Locations of Crystal River (8CI1) and other sites mentioned in text.
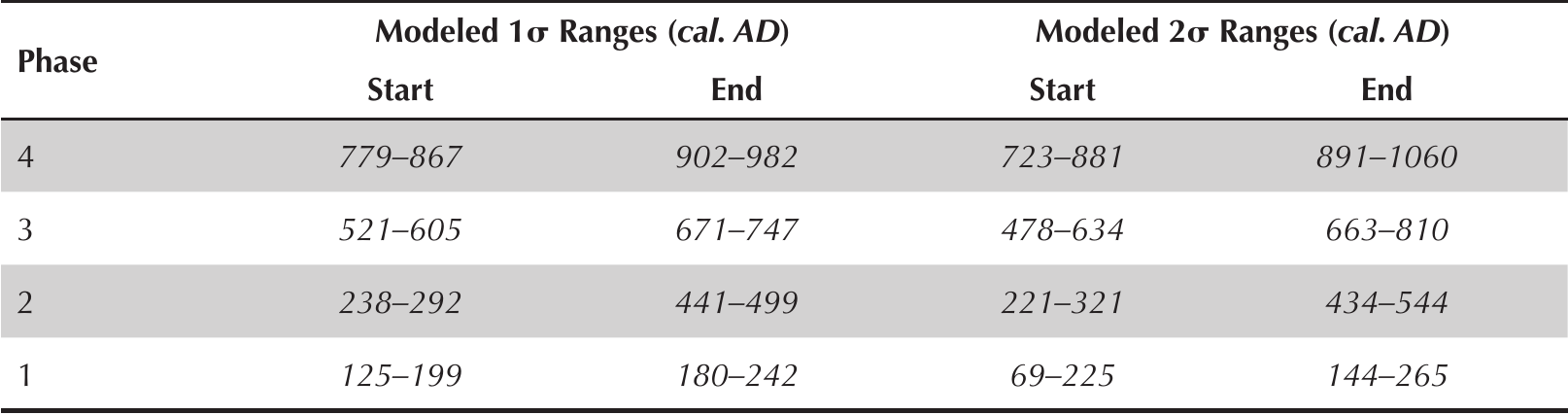
Ancient communities of the Big Bend Coast constructed the Crystal River site on the northern bank of the Crystal River about half-way along its nine-kilometer run (Figure 1). More than 30 radiocarbon dates obtained during recent coring and excavation at Crystal River enabled Pluckhahn and colleagues (2015) to develop a Bayesian statistical model for the chronology of site activity that we adopt here (Table 1). Earthwork construction began by at least 200 BC with the formation of two sand and shell mortuaries (Mound G and portions of the Main Burial Complex) (Pluckhahn and Thompson 2018). Seasonal or episodic residential occupation began during the first century AD and, after ca. AD 220, the Crystal River site supported a considerable number of permanent or semi-permanent villagers, with local population swelling during seasonal, annual, or multi-annual social aggregation events. At the final phase of construction, ca. AD 800, the mound complex featured two mortuary mounds, three platform mounds, a well-defined plaza, and a deeply stratified shell midden ridge (Figure 2).
Bayesian phase date ranges for occupation at Crystal River (8CI1), adapted from Pluckhahn et al. (2015).
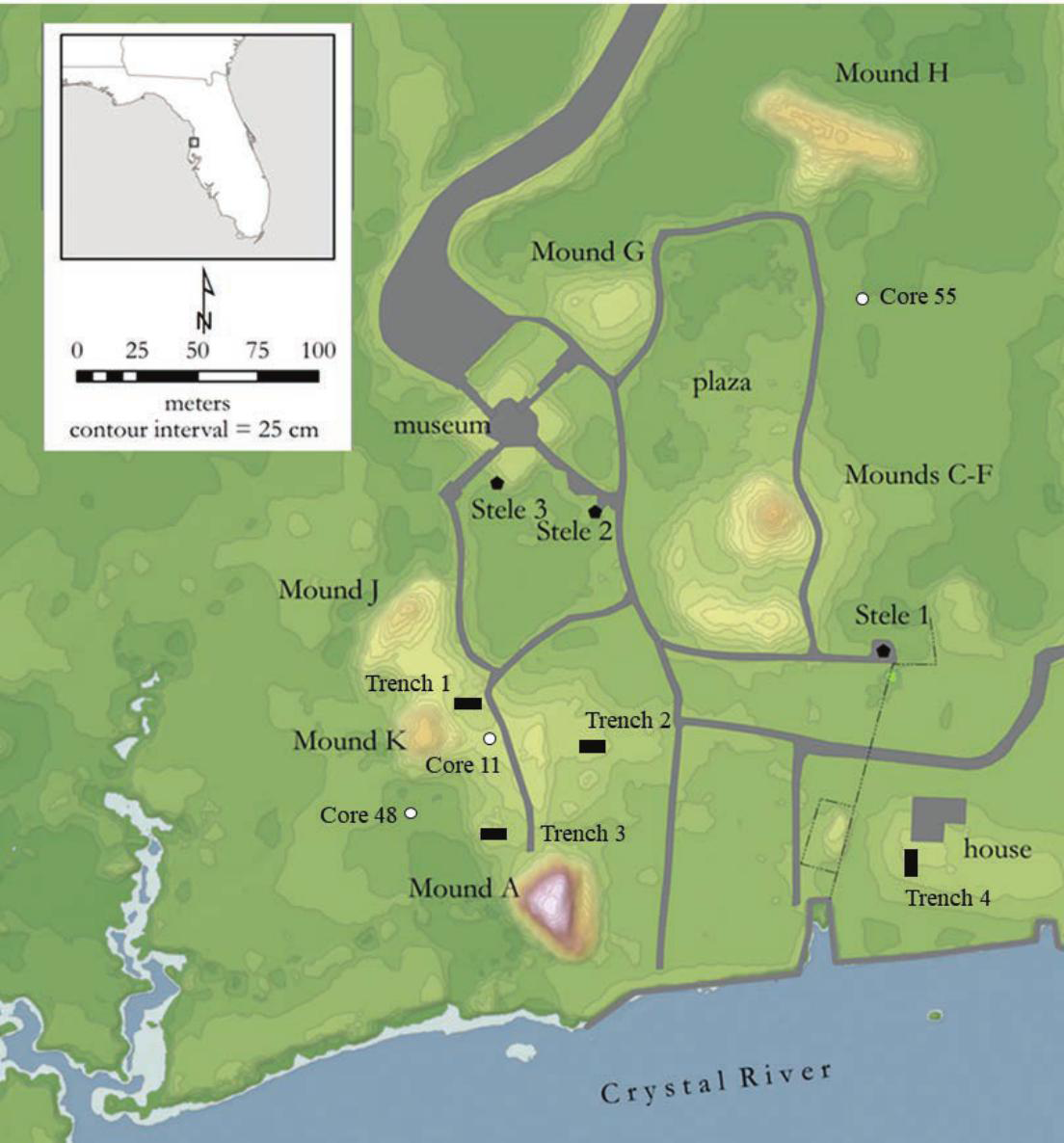
Site layout of Crystal River (8CI1) with locations of core samples and excavation trenches.
In recent zooarchaeological work, Duke and colleagues (2020) analyzed invertebrate and vertebrate assemblages from well-dated contexts within a 1 × 2 m test unit excavated into a central portion of the midden ridge at the Crystal River site. Biodiversity and equitability estimates calculated from these data were compared with like estimates from zooarchaeological assemblages collected from two 50 × 50 cm shovel tests at the Robert's Island mound complex (8CI41)—an island-hammock site roughly one kilometer downstream that was established as occupational and civic-ceremonial use of the Crystal River site waned in the seventh and eighth centuries AD. Duke and colleagues (2020) find that assemblages at Robert's Island exhibit considerably less biodiversity and equitability estimates than those from Crystal River—indicating a trend of decreasing diet breadth that they attribute to the development of corporate resource intensification strategies designed to mitigate environmental perturbation and negotiate new socio-political dynamics. The intensified harvesting of oyster (Crassostrea virginica) and marsh clam (Polymesoda caroliniana) at Robert's Island appears to drive this biodiversity trend and may also signal increased utilization of oligohaline marsh habitats (see Heard 1982:21).
Palynological analyses at Crystal River were conducted on core samples collected from village features and adjacent wetlands in order to track changes in vegetation communities across the late-Holocene (Jackson and Pluckhahn 2020). Variation in pollen spectra, ca. 2500 BC to AD 1050, evidence the local manifestation of regional climatic trends, as well as meso-scale patterns in marine transgression. Further, the relative prevalence of early-successional, disturbance-marker taxa (Solanum sp., Brassicaceae, Commelina sp., Salix caroliniana) within “black-earth” midden strata deposited during phase 2 (cal. AD 221–544) corroborate sedimentological and geochemical evidence for year-round site use during this interval (Jackson and Pluckhahn 2020:71–73).
Research Design and Methods
Many of Florida's most useful coastal wetland forbs do not produce durable seeds or woody tissues and are unlikely to be represented in macrobotanical samples. These taxa do, however, flower and produce pollen, which the plants release to the air column (anemophily), to insects (entomophily), and/or to vertebrate animals (zoophily). Inferring plant use from palynological assemblages requires close consideration of pollination strategies. Anemophilous plants release their pollen on the wind to join the mostly arboreal pollen rain, which may be transported considerable distances (hundreds of kilometers) before settling out of the air column. In contrast, entomophilous and zoophilous taxa have restricted ranges of pollen dispersal generally limited to areas directly below plant stands (see Hicks and Hyvärinen 1986:225; Hyde and Williams 1945; Potter 1967), but include areas where plant material or underlying soils have been transported, stored, processed, or discarded. Thus, while pollen from anemophilous taxa may be expected in some abundance across wide areas, pollen from entomophilous and zoophilous wetland taxa may serve as a marker for the localized presence of plants and/or associated soils.
If ancient coastal dwellers were harvesting and processing flora from local marshes and swamps, then we should also expect evidence of bycatch in the form of small fauna that are closely associated with marsh and swamp understory plants (e.g., Ainis et al. 2014). In our research area, these fauna include freshwater/brackish sponges (Family Spongillidae) that proliferate alongside emergent aquatic vegetation within shallow wetlands and leave behind diagnostic silicate skeletal spicules (Eshleman 1950; Schwandes and Collins 1994). Additionally, a variety of small (< 2 cm) gastropods occur in close association with freshwater and brackish wetland flora, which they depend on for anchoring substrate, forage, and/or structure for escaping aquatic predators (Heard 1982). Small gastropods have elsewhere been intentionally harvested for subsistence and medicinal uses in both coastal (Giovas et al. 2013) and inland freshwater (Klippel and Morey 1986) environments; however, despite relatively frequent occurrence within Florida Gulf Coast midden deposits, there is no published evidence in the region for targeted exploitation of small snails.
Field Methods
In Summer 2011, sediment cores were collected from village features and adjacent wetland areas at the Crystal River site as part of a site-wide geoarchaeological survey (Pluckhahn et al. 2015). A Geoprobe hydraulic coring system was utilized to recover 4.5 cm diameter cores from site features, and wetland cores (also measuring 4.5 cm in diameter) were recovered using a small, hand-operated vibracore system. Three cores were selected and sub-sampled for stratigraphic palynological analysis, including: one core from an intact section of the site's midden ridge (Core 11), one core from a stand of marsh southwest of the village midden (Core 48), and one core from the hydric hammock east of the site's plaza (Core 55) (Figure 2). Four excavation trenches (measuring between 1 × 2 and 1 × 4 m) were placed across the extent of the midden ridge at the Crystal River site in order to define the composition, stratigraphy, and chronology of village occupation. Units were excavated in 10 cm levels within observed strata, and sediments were processed through 3.2 mm (.125 in) screen in order to facilitate the recovery of fish bone, small mollusk remains, and small artifacts. Figure 3 displays the southern stratigraphic profile from Trench 1—which approximates the sedimentary character and pattern of midden deposits encountered in all four trenches, as well as in Core 11.

South profile of Trench 1, showing black-earth deposits (Str. Va, Str. IV) between layers of oyster midden (Str. II, Str. III, Str. V). Chronological associations: Strata I and II: unassigned; Stratum III: phase 3; Stratum IV: phase 2; Stratum V: phase 2; Stratum Va: phase 1.
Laboratory Methods
Sediment cores were described and sub-sampled stratigraphically for geoarchaeological and palynological analyses. Individual sub-samples were processed to simultaneously yield pollen and silicate microfossil assemblages. Laboratory procedures followed principles in Faegri and Iversen (1989), more detailed protocol by Bryant (2014), and were modified as needed to account for abundant carbonates, charcoal, and fine sand. To facilitate simultaneous isolation of fossil pollen and silicate microfossils (i.e., sponge spicules and phytoliths), samples were processed by heavy liquid separation in saturated zinc chloride solution.
Taxonomic identifications of pollens, diagnostic phytoliths, and sponge spicules were made via light microscopy between 400 and 1000x magnification. Pollen identifications were made to the lowest possible taxonomic specification and cross-checked using modern comparative collection specimens and regional identification keys (e.g., Willard et al. 2004). All intact sponge spicules and spicule fragments measuring greater than 50 microns in length were counted during pollen analysis to estimate relative spicule frequencies. Preserved diagnostic phytoliths were limited to those of the palm Family (Arecaceae) but were counted to estimate their relative frequencies. All pollens, identified phytoliths, and sponge spicules were summed to yield total microfossil values, from which relative percent of total microfossil values were derived.
Molluscan remains recovered from 3.2 mm screening were processed following protocols established by Reitz and Wing (2008) and identified to the lowest taxonomic specification possible (genus or species). Intact gastropods were counted and weighed; fragmentary shells were analyzed to yield minimum number of individuals (MNI) estimates based on the identification of non-repeating shell features (see Mason et al. 1998:307).
Results
Spongiidae Microfossils
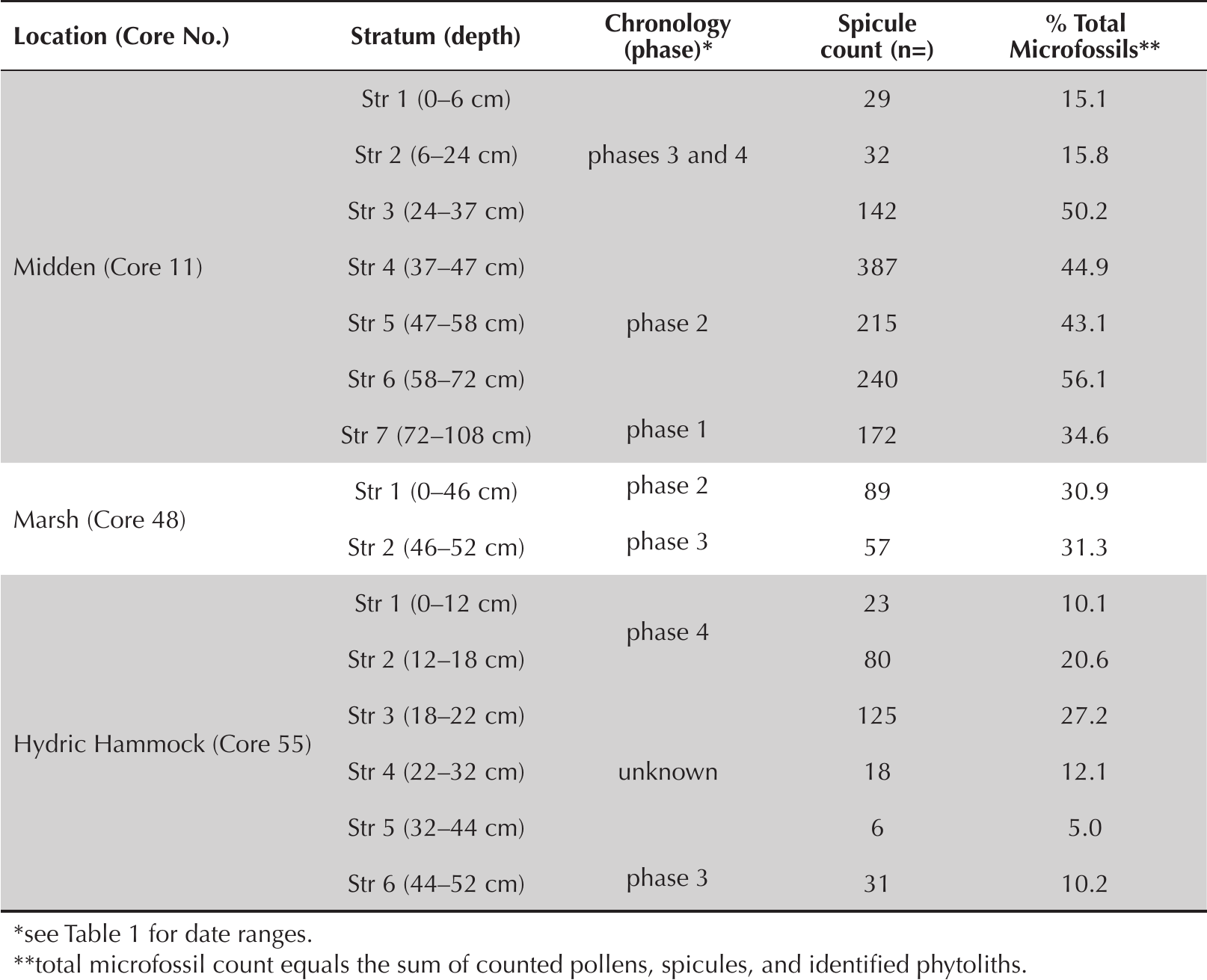
At the onset of this study, we expected spicule concentrations in wetland soil samples (Cores 48 and 55) to be considerably greater than concentrations within shell midden strata (Core 11). However, spicules are present in considerably greater relative abundance within midden soils (Table 2) and, on average, spicules are twice as prevalent in midden strata (37.1% of total microfossils) than they are in wetland core strata (18.4% of total microfossils). These comparisons suggest that spicules accumulated over time within midden soils at greater rates than they are naturally produced and deposited in local freshwater wetland habitats. We attribute this signature to the anthropogenic transport of wetland plants or plant elements to activity areas atop the midden ridge where they were cleaned and otherwise processed, resulting in the deposition of wetland mud and sponge inclusions. Alternatively, this microfaunal signature may have been produced through the harvesting and transport of wetland soils themselves for various potential uses, including the manufacture of daub, water-tight basketry, and ceramics. See Lollis and colleagues (2015) and Bloch and colleagues (2019) for experimental and analytical work verifying the use of wetland soils in the production of Precolumbian ceramic pastes in Florida.
Small-Gastropod Taxa
Analyses of invertebrate zooarchaeological assemblages suggest that small gastropods occur with relatively high spatial and temporal ubiquity within shell midden deposits at Crystal River. Among these taxa (Table 3) are 14 species associated with herbaceous wetland plants and several small snails (e.g., Cerithridea sp., Littorina irrorata, Melampus bidentatus, Neritina reclivata, Olygyra sp., Polygyra sp., Prunum apicinum) that depend directly on coastal marsh and swamp understory plants for forage and structure (Figure 4). For example, eastern melampus snails (M. bidentatus) are well-documented wetland forb-obligates that escape UV desiccation hidden at the bases of marsh plants and avoid submersion and predation by climbing stems (Capaldo 1983; Heard 1982:19–20). Supplementary Table 2 displays aggregated MNI data for small gastropods organized by chronological phase at Crystal River. Invertebrate zooarchaeological assemblages across midden contexts at Crystal River are dominated by oyster shells—accounting for between 75 and 89% of invertebrate MNI by chronological phase. However, small wetland-associated snails, together, make up a sizable component of the invertebrate assemblage (between 9 and 14% MNI) across the site's occupational history. This is despite considerable sampling biases and high rates of fragmentation that predispose small gastropod taxa to under-representation in zooarchaeological analyses.
Abundance and relative abundance data for sponge spicules in midden and wetland core samples from Crystal River (8CI1).
Microbotanical Evidence
Palynological analyses of midden soils in Core 11 from the Crystal River site document the representation of various edible, technologically useful, and ethnohistorically documented flora (Table 4). At this point, we refrain from interpreting palynological evidence of wind-pollinated plants as cultural residues. While many of these taxa, such as cattails and southern red cedar (Juniperus silicicola), represent important ethnobotanical resources of known dietary, economic, and cosmological significance, the presence of their pollen in terrestrial soils is not particularly suggestive of cultural use. The midden pollen assemblages also contain numerous entomophilous/zoophilous taxa with pollen dispersal patterns limited to areas directly below vegetation stands and locations where plant materials or associated soils were stored, processed, and/or utilized. These include a set of conspicuous, ethnobotanically known herbs that grow in fresh- to brackish-water wetlands (Figure 5a–d). Together, these palynological remains support the proposition that ancient peoples transported wetland flora and/or wetland soils for use in village activity areas; the identities of these plants enable us to propose the potential harvesting, processing, and utilization of particular floral resources that, to date, have escaped archaeological recovery.
Taxonomic and ecological information for small gastropods identified in Crystal River (8CI1) midden deposits.
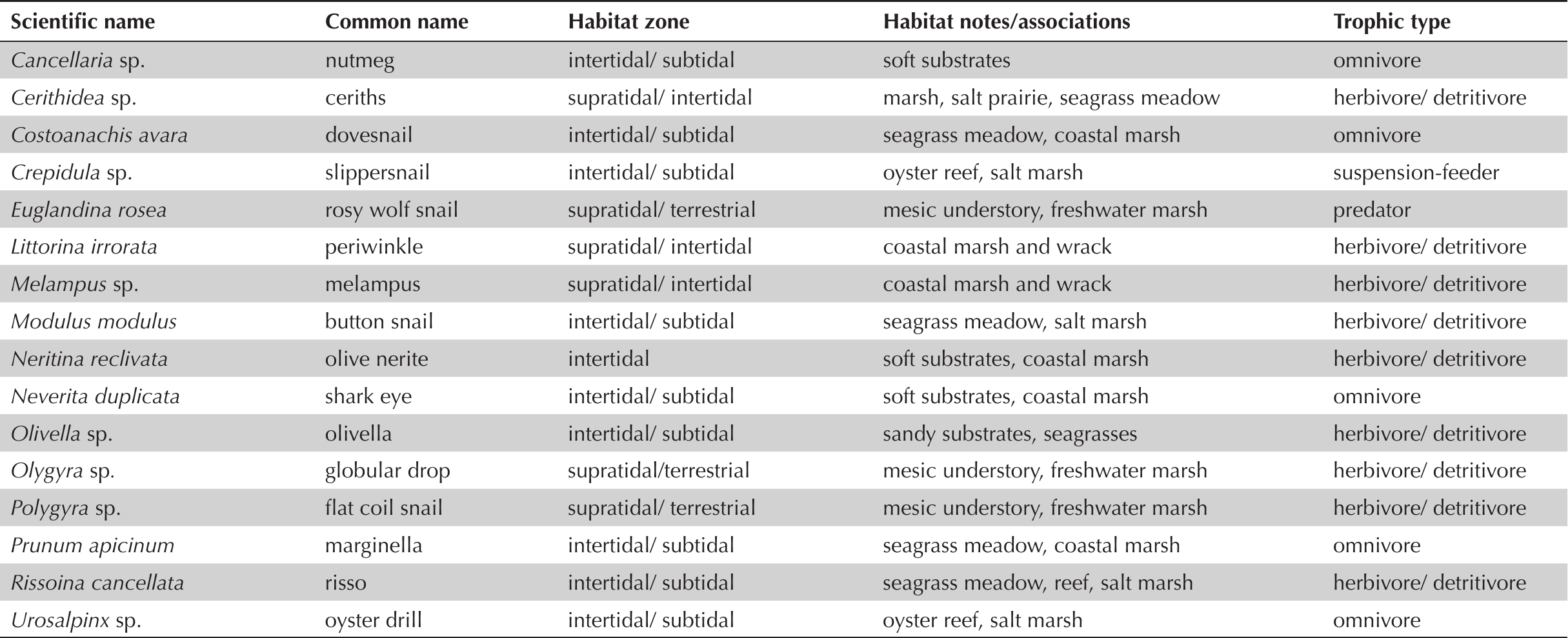

Archaeological examples of wetland-associated small gastropods identified in this study: a. Cerithium muscarum; b. Crepidula sp.; c. Polygridae; d. Urosalpinx tampaensis; e. Neritina reclivata; f. Euglandina rosea; g. Melampus bidentatus; h. Littorina irrorate; i. Prunum apicinum.
Wapato (Sagittaria sp.)
Wapato (Sagittaria sp.) is a widely distributed emergent wetland herb inhabiting freshwater marshes, swamps, lake margins, stream banks, and ditches (Figure 5a) (Judd et al. 2016:266–268). It is represented throughout the midden record at Crystal River (AD 69 to 1060), making up between 3 and 12% of total pollen counts (Jackson and Pluckhahn 2020:66). Mature wapato plants prefer deep mud and ample sunlight, but can thrive in areas with widely ranging temperatures, hydroperiods, and water depths (Thayer 2006:101–116). Indeed, wapato is among very few obligate wetland forbs that remains prevalent within the Crystal River pollen record across the highly variable, but predominantly cold and arid, Vandal Minimum climatic episode, ca. AD 600–800 (Jackson and Pluckhahn 2020).
Wapato is a well-documented ethnobotanical resource among Indigenous societies across North America (Darby 2005; Garibaldi 2003; Sturtevant 1955:298). Mature wapato stands produce crops of edible starchy underground tubers that have been intensively cultivated, harvested, and exchanged as an important geophyte food resource among First Nations peoples of the Pacific Northwest for millennia (Darby 2005; Hoffman et al. 2016). Garibaldi and Turner (2004) argue that wapato constitutes a cultural keystone species—taxa crucial to the operation of both local ecosystems and sociocultural and economic systems—among historic Pacific Northwest First Nations peoples. Darby (2005) traces the complex process of wapato intensification and its role in historical land-ownership practices and exchange networks among the Chinookan People of the Lower Columbia River valley. Wapato is also well documented as a Native American medicinal resource. The Potawatomi, Lakota, Iroquois, Chippewa, Cherokee, and Seminole have utilized various parts of the wapato plant in topical medicines for wounds, sores, and boils, as well as in gastrointestinal, antirheumatic, and laxative aids (Hamel and Chiltoskey 1975:23; Mechling 1959:124; Rogers 1980:26; Sturtevant 1955:298).
Ethnobotanical resources identified in midden pollen assemblages at Crystal River (8CI1).
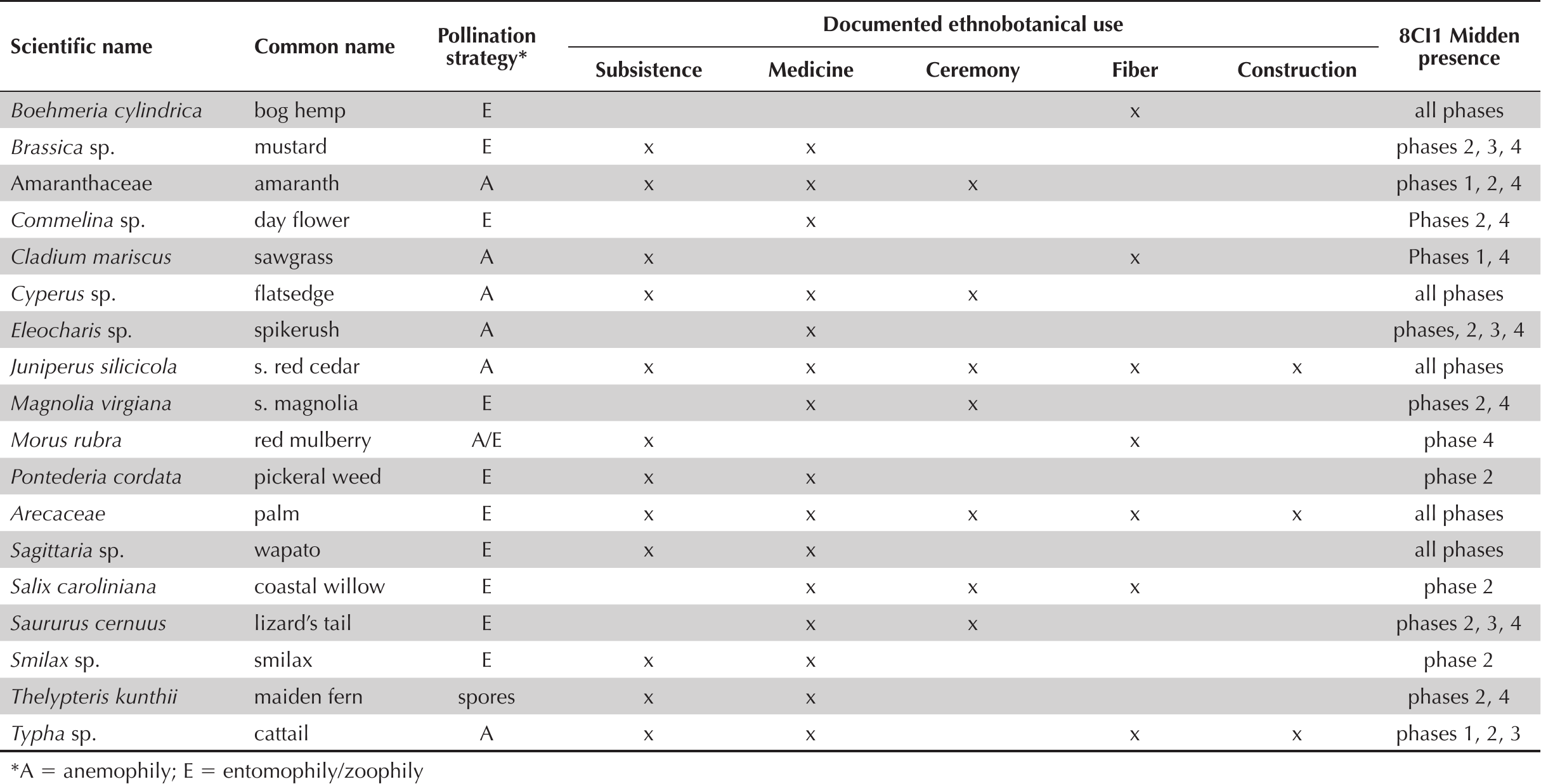
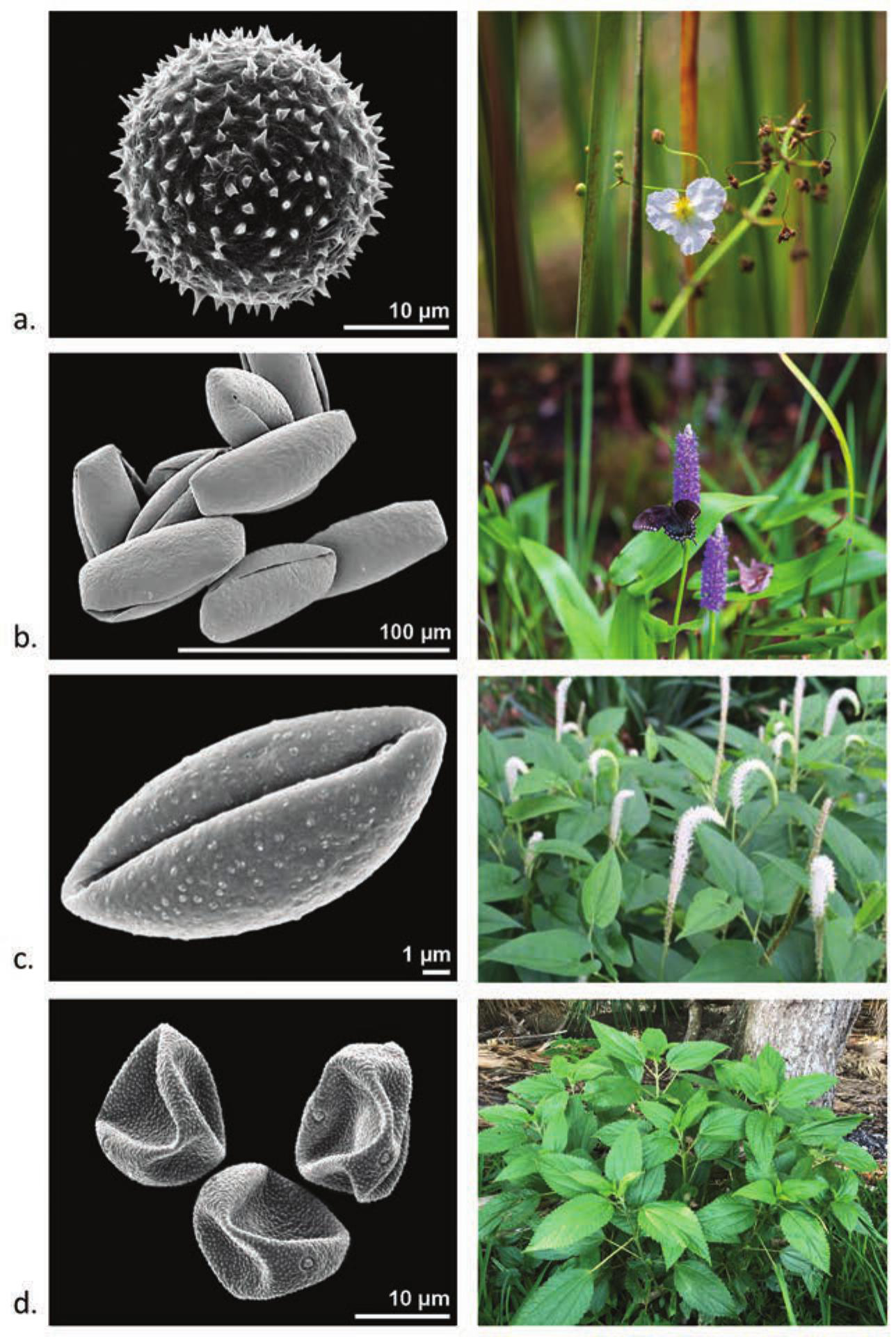
Pollen grains and plant habits for taxa discussed in text; a. Sagittaria latifolia; b. Pontederia cordata; c. Saururus cernuus; d. Boehmeria cylindrica. Scanning electron micrographs by Halbritter (2016). Photography by Emily Keen.
Pickerel Weed (Pontederia cordata)
Pontederia cordata, or pickerel weed, is found in marshes, swamp understory, ponded areas, and along stream banks (Figure 5b) (Judd et al. 2016:298–300). It is represented at Crystal River in midden strata deposited during the florescence of site activity (AD 221–544), where it accounts for 5% of the total pollen assemblage (Jackson and Pluckhahn 2020:66). The North American historical ethnobotany of pickerel weed is not well developed; however, Indigenous groups in the Northeast are known to have harvested and utilized its corms as a food resource and other elements as medicinal resources (Chandler et al. 1979:59; Mechling 1959:259; Speck 1917:315).
Lizard's Tail (Saururus cernuus)
Saururus cernuus, or lizard's tail, is found in swamp understories, and along stream banks and lake margins (Figure 5c). It can tolerate a relatively wide temperature range but is always associated with surface-water (Hall 1940). Lizard's tail is represented in Crystal River midden deposits from AD 221 through AD 1000—making up between 5 and 15% of total pollen counts—and is best represented during the floresence of village activity (AD 221–544) (Jackson and Pluckhahn 2020:66). Its documented ethnobotanical uses are predominantly medicinal and ceremonial. It is used in topical poultices for wounds, in medicinal washes/baths, within internal medicines as a mild tranquilizer, and in ceremonial emetics taken during ritual fasting (Austin 2003; Hamel and Chiltoskey 1975:43; Morgan and Perry 2010; Sturtevant 1955). Snow and Stans (2001) record its use in Seminole/Miccosukee herbal medicines that are ingested and bathed in, including use as a death medicine and as treatment for stroke, excessive sleep, and bad dreams. Experimental analyses have shown that the chemistry of lizard's tail lends itself to medicinal use; it contains antiseptic and antitumor compounds, as well as antioxidant, antiviral, and sedative compounds (Rao and Rao 1990; Rao and Reddy 1990).
Bog Hemp (Boehmeria cylindrica)
Boehmeria cylindrica, or bog hemp, inhabits swamp understories, marshes, bogs, wet meadows, and ditches (Figure 5d) (Judd et al. 2016:392–393). It is well represented across the midden record at Crystal River (AD 69–1060), and is especially prevalent in midden strata below 24 cm depth, where it makes up between 30 and 70% of total pollen counts (Jackson and Pluckhahn 2020:66). Bog hemp and other members of the nettle family are well known sources of strong, fine-diameter fiber (Judd et al. 2016). Whitford's (1941) examination of various ethnohistoric documents and museum artifacts found that nettle family herbs were selected as a prominent source of cloth and cord fiber among historic and pre-contact southeastern Native American communities. In the Pacific Northwest, nettles are well documented ethnobotanical resources that were widely transplanted, cultivated, and processed to make fishing nets, lines, mats, and various other cordage-products (Turner and Peacock 2005). In the same region, large patches of stinging nettle (Urtica dioca) are still found outside their natural range, proliferating in low-lying cultural soils surrounding historic and pre-contact coastal village sites (Turner 2014).
Palm (Arecaceae)
In addition to fossil pollens and sponge spicules, palm (Arecaceae) phytoliths are differentially preserved and well represented across all phases of midden formation at the Crystal River site (ca. AD 69 to 1060). These microfossils account for between 29 and 57% of total microfossil counts and clearly indicate the local deposition of palm elements (Jackson and Pluckhahn 2020). The most likely taxa include cabbage palm (Sabal palmetto) and saw palmetto (Serenoa repens); both of these plants grow prolifically at the Crystal River site today and are well documented in the ethnobotanical and paleoethnobotanical records from the peninsular Gulf Coast as important resources for house building and fiber-craft (Baber 1942; MacCauley 1887; Newsom and Scarry 2013).
Discussion
The results of our analyses support the proposition that herbaceous wetland plants were utilized by ancient Native American peoples at Crystal River across the first millennium AD. We argue that the harvesting, processing, and use of these floral resources represents the most parsimonious explanation for the accumulation of small, wetland-associated gastropods, invertebrate microfossils, and fossil pollen from zoophilous wetland forbs. Further, palynological remains suggest that a set of particular coastal wetland plants were utilized as dietary carbohydrates, medicines, and raw materials for fiber-craft. Below, we discuss social and political implications of Precolumbian wetland plant use at a coastal mound complex that functioned as a node for civic and ceremonial aggregation and a regional epicenter of village coalescence. We then place this exploratory study into broader methodological context and propose avenues for future research.
In recent investigations at Crystal River, Robert's Island, and several other Woodland-period (ca. 1000 BC–AD 1000) mound centers on Florida's north-peninsular Gulf Coast, archaeologists have reconstructed traditions of faunal estuarine resource intensification tied to ceremonial feasting, mound-building, mortuary interment, and world-renewal (Duke et al. 2020; Goodwin et al. 2019; Sampson 2015; Sassaman et al. 2020; Wallis and McFadden 2020). While these studies emphasize mass-captured faunal resources, namely oyster and small estuarine fishes, the evidence reported here justifies further archaeological consideration of wetland geophytes as important risk-reducing subsistence resources. The biology and ecology of wapato predisposes it to intensification. Wapato reproduces rhizomatically and can grow rapidly in dense monoculture, reaching approximately 30 plants per square meter in mature stands (see Darby 2005). It has a long season of harvest (October through April), and tubers can be collected rapidly without considerable effort or specialized technology. Further, due in part to a co-evolutionary relationship with migratory waterfowl, disturbance and harvesting in wapato stands encourages further proliferation and expansion—as these activities facilitate the multiplication and spread of rhizome and tuber fragments from which new plants emerge (Darby 2005). While less published research is available concerning the propagation and harvesting of pickerel weed and other starchy coastal wetland forbs (e.g., cattails), it should be noted that these plants also reproduce rhizomatically and are well suited for intensive utilization as subsistence resources (Morton 1975; Nicholas 1998). Taking a broad view, Nicholas (1998:725) notes that wetland systems represent accessible and relatively stable environments for procuring resources, which may explain close geographic and temporal associations between productive wetland ecosystems and the development of dense human societies with reduced residential mobility and systems of territoriality (c.f., Grier 2014; Marquardt 1992).
Geochemical analyses on oyster shell from well-dated contexts at Crystal River (Lulewicz et al. 2017) and Garden Patch (Lulewicz et al. 2020) suggest that civic-ceremonial aggregations and mound building events took place during cool-season months that are historically associated with regional harvesting times for oyster and mullet (Mugil sp.). We note that cool-season aggregations would have also been timed appropriately to harvest wapato, as well as other rhizomatic wetland herbs. The close ecological associations between wetland geophytes and migratory waterfowl should also not be overlooked by Gulf Coast archaeologists, particularly given the prominence of wading birds in Woodland-period mortuary iconography (e.g., Donop 2017; Spivey-Faulkner 2018) and the incorporation of waterfowl remains within ritual pit features at north peninsular Gulf Coast mound centers (Goodwin et al. 2019).
Archaeological discussions of estuarine resource intensification, ceremonial aggregation, and village formation on the Florida Gulf Coast have recently reinvigorated investigation of ancient Native American mass-capture technologies first hypothesized by Cushing ([1897] 2000) (Mahar 2019; Sassaman et al. 2020; Thompson et al. 2020). Analyses of vertebrate zooarchaeological assemblages at Crystal River/Roberts Island (Duke et al. 2020), Shell Mound (Sassaman et al. 2020), and other north peninsular Gulf Coast sites (e.g., Jenkins 2019) have emphasized the dietary and economic importance of demersal and schooling pelagic fishes that would have been most efficiently harvested with trap and net technologies. Ethnohistoric accounts (Hann 1991:311), technological studies (Walker 2000), and rigorous experimental-archaeological work by Mahar (2019) attest to the wide-spread use of nets by Indigenous Gulf Coast peoples. However, few investigations have presented material remnants of net manufacture, and fewer have sought evidence for the cultivation or sourcing of raw materials for fiber crafting (Newsom et al. 2013). At Crystal River, the high ubiquity and conspicuous abundance of bog hemp pollen within the midden record likely represents the remnants of fiber crafting practices that produced nets, lines, basketry, and other cordage goods. Palm trees have been cast as principal sources of cordage fiber on the Gulf Coast and, indeed, the widespread use of palm as a source of fiber, dietary carbohydrates, and construction material seems likely. Cordage made from palm fiber has been identified at the Pineland complex in southwest Florida (Newsom et al. 2013) and palm phytoliths are both abundant and ubiquitous within Crystal River's midden deposits. However, in addition to palm, we propose that nettle family herbs should be examined further by archaeologists as important sources of craft fiber on the northern Gulf Coast. Nettles, like bog hemp, grow rapidly in wet soils and form dense perennial stands that regenerate adventitiously from stem and root tissues (see Turner and Peacock 2005). These characteristics predisposed nettles to cultivation, transplanting, and intensive use by First Nations peoples of the Pacific Northwest Coast and position bog hemp as an ideal source of fiber for Gulf Coast communities in need of large, reproducible quantities of cordage for net manufacture and repair.
Direct evidence for the importance of herbaceous wetland plants in ancient Native American and First Nations lifeways has been amply demonstrated by archaeologists working within water-logged, anaerobic contexts, large carbonized features, and xeric/rock shelter sites (Supplementary Table 3). In the absence of these ideal conditions, paleoethnobotanists (particularly in Mesoamerica) have turned to starch grains and phytolith assemblages to reconstruct the use of floral resources (Berman and Pearsall 2008; Messner 2011; Piperno et al. 2009). Within more challenging environments of botanical preservation, where starch grains and phytoliths are readily degraded, such as the Florida Gulf Coast, ancient traditions of wetland plant use have been rendered invisible within the archaeological record and largely neglected in reconstructions of Precolumbian lifeways. Our interpretation of small mollusk assemblages as incidental bycatch follows Ainis and colleagues' (2014) analysis of archaeological deposits on the San Miguel and San Nicolas Islands, California, where small gastropod assemblages appear to represent the persistent remnants of marine macrophyte harvesting across much of the Holocene. We have aimed to strengthen this bycatch-focused analytical approach by integrating zooarchaeological data with analyses of invertebrate microfossils and palynological remains that together implicate the harvesting, processing, and use of herbaceous coastal wetland plants.
In this study, we recognize the material signatures of ancient wetland plant use by analyzing classes of environmental-archaeological data that are small enough to be overlooked in conventional archaeological practice. Following Deetz (1996: 259–260), we argue that “seemingly little and insignificant” archaeological materials may represent all that remains of past behaviors that were once of immeasurable cultural importance. Moving forward, microbotanical analyses and biochemical work on targeted archaeological contexts—such as roasting pits, ceramic vessel interiors, and inundated midden deposits—are necessary to further elucidate patterns of ancient plant use on the Florida Gulf Coast and, by extension, other coastal areas of North America where Indigenous plant use has been understudied by archaeologists. Further analyses of environmental-archaeological proxy datasets may improve upon the present study through quantitative analyses that incorporate factor reduction techniques (e.g., CA, PCA, DCA) to more rigorously isolate and explain material signatures. Future inquiries may also compare plant use signatures between inland and coastal sites to investigate specialization and exchange or between site features to determine how particular types of plant resources were processed and utilized in the ancient past.
Footnotes
Acknowledgments
This study would not have been possible without the cooperation and assistance of Dr. Thomas Whitmore and Dr. Melanie Riedinger-Whitmore of University of South Florida, St. Petersburg, Department of Biology. We thank Dr. Dana Lepofsky, Dr. Natasha Lyons, and two anonymous reviewers for providing thoughtful and insightful comments and suggestions that greatly improved the quality of this article. Funding was granted by the Society for American Archaeology Geoarchaeology Interest Group and the Florida Paleontological Society.