
Abstract
Food habit studies in the Amazon emphasize the consumption of fish and manioc (Manihot esculenta) flour as basic diet in different ethnic groups, while little is known about human plant-related diet dynamics during the hydrological regime. In a scenario of food transition in rural Amazonia, with insertion of industrialized items in the diet, traditional food and consumption of regional products have undergone transformations, affecting the autonomy and lifestyle of traditional populations. Considering this, the objectives of the current case study were: (1) to contextualize plant-related eating habits in five Riverine communities on the Lower Purus River and (2) to characterize the dietary dynamics of plant eating according to the hydrological regime, considering the provenance of food items. We conducted interviews and sample collection in both low- and high-water seasons. We calculated species richness and diversity per meal and season and used multivariate analysis to access differences in plant consumption between seasons. The greatest richness and diversity of plants is consumed in the flooding season and mainly as snacks, the meal most susceptible to being replaced by industrialized foods. Despite a significant difference in diet between seasons, the basic array of plants consumed is similar in both seasons, with availability topped up by external purchases. This tendency deserves attention and action from public policies aimed at providing food security in the region.
Introduction
The study of eating habits includes patterns of food selection and consumption and the factors that influence these behaviors, such as seasonal resource availability. For human societies, the context of agricultural production or socioeconomic status can provide important information about relationships between different cultures and the social and biological environment (Dufour and Teufel 1995). Eating habits in a globalized society commonly show a progressive departure from food knowledge and increasingly rare consumption of traditional and regional cuisine with an enhanced consumption of processed foods, all of which impact the nutritional status of populations (Cerda 2014; Hernández 2005; Pons 2005).
In recent decades, the diet of people in remote regions, such as rural Amazonia, have undergone changes as integration of such populations into wider markets increases. Traditional foods have been partly replaced by industrial “similars” in a process of “food transition” (Batista and Rissin 2003; Nardoto et al. 2011; Piperata 2007; Popkin 1993; Silva et al. 2017). The literature on food in the Amazon revolves around the basic binomial of fish and manioc (Manihot esculenta) flour (Adams et al. 2005; Alencar et al. 2002; Murrieta 2001; Murrieta and Dufour 2004; Murrieta et al. 2008), while vegetables, spices, and fruits are relegated to the category of “not food” (Murrieta 2001). It has been proposed that fruits are mainly consumed seasonally (Murrieta 2001), according to the fruiting cycle of forests in the wet season (Haugaasen and Peres 2005); however, there are still few studies that emphasize the plant diversity consumed by Amazonian populations, despite the remarkable contribution that plants actually make to their diets (Dufour et al. 2016).
Since pre-Columbian times, the Amazon has been the origin of many domesticated plant species, such as manioc (Manihot esculenta), cocoa (Theobroma cacao), pupunha (Bactris gasipaes), and Brazil nut (Bertholletia excelsa) (Clement et al. 2015). Recent studies have revealed a strong relationship between the occurrence of these domesticated species and that of archaeological sites and Terra Preta de Índio (TPI) sites, showing both the transformative effect of human societies on the Amazonian landscapes and the importance of these species in the diet of these populations throughout their existence (Clement 1999; Junqueira et al. 2011; Levis et al. 2017).
Currently, food security in the Amazon is threatened by such factors as the instability of manioc production in flooded regions, the process of food and nutrition transition, and environmental seasonality (Dufour et al. 2016). For example, the difference in navigability of rivers where water levels show great seasonal variation directly influences the access by rural communities to industrialized products and, consequently, eating habits and food costs. In addition, availability of fish and farmland also varies according to river flood pulses, so affecting eating habits, especially for populations inhabiting flooded forest areas (Costa 2015).
The concept of food sovereignty developed in Brazil by rural social movements embraces the decision-making power on food production, distribution, and consumption strategies, and also the right to cultural perpetuation of food habits (CONSEA 2006). Schor et al. (2015) contribute to the discussion of food security in Amazonia with the concept of food sovereignty, which focuses on food becoming more dependent on external resources, weakening ties with local resources and traditional cuisines.
In order to better understand food dynamics and potential threats to food security and sovereignty in Central Amazonian communities, the objectives of the current study were: (1) to contextualize plant-related eating habits by assessing the food transition process in communities on the Lower Purus River, Central Amazon, and (2) characterize the dietary dynamics of plant-eating according to the hydrological regime, in the high- and low-water seasons, considering the provenance of food items.
Methods
Study Area
The Piagaçu-Purus Sustainable Development Reserve (RDS-PP: Reserva de Desenvolvimento Sustentável Piagaçu-Purus) is located on the Lower Purus River, Amazonas state, Brazil, about 24 hours by boat from the state capital, Manaus. The region includes a wide variety of environments, with portions of non-flooded terra firme forests and floodplain environments of different origins, such as várzeas (sediment-rich floodplains), igapós (sediment-poor floodplains), and lakes (Piagaçu Institute 2010). Much of the RDS-PP is covered in alluvial dense ombrophilous forest and lowland dense ombrophilous forest, with 45% of its area occupied by várzea. The climate in the region is tropical rainy, with constant high temperatures and two well-defined seasons: rainy (February to June) and dry (July to October) (Haugaasen and Peres 2006). Annual precipitation in the Purus River basin ranges from 1550 to 3350 mm, and Purus River annual water-level variation may reach 12 meters. (Haugaasen and Peres 2006; Piagaçu Institute 2010).
Since the early twentieth century, the reasons that attracted non-Indigenous populations to occupy the Purus River region were related to the extraction of latex from rubber trees (Hevea brasiliensis), fishing, especially pirarucu (Arapaima gigas), collection of Brazil nuts (Bertholletia excelsa), cultivation of jute (Corchorus capsularis) and mallow (Urena lobata), and the hunting of animals for the international fur and skin trade (Antunes et al. 2014; Piagaçu Institute 2010). Currently, fishing and nut extraction are the most economically important activities for the human population in the region (Piagaçu Institute 2010).
Within RDS-PP, the local non-Indigenous population (Ribeirinhos) is around 4000 people, distributed between 57 communities located in or around the reserve boundaries (Piagaçu Institute 2010). The reserve surrounds two demarcated Indigenous lands, occupied by the Mura and Apurinã ethnic groups, which are not part of the reserve, but are within its limits (Piagaçu Institute 2010).
Sample Design and Data Collection
Five Riverine communities were invited to participate in the research program, all located in the northern part of the reserve (Figure 1). To maximize the diversity of environments studied, we chose communities located in both várzea and terra firme, as well as those in intermediate situations, where the two environments were both important in community life.
Field research was conducted between September 2016 and November 2017 and totaled 133 field days. Visits were made at different times of the year, first to become acquainted with the study area, the residents, and work-related logistics (September and March), then at the peak of the Purus River high- and low-water seasons (June and November, respectively). Survey participants were selected through probabilistic sampling (Albuquerque et al. 2010), where a minimum of 20% of the total households were drawn for each of the five communities. All residents of the selected households were invited to participate, including children, and each participant was interviewed once each season (high- and low-water).
For the eating habit survey, the 24-hour recall method (Bernard 2006; Dufour and Teufel 1995; Zent 1996) was used only for consumed plants in their natural form (“in natura”) or minimally processed, such as cleaning processes, removing inedible or undesirable parts, fractionation, grinding, packing, drying, fermentation, and similar processes that do not involve aggregation of other substances to the original food. Processed and ultra-processed foods with ingredients of plant origin were excluded. To address these cases, we interviewed the elderly about the food transition process. For this classification, we take as reference the official Brazilian document “Food Guide for the Brazilian Population” (Ministério da Saúde de Brasil 2014). The frequency data on consumption of plants was calculated according to the times they were mentioned in the interviews, and the total number of different plants consumed was extracted from a distinct count of all citations. Species referred to during interviews were subsequently collected on site visits accompanied by a local informant (Alexiades 1996) and taxonomically identified via expert consultations and examination of specimens in physical (INPA) and virtual herbarium (Reflora 2019; Missouri Botanical Garden 2019).
To gather information on past eating habits and the food transition process, long and detailed free-form interviews were conducted with seven elderly residents living in communities since childhood. Based on the reported dates and life-lines, it is probable that this memory-base dates back to approximately 1945.
Data Analyses
The provenance of consumed food was categorized as: external purchase (purchase of items produced in other communities or in the urban center) and agroextractivism (community agricultural production or extractivism). We used the following categories to characterize the diet in terms of consumed plant parts: bark (or trunk), fruit, seed, tuber, and leaf. We categorized the preparation form as: baked, beverage, boiled, flour, fried, “in natura” (fresh), spice, and sweets. The proportion of items consumed in relation to plant part and type of preparation was compared between four meal classes: breakfast, lunch, snack, and dinner.
To describe the diversity of consumed plants (Begossi 1996), we calculated the species richness (i.e., number of “plants” cited) and Fisher's alpha diversity per meal type, for both flooding seasons (high- and low-water). We used a chi-square test to assess if frequencies of items consumed differ in relation to provenance (i.e., external purchase or agroextractivism) between the high- and low-water seasons (Zar 1999).
For an analysis of consumption at the household level, we considered the sum of all plants mentioned in the 24-hour recall by each member of the houses, in each season separately. In order to compare the similarity of plant consumption at the household level in high- and low-water seasons, we used a Non-Metric Multidimensional Scaling (NMDS) analysis. For this, a Bray-Curtis Index was used to generate a matrix of floristic dissimilarity for high- and low-water seasons from the per household plant abundance consumption data. To test the hypothesis that household diet composition is different between water-level seasons, a permutation multiple variance analysis (np-MANOVA) was applied from the dissimilarity matrices calculated by the Bray-Curtis method, using the function “adonis” from the vegan package (Oksanen et al. 2009). The number of “plants” or “ethnospecies” (not number of species) was used since consumption may involve “plants” that correspond to the same botanical species, but which are culinarily distinct, for example Manihot esculenta prepared as flour (toxic mandioca) or boiled (non-toxic macaxeira). In this way, the data analyses were carried out considering the cultural definition of culinary plants (here called “plants” or “ethnospecies”) and not the scientific definition of species (Appendix 1). All analyses were performed using the R Development Core Team 2019 program using the vegan package (Oksanen et al. 2009; R Development Core Team 2013).
Ethical Aspects
The research project was approved by the Research Ethics Committee of the National Institute of Amazonian Research (CEP/INPA) (Certificate of Presentation for Ethical Appraisal [CAAE] number 68477417. 5.0000.0006). A Prior Consent Form and the Free and Informed Consent Form were signed by community leaders and by each interviewee, respectively. Results feedback group activities were conducted with all participating communities in July 2018.
Results
Plant Consumption and Food Context
This study involved 249 residents of five communities, between four and 77 years old, from 70 households (141 females and 108 males). Consumption of 72 plants (ethnospecies) was recorded during the field study period, corresponding to 67 botanical species (Appendix 1). The free-form interviews about the food transition process involved seven elderly people between 63 and 92 years old.
The daily eating habits of the communities start with breakfast as soon as people wake up. This meal can contain anything from a single cup of coffee with industrialized cookies to fried plantains or fish with manioc flour. Lunch is eaten around noon and it is when the most amount of food is consumed. Dinner is just after dark and snacks (merenda) take place between these main meals, but mainly between lunch and dinner, in the afternoon.
The greatest variety of consumed plants was recorded in the snack category, followed by lunch and dinner, with breakfast being the meal form with the lowest richness. Proportionally, fruit was the most consumed plant part (all meals summed), though mainly in the snack category (e.g., Theobroma grandiflorum, Psidium guajava, Inga edulis). Leaf and tuber consumption was proportionally highest at lunch and dinner (e.g., Manihot esculenta, Dioscorea trifida, Talinum paniculatum, Coriandrum sativum, Eryngium foetidum). Bark consumption was recorded only for breakfast and tea (as Aniba canelilla), and seed only as snacks (as Bertholletia excelsa and Acioa edulis) (Figure 2).
For consumption frequency, plants were most often fried for breakfast (notably fried plantains), while a highest proportional use of plants as spices was recorded for lunch and dinner (aromatic herbs such as Allium fistulosum, Coriandrum sativum, and Eryngium foetidum) and the highest proportion of “in natura” (fresh) plant consumption occurred as snacks. The latter category was also responsible for the greatest variety of forms of consumed plant preparation (Figure 2).
In terms of the number of plants reported, the greatest recorded numbers were eaten for lunch and dinner (as either spice or flour). That is, although the greater diversity of reported plants was consumed as snacks, lunch and dinner had greater consumption in quantity, though with less plant diversity (Table 1).
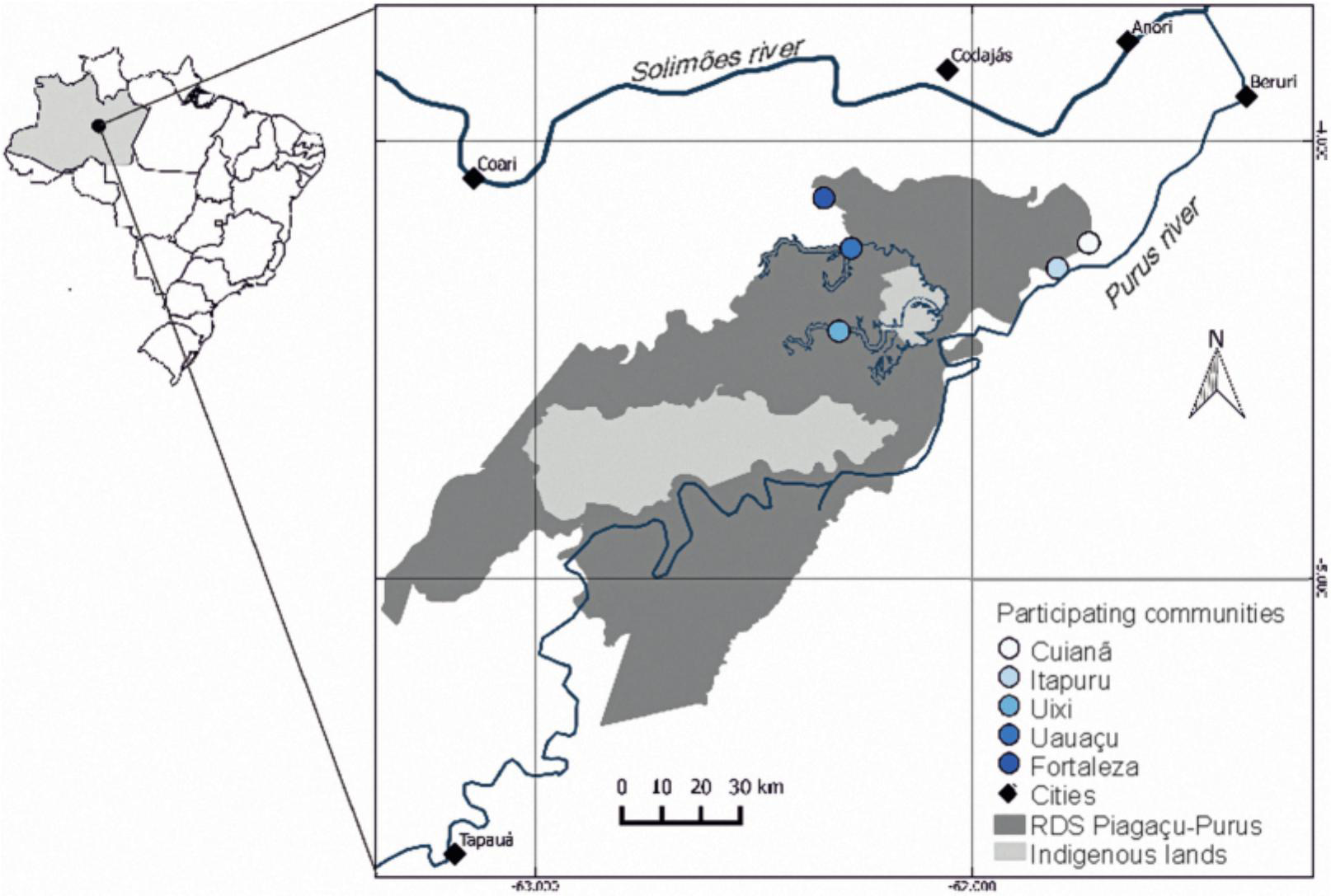
Location of the five communities selected for the study in the Piagaçu-Purus Sustainable Development Reserve.
Plant Consumption and the Hydrological Cycle
Richness and Fisher-a diversity were higher in the high-water season than in the low-water season (61 and 11.5, 49 and 8.2, respectively). Richness and diversity per meal were similar between high- and low-water seasons for breakfast, lunch, and dinner. For snacks, both richness and diversity were about twice as high for the high-water season (39 plants and a 12.7 Fisher-a diversity value, versus 23 plants and a 6.1 Fisher-a in the low-water season).
Plants consumed by households were significantly different between high- and low-water seasons (np-MANOVA, F 5 4.99, p 5 0.001). Ordination analysis (NMDS) showed that household plant consumption tends to be more similar (i.e., households tend to form groups in the graphs) according to hydrological season (Figure 3). These results show that high-water season consumption is broader and more diversified compared to the low-water period, when by-household consumption shows greater similarity. On the other hand, there is a certain overlap between seasons, demonstrating the existence of a staple plant-related diet which does not change between seasons but becomes more diverse during the high-water period (Figure 3). In contrast, during the low-water season, the diet is more restricted and does not differ greatly from this staple diet.
Total number of plants consumed reported, relative frequency, richness (total of different plants reported) and diversity per meal and flooding seasons.
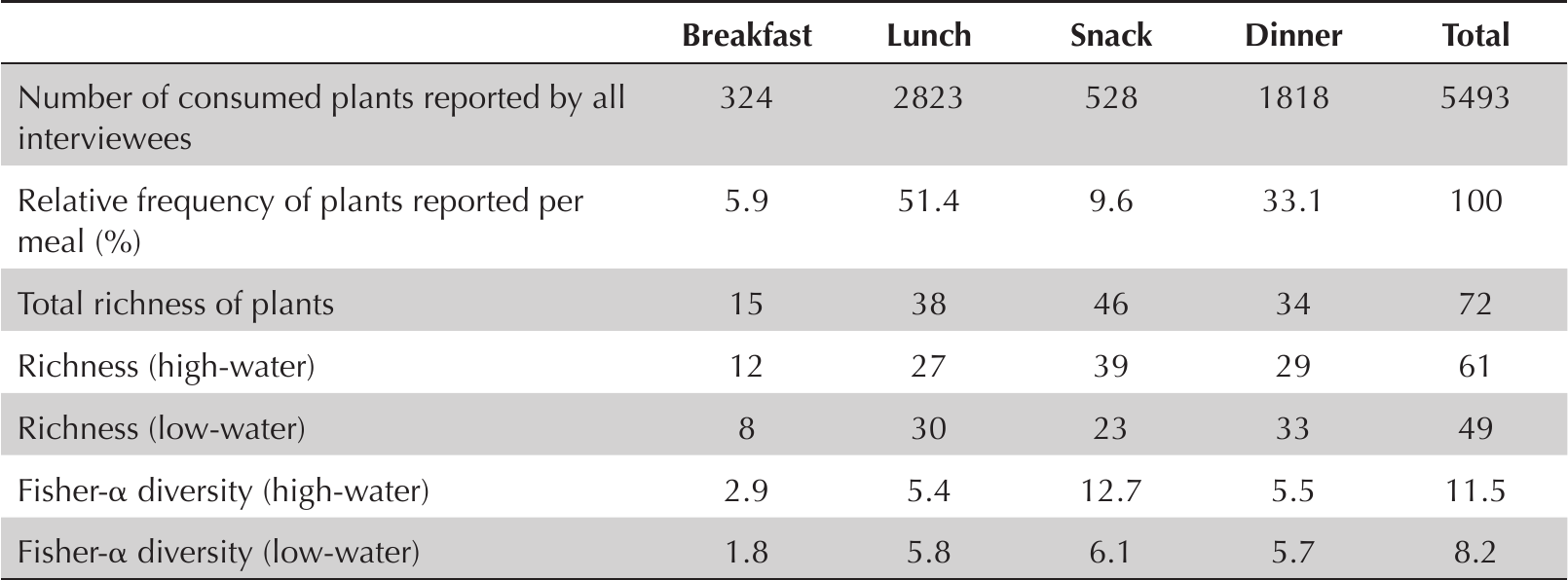
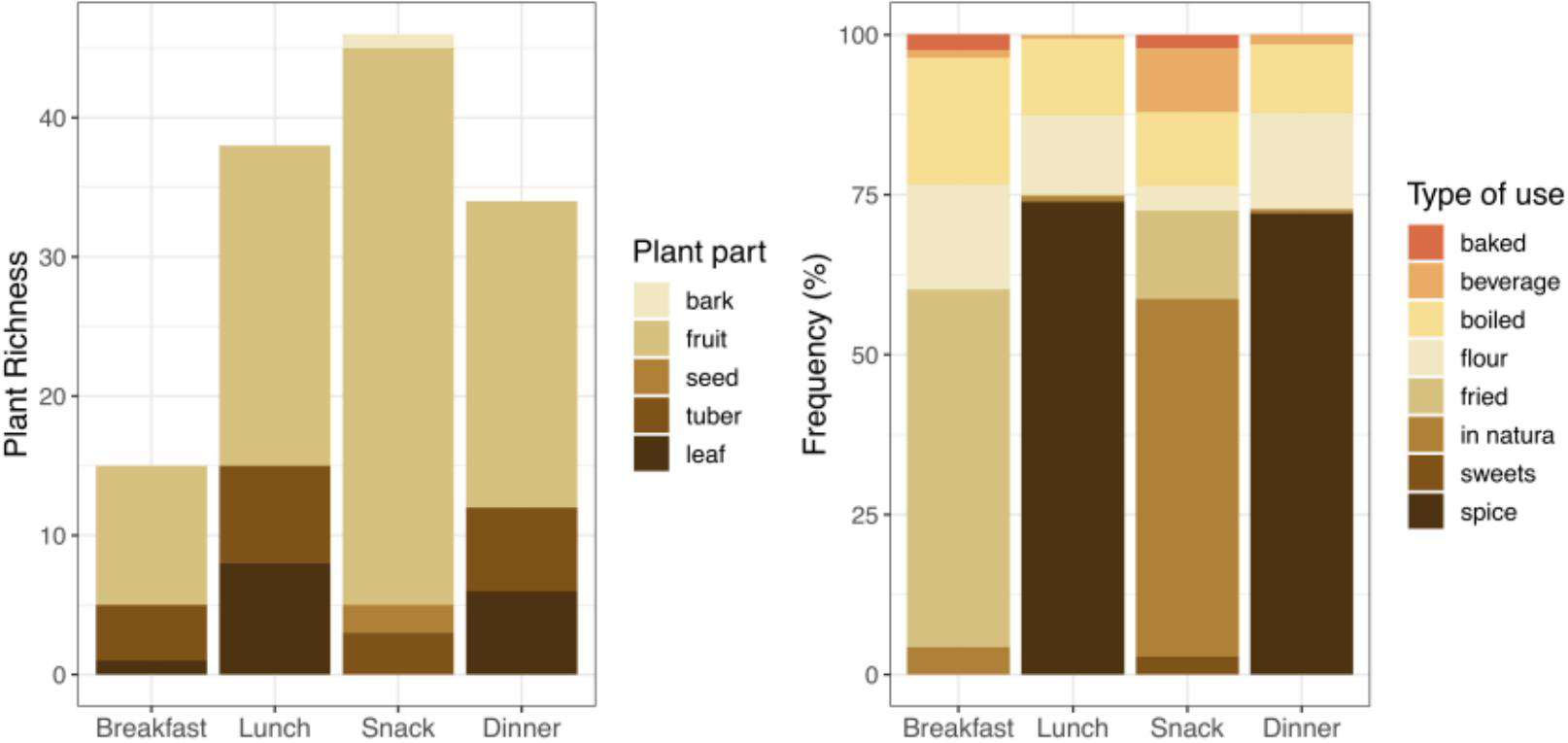
Richness of plants consumed per meal by parts used, and frequency of plant consumption per meal in relation to preparation method.
Qualitative Differences between Seasons
For the staple diet, the daily consumption of plants in all communities is around ten plants (Table 2). The ten most-consumed plants represent 80.3% of total high-water season plant consumption and 76.9% of the low-water season total (Table 3).
When all plant-consumption records in the sample period are included, 61 plants were consumed in the high-season and 49 in the low-water season, with 38 plants in common in both seasons. Thus, even though plant consumption is sporadic and seasonal, most (52.8%) of the plants consumed in high- and low-water seasons are the same. Such plants are mainly cultivated and form part of the staple diet. Although the same basic plants are consumed, their quantity varies between seasons, as a result of seasonal agricultural availability. Plants exclusive to a single season form the smallest part of the diet and are most numerically prominent among those consumed in the high-water season, which are species found in back-yards, agricultural clearings (roçados), and those taken from flooded and terra firme forests. On the other hand, plants consumed exclusively in the low-water season are not derived from forest areas but are cultivated species and obtained either via trade or from local fields (Appendix 1). The eight food plants derived from native forest tree species in flooded forests and consumed exclusively in the high-water season are: Humiriastrum sp., Annona sp., Acioa edulis, Licania hypoleuca and Couepia paraense, Myrcia umbraticola, Bertholletia excelsa, and Aniba canelilla.
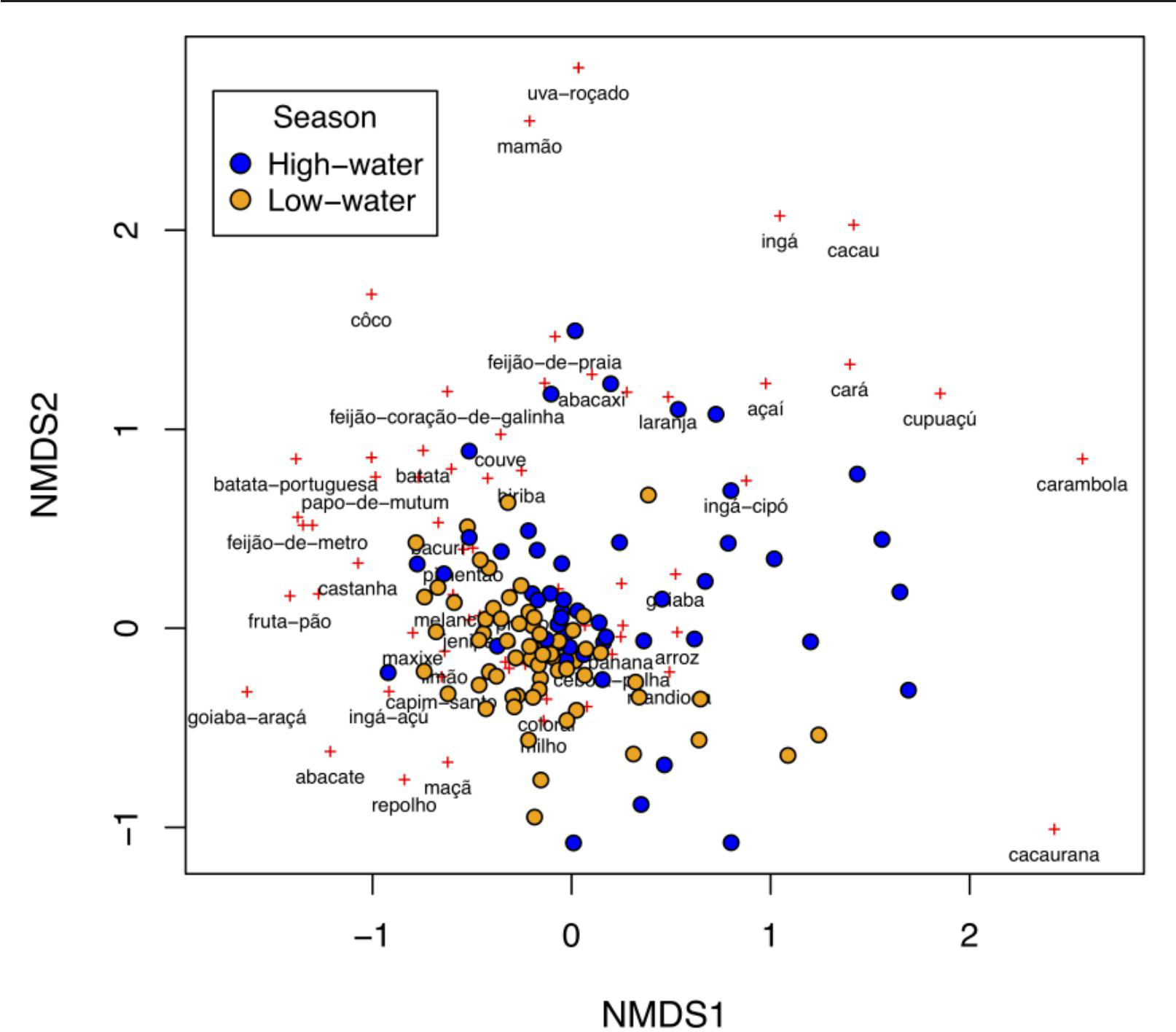
Ordination of samples (interviewed households) using Non-Metric Multidimensional Scaling (NMDS) analysis of between-interviewed households' dissimilarity (Bray-Curtis) matrix for consumed plants, in both high-water (blue points) and low-water (orange points) seasons in the Piagaçu-Purus Sustainable Development Reserve. Red crosses represent the consumed plants (written by the common name) in the ordination analysis.
Provenance of Items Consumed at Different Seasons
When looking at the provenance of plants consumed in the different seasons, there is a seasonal switch in patterns. During high-water, consumption of plants derived from agroextractivism was significantly higher than that from external markets, while in the low-water season, consumption of externally derived plants was significantly higher than those derived from agroextractivism (Pearson's Chi-squared test, x 2 (1) 5 10.2, p 5 0.0014; Table 4).
Mean and standard deviation for total plants consumed by respondents in participating communities.

Relative frequency (%) of the 10 most reported plants in the 24-hour recall and their provenance: agroextractivism (from agriculture or extractivism) and external purchase (produced externally to communities) in high-water and low-water seasons in the Piagaçu-Purus Development Reserve Sustainable, Amazonas, Brazil.
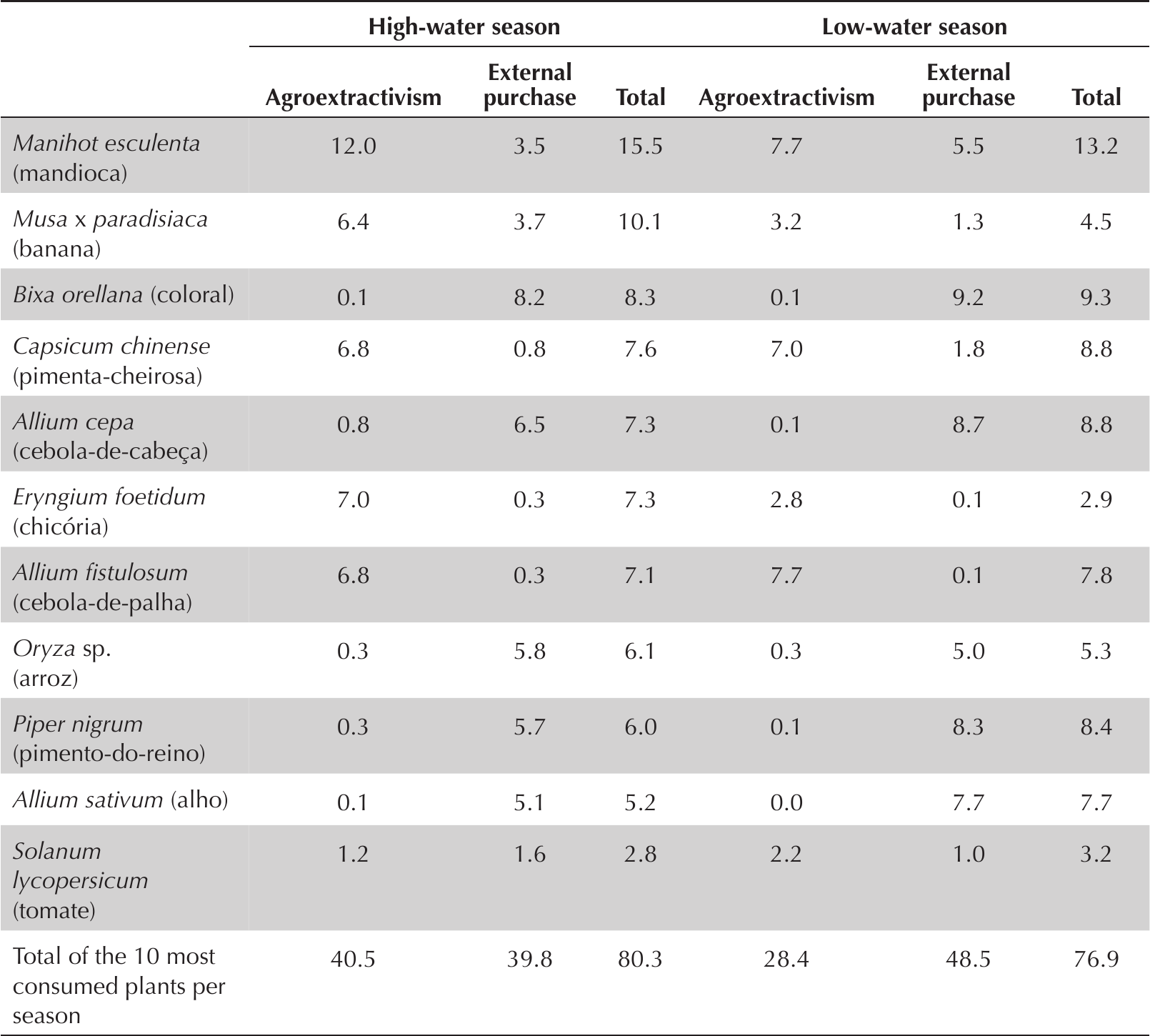
Within the staple plant-related diet, the difference between the high- and low-water season lists of the ten most-consumed plants is minimal. Although the same plants are consumed (e.g., Musa x paradisiaca and Piper nigrum), the frequency of consumption varied between seasons (Table 3). The provenance of such highly consumed items is generally via purchase from sources external to the communities.
Total of consumed plants reported by all interviewees per provenance and flooding season.

Manioc was the most important plant item for the daily diet of the communities. There were notable seasonal differences in provenance: in the high-water season, 77.7% came from local production and 22.3% from external purchasing. In the low-water season, 58.2% of total consumption of manioc came from production and 41.8% from external purchasing. In both cases the consumption of locally produced manioc is higher, but in the dry season, the purchase of manioc is almost double that in the high-water season.
Discussion
The Importance of Food Diversification, Regionalized Food, and the Threats
Low levels of plant matter consumption are historically well recorded for native Amazonian populations and plant diversity is not their main source of energy. The basic diet in the region consists of fish as a protein source and plantain and manioc flour as a starch source (Cascudo 2011; Dufour et al. 2016). Other plants, especially fresh fruits, are consumed sporadically as “secondary foods” (Adams et al. 2005; Alencar et al. 2002; Dufour et al. 2016; Katz et al. 2012; Murrieta 2001; Murrieta and Dufour 2004; Murrieta et al. 2008). Plants are mostly consumed with main meals, although with little variation, as the prevailing use is as a spice. The greatest diversity of plants is usually consumed as snacks, as registered here (Table 1) and in Murrieta and Dufour (2004); however, in our study, this occurred mainly in the high-water season, due to the phenology of fruiting plants and the easier access by boat to flooded forests for plant harvest. This is particularly important because snacks have been significantly replaced by industrialized foods.
For eating habits, besides variation associated with annual seasonality, phenology of the species involved and the culturally and historically constructed eating habits must be considered. This now needs to include a food transition variable, since rural communities in the Amazon have increased access to the ultra-processed products that are likely to eventually replace diet functions previously filled by plants (Cerda 2014; Popkin 1993). While living with regional communities, powdered juices were seen to be more commonly consumed than fruit-derived juices, even though appropriate fruits grew close to the communities. Likewise, as reported by the elders, macaxeira porridge, green banana, or carimã (cassava starch) has been replaced by industrialized corn or rice starches for feeding children. Afternoon snacks, for example, are more easily bought in the form of salty fried snacks than they are collected from the forest. In the community schools, the menu offered by the city hall is based on processed, canned, or frozen items, no product on the menu is purchased from local producers, and the school cooks add spices brought from their own gardens to maintain the flavor of local culture in the predefined school menu.
Industrialized products are not new to these populations, since they have been present in the diet since rubber exploitation began (Loureiro 1981). However, such components are no longer basic items, such as sugar, coffee, and oil, but also secondary items, such as industrialized snack foods (cookies, salted chips), soft drinks, and juice powder, and occupy, in part, the space previously occupied by fruits, especially for snacks between main meals. For example, we can cite items mentioned by older respondents, such as amapá milk (leite do amapá, the latex of Brosimum spp.) previously consumed with coffee but now replaced by powdered milk; annatto powder (Bixa orellana) replaced by industrialized colored flour; cooked breakfast tubers replaced by wheat flour; and sugarcane molasses replaced by refined sugar. Products appearing on the shelves of all the communities are the same and, therefore, the diet cannot be diversified if it depends only on trade. The arrival of ultra-processed sales products and easier access to them through government cash transfer programs favors the process of food transition and homogenization that is associated with lifestyle changes (Byg and Balslev 2001; Reyes-García et al. 2005), impacting rural communities even within Amazonia, one of the most biodiverse places in the world.
The Amazon rainforest and its useful species are the result of a process of continuous domestication by peoples who historically have inhabited, used, and managed them, and the conservation of populations of these species is essential to ensure food security of Amazonian Peoples today (Levis et al. 2017). Changing eating habits so that locally grown crops and forest-derived products are replaced by those from the food industry, albeit only between main meals, can pose a threat to the overall food security of communities, which may end up relying on low-quality food, accessed only via money-based transactions and contributing to the devaluation of regional culture and products. Although the focus of this study was the consumption of plants, an extensive consumption of canned and processed meats (such as corned beef and sausages, respectively), replacing fish or game in the main meals, leads us to believe that the effects discussed here about changing eating habits are present throughout the Riverine diet (Nardoto et al. 2011). Consumption of forest plants by Amazon Riverine Populations has long been important, with estimates indicating that such plants provide approximately 16% of the energy consumed by such cultures, as well as being important sources of micronutrients (Dufour et al. 2016). In consequence, low dietary diversity may contribute to suboptimal values for such micronutrients, such as zinc, iron, calcium, and vitamin C (Dufour et al. 2016; Yuyama et al. 2008), while traditional feeding systems are primarily responsible for nutritional diversity and quality in populations from the Amazon (Roche et al. 2007). Forest plants are important sources of micronutrients, especially beta-carotenes (Mauritia flexuosa) and antioxidants (Euterpe spp.); some are good sources of lipids (Poraqueiba sericea) (Dufour et al. 2016). Likewise, cultivated plants are sources of micronutrients, such as vitamin C (especially Citrus spp., but also Anacardium occidentale, Mangifera indica, Psidium guajava, and Theobroma cacao) and beta-carotenes (Bactris gasipaes), as well as macronutrients, especially carbohydrates (Dioscorea trifida, Zea mays, Musa x paradisiaca) (Dufour et al. 2016).
In this context, considering that the greatest richness and diversity of consumed plant species occurs as snacks, our study highlights the fundamental role that this diet class plays in the nutrition of these rural Amazonian populations and the attention it deserves since this category is one of the most vulnerable to the food transition process. Food education actions, initiatives to regionalize school meals, and debates with the communities play a fundamental role in valuing the local food culture.
Despite the ongoing food transition process, the number of plants consumed by the RDS-PP communities remains relatively high. A survey of regional knowledge of food plants, carried out concurrently and with the same interviewees participating as in the present study, identified 220 species of food plants (Machado and Kinupp 2020). The number of species recorded here resulted from sampling of only one day of consumption for each season and it was possible to recover about 33% of the total known species in just two days of sampling. Thus, it is likely that the true number of species consumed annually is substantially higher than that recorded in the current study, if all times of the year were fully sampled. These results, therefore, indicate a diet that includes its regional resources and knowledge, despite the threats.
Diet Seasonality and the Dietary Dynamic in Different Seasons
If diversity of plant-derived items does not represent the most important part of the diet of Amazonian populations, with plants being consumed sporadically and with little diversification (Katz et al. 2012), it is essential to consider the seasonality with which plant consumption occurs in the region so that possible food-security-related-actions can be planned. Regionally, mature fruits are available mostly from December to March in the terra firme areas, at the rainy season, and in flooded areas from January to August, the peak height of river floodwaters, given the seed dispersal strategy is basically by water or fishes (Haugaasen and Peres 2005). During the interviews, many participants reported that the fruiting season of the most-consumed forest species corresponds to the “nut time,” which extends from January to June, when they work collecting the fruits of the Brazil nut, Bertholletia excelsa, and then sell its seeds. As this tree grows in terra firme, the period also sees an intensification of activities, such as hunting. In this period, the consumption of seeds also occurs in the main meals, especially in the form of broth (nut-milk) that accompanies game meat. The importance of conducting dietary surveys in the rainy season should be noted, since it is likely that the number of records of the consumed terra firme derived plants will be highest and most diverse at this time.
Diet seasonality represents a link between human food and the cycle of forests and natural environments. Eating what the surrounding environment can produce minimizes both the impacts of agriculture on natural resources and the labor required for production, with rationality and seasonality being one of the agroecological precepts of food production (Khatounian 2001). Thus, in an attempt to reconcile biodiversity and food security, the importance of increasing forest species planting and management is emphasized, as this provides a means with which to diversify production and consumption in different seasons (Chappell and LaValle 2011).
Comparing food consumption in cities along the middle Solimões river during high- and low-water periods, Costa (2015) reported that processed foods brought from Manaus (the state capital) or other regions of the country were more frequently consumed during the high-water season and regional and production products consumed more when river waters were low. This seasonality can be explained by the greater availability of fish during low-water, as well as by the possibility of cultivation in the floodplain during this period. In the high-water season, the navigation is facilitated, the consumption of external products increases and food costs are generally higher due to the scarcity of regional products.
From the data collected here, it is possible to add to this discussion that, with regard to plants, consumption tends to be higher during the high-water period, not only because native, flooded forest species are fruiting (Haugaasen and Peres 2005; Shanley and Medina 2005) but because the high waters make it easier to access and collect them (Fernandes 2012). In the dry season, however, when access to flooded areas is more difficult, forest fruit consumption decreases, and the purchase of traded plants increases. In addition to species phenology and access to forests, another influential factor is the planting time in agricultural areas. Because communities do much of their agriculture in seasonally available floodplain areas, planting occurs mainly during the low-water season and harvesting as the waters begin to rise. As a result, during the high-water period, abundance of plants for consumption, including manioc flour, is higher (Murrieta and Dufour 2004).
In this context, it is interesting to note that, based on observations made during field work, the high-water period is considered to be the time of scarcity, when it is difficult to obtain food in general. Even though there is a greater abundance of plants, fish are less abundant than in the low-water season and this leads to discussion of what is considered “food” by communities. According to Murrieta (2001), in Riverine communities, foods, such as spices and vegetables, are relegated to the category of “not food.” Thus “food” comprises the central items of the families' diet: fish and manioc flour. The lack, or even scarcity, of this combination is accompanied by the perception of hunger, even though manioc is sometimes considered “poor food” and does not have the status of rice, which is bought externally.
Speculatively, we suggest that the different consumption patterns presented by each community separately in the Non-Metric Multidimensional Scaling (NMDS) analysis may be related to their productive profiles, the environment in which they are located and the access to urban centers. Uauaçu and Fortaleza communities, for example, have similar patterns (Supplementary Figure 1), and have in common a remote location, with difficult access to urban centers, especially in the low-water season when the small rivers that connect them to the nearest urban center dry up completely. Thus, the supply of these communities becomes restricted to a single supplier boat in the low-water season, visibly differentiating consumption between seasons. Uixi and Itapuru communities are located in terra firme areas, but Uixi presents a more agricultural profile, with greater consumption of food produced within the community and with more extractive activities. Perhaps that influences the strong consumption diversification in the high-water season. Itapuru is nearby an urban center and other communities, for which the access rivers do not dry, maintaining a constant trade between seasons, with a diversification in the high-water season. The Cuiuanã community is located in a várzea area, with no access to nearby terra firme, and it is supplied mainly by the urban center or other communities, perhaps that influences the similar consumption pattern for both seasons.
Food Sovereignty and the Provenance of Consumed Items
The predominance of purchases of externally sourced items among the staple foods is of concern in terms of the food sovereignty of these communities, and underscores the importance of increasing agroecological production for self-consumption as a key factor for both autonomy and food diversification in such communities (Fernandes 2012; Khatounian 2001; Siliprandi 2015). Especially in view of the staple diet, which is maintained in both seasons, and where manioc flour plays a fundamental role, Adams et al. (2005) point to manioc agricultural instability as the most sensitive aspect of food security in Amazonian Riverine floodplain communities. Similarly, Silva et al. (2017) found that várzea-based floodplain communities, subject to the uncertainties of flooding during the agricultural period, are major consumers of industrialized products. Therefore, this fluctuation in manioc production at different phases of the annual hydrological cycle deserves attention and action from public policies aimed at providing both food security and sovereignty in the region.
The need to guarantee food sovereignty of traditional populations proved to be urgent in the scenario of the COVID-19 pandemic, which strongly threatened the most vulnerable populations in the Amazon, such as traditional and Indigenous communities (Ferrante et al. 2020). Dependent on external purchases for food, traditional communities were exposed to the risk of contracting the virus, facing difficulties in establishing social distance, and isolation from cities (Ferrante and Fearnside 2020; PAHO/WHO 2020). Important Indigenous and Riverine leaderships were victims of COVID-19. Intensifying community production and strengthening local trade of cultivated regional products, in a way that they are maintained in food habits, as well as promoting fair trade of non-timber forest products are initiatives capable of safeguarding culture, life, autonomy, and social development of Amazonian Traditional Populations. Although Amazonian rural communities are capable of supplying cities with regional agroecological products, there are conflicts of interest, logistical difficulties, and complex political, economic, historical, and cultural issues that make this scenario almost utopian. This study is limited in its ability to answer so many complex questions regarding Amazonian food, but the joint work of different actors, such as NGOs, public authorities, local leaders, scientists, and organized civilian groups, has been essential to achieve better development conditions for the region.
Conclusion
Valuing regional food is a way to conserve sociobiodiversity, guaranteeing respect for the local food culture and the conservation of food plants populations historically domesticated by native peoples. Intensification of autonomous and culturally acceptable production of the food itself through the planting and management of forest species contributes to food sovereignty, and restricts the need for external purchases. Far from exhausting this very complex subject, we believe that the development of public policies aimed at producing economic incentives to intensify production, the introduction of regionalized school meals, and fair local trade of regional products, associated with traditional knowledge, would help provide food and economic security to these populations, together with Amazonian biodiversity conservation, in a socially and environmentally just manner that is in line with the intertwined ecological history of Amazonia and its peoples.
Footnotes
Acknowledgments
The authors would like to express a genuine thank you to the communities of Piagaçu-Purus Sustainable Development Reserve; without your deep ecological and botanical knowledge and great hospitality and patience with us, there would be no research. This work was made possible through Pró-Amazônia Program (Project 052) by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) funds and through CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) scholarship funds. We acknowledge the Programa de Pós-Graduação em Botânica of Instituto Nacional de Pesquisas da Amazônia (PPG-BOT/INPA) and both herbariums where collected specimens have been deposited: Herbário do Instituto Nacional de Pesquisas da Amazônia (INPA) and Herbário EAFM of Instituto Federal de Educação, Ciência e Tecnologia do Amazonas.
The research project was approved by the Research Ethics Committee of the National Institute of Amazonian Research (CEP/INPA) (Certificate of Presentation for Ethical Appraisal [CAAE] number 68477417.5.0000.0006). A Prior Consent Form and the Free and Informed Consent Form were signed by community leaders and by each interviewee, respectively. Permission for entering and researching in Conservation Units was conceded by Departamento de Mudanças Climáticas e Gestão de Unidades de Conservação (DEMUC), number 80/2016. Register for collecting plant material at Sistema de Autorização e Informação em Biodiversidade (SISBIO), number 55263-2.