
Abstract
Semi-domesticated plants are good resources for agroecology, because they can survive without human intervention and can be managed together with non-domesticated plants. To explore the ecological know-how underlying farmer management of semi-domesticates at the landscape level, we compared how Yanacona Indigenous People in the southwestern Colombian Andes manage two semi-domesticated species of Phaseolus beans, both locally named cacha (Phaseolus coccineus and P. dumosus), and a full domesticate (the common bean Phaseolus vulgaris). We investigated what functional traits of Phaseolus beans farmers identify and how farmers link these traits with the ability of beans to develop in different cultural-ecological niches within the landscape. In 60 semi-structured interviews conducted with Yanacona farmers, we found that (i) farmers compare functional traits of the different bean species, including perenniality, phenology, resistance to pests and diseases, and ability to compete with surrounding vegetation; (ii) farmers recognize and use the variation generated by hybridization between the two cacha species; (iii) farmers take advantage of the traits of cacha to grow them under a wide array of niches at the landscape scale, including low-intervention spaces, such as hedges and swidden plots in fallow, and have detailed knowledge of cacha beans' interactions with trees; and (iv) cacha traits contribute to the resilience of agroecosystems through the management practices and social relationships with which they are associated. Emic and etic perspectives must be combined to gain full understanding of the roles of functional traits of cultivated plants.
Keywords
Introduction
In addition to the pool of species that have become truly dependent on humans for their survival and dispersal, many plant management systems created by Indigenous Peoples and local communities (IPLC) encompass plants that may be termed “semi-domesticates” (Blancas et al. 2010; Iriarte et al. 2020; Neves and Heckenberger 2019). This term applies to any plant population “that is significantly modified by human selection and intervention [and retains] sufficient ecological adaptability to survive in the wild if human intervention ceases” (Clement 1999:190). Semi-domesticated plants are often viewed merely as “future domesticates” or “unfinished domesticates” that compare unfavorably with true domesticates in terms of their performance in field conditions. Interest in them is often limited to how they might contribute to breeding programs for true domesticates. However, traits that are considered sub-optimal in monocultures can prove to be very important in agroecological systems, and semi-domesticates may, in some cases, be seen as examples of balanced partitioning between the supply of benefits to humans and ecological adaptation. Thus, semi-domesticates are a good model for studying how various plant traits and human management can interact with positive effects in agroecological systems.
Although some publications suggest that farmers recognize and value functional traits (Isaac et al. 2018; Martin and Isaac 2015), we currently lack studies designed to understand how local people describe plant traits linked to ecological processes (e.g., competition, facilitation, adaptation to different abiotic conditions). As functional traits, such as perenniality (which characterizes some semi-domesticates), are increasingly recognized as key for agroecology (Barot et al. 2021; Ciotir et al. 2019; Ryan et al. 2018; Soto-Gómez and Pérez-Rodríguez 2022), we argue that knowing how these traits are perceived and mobilized by IPLC can help us understand the coherence of local agroecological systems.
The purpose of this article is to investigate, within a wild/domesticated gradient, what plant functional traits farmers consider to be important and how these traits interact with management practices at the landscape scale. We focus on the management by the Yanacona people in the Colombian Andes of three species of beans (Phaseolus, Fabaceae): P. vulgaris, the common bean, and two species grouped as “cacha” beans, P. dumosus and P. coccineus. For all plants mentioned in this study, full scientific names with the author of the name, common names, and botanical family are given in Table S1. Beans are, along with maize (Zea mays), the main crops in this area. The two closely related species, known collectively as cacha, are interfertile and are within the secondary gene pool of P. vulgaris. All three species are grown within the same landscape in our study site. Phaseolus vulgaris is annual and is, in this area, a full domesticate. In contrast, the other two species are perennials characterized by intense and prolonged vegetative growth. Phaseolus dumosus can be considered to be a semi-domesticate, as it shows signs of domestication (see Methods) while having the ability to form feral or escaped populations in secondary vegetation (Schmit and Debouck 1991). Phaseolus coccineus shows this ability to feralize to a lesser degree. These three species were studied because they are key elements of farmers' diet and together cover a large phenotypic and ecological range. Specifically, we addressed the following questions: (1) How do farmers perceive and describe the interspecific and intraspecific variation in bean functional traits; (2) Are these locally important functional traits associated with different management practices in the different landscape units; and (3) Does the inclusion of the semi-domesticated beans make the food system more stable or resilient than if only P. vulgaris were present?
Methods
Study Area
We report here the results of a field study in Pancitará, an Amerindian resguardo (a sociopolitical entity having its own legal system) of the Yanacona Indigenous People, with a population of about 4000 persons and covering an area of 78 km2, ranging from 2200 to 3500 masl. This resguardo is part of a larger region occupied by the Yanacona in the Colombian Macizo, in the Andean region of the Department of Cauca, in southwestern Colombia (see the broader geographical context in Figure 1). The Yanacona are Spanish speakers, although some toponyms and patronyms and some elements of their vocabulary are related to Quechua and other Indigenous languages (Romoli de Avery 1962). The Yanacona are organized within a resguardo that requires weekly participation in mingas, i.e., collective work sessions or meetings.
The climate of the area is tropical montane, with an ecological gradient driven by temperature between the coffee zone (1000–2000 masl), the intermediate bocage zone, marked by mixed woodlands, pastures, and fields, and the páramo zone (above 3000 masl). Our study focuses on the intermediate zone (2000–3000 masl), where all three bean taxa are cultivated (see a representative photograph of the area in Figure 2). The temperature varies little during the year (mean annual temperature estimated at 13 °C in the center of Pancitará). Variation in rainfall defines four seasons: the main dry season being between June and September and a period of reduction in rainfall during the rainy season happening from December to February. The average annual rainfall is 1257 mm (DHIME 2022, data calculated between 2011 and 2021). The vegetation present in the intermediate zone corresponds to an Andean forest (bosque andino) more or less fragmented by fields and pastures, developing on steep valleys with schist-derived soils.
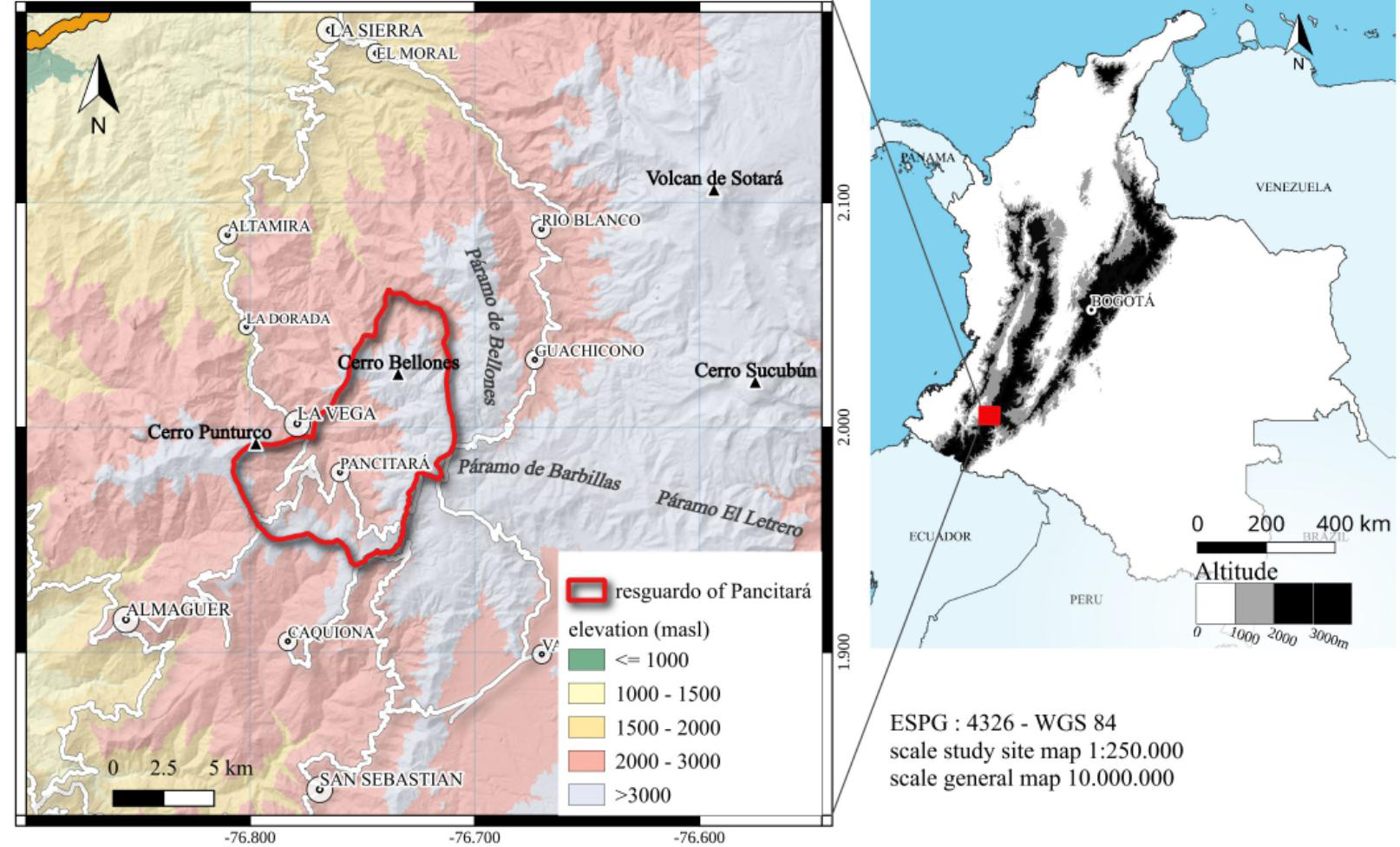
Study site (Yanacona resguardo of Pancitará) and its location in Colombia (1° 58′ 45″ N, 76° 45′ 40″ W).
Farming is non-mechanized and predominantly subsistence-oriented, with small quantities produced for local markets. Wheat was an important crop in the past but experienced a decline in the 1990s, replaced by amapola (poppy, Papaver somniferum), whose cultivation introduced social and agricultural instability for about 15 years. The intense repression by the State and the fall in the price of poppy caused the cessation of cultivation of this cash crop, which led to an extensive rural exodus. The total cultivated surfaces decreased and the agricultural system was reoriented towards food crops. A greater part of the surface was devoted to cattle raising, which today constitutes the most important financial resource. The use of ox-drawn ploughs was common until the 1990s, especially for wheat, but after the period of poppy cultivation, this technique was abandoned in favor of manual hoe tillage and shifting cultivation. As in many Andean regions (Mayer and Fonseca 1978; Murra 1972), people in the Macizo developed a food exchange network between the zona caliente (up to 2000 masl), where coffee (Coffea arabica), sugar cane (Saccharum officinarum), and plantain (Musa × paradisiaca) are grown, the zona fría (above 3000 masl), where potato (Solanum tuberosum) and broad bean (Vicia faba) are the major crops, and the intermediate zone—the focus of this study—where maize and Phaseolus beans are the main crops. Households of this last zone own on average 2 ha of land (ranging from 0.5 ha to 6 ha [resguardo Administration, pers. comm., Pancitará, December 12, 2019]). The products are exchanged between ecological zones through local markets, but many Indigenous families have plots in all three zones, even outside of the resguardo, managed either by the same individuals or by different members of the family.

View of the landscape of the intermediate bocage zone where our study took place. Vereda El Higuerón, Pancitará, March 2, 2020. 1) maize crop in a homegarden; 2) roza (first year swidden); 3) rastrojos (swidden plots in fallow); 4) pasture; 5) hedges; 6) Pancitará resguardo main town.
Within the resguardo, the agricultural landscape is characterized by scattered settlements with a combination of homegardens, pastures with hedgerows, swidden plots, and intermediate spaces described by the term monte. As these spaces are important for understanding how the Yanacona manage cultivated biodiversity, we will briefly describe the characteristics of each.
The homegarden (huerta or huerto; in Indigenous cultures chagra or chakra) is the space closest to the dwelling. It is also the space that is given the most attention in terms of time and labor. Traditional houses are built on a slope shoulder and the homegardens, whose surfaces range from 0.05–0.3 ha, are located in the slope immediately below the house, after a row of flowers and fruit trees. The main species cultivated in Yanacona homegardens are maize, beans, pea (Pisum sativum), broad bean, and vegetables, such as arracacha (Arracacia xanthorrhiza), papa cidra (Sechium edule), and alchucha (Cyclanthera pedata). Southwestern Colombian homegardens also host many wild and cultivated trees that will be described below.
Swidden plots range from 0.1–1 ha and are dispersed within the landscape. They are often located within a radius of a one-hour walk from the house. In July and August, some of the trees and shrubs are cut down, left to dry, and subsequently burnt on the ground. Swidden plots have two different names: in the first year, a plot is fully cultivated and called roza and the subsequent fallow (for 2–10 years) is named rastrojo. The latter may also be converted to pasture.
Pastures are the most common landscape feature, as cattle raising is the main cash source. Tree hedges, which serve to delimit pastures, are a key feature of the Pancitará landscape. By far the most common tree species in hedges is lechero (Euphorbia laurifolia), because it grows easily from cuttings. It is not browsed by cattle because of its toxicity.

Variation in visual aspect of seeds and flowers in common bean and cacha beans. The size bars are for bean seeds. All photographs are from Pancitará in 2019 and 2020. Flowers of P. dumosus are typically white, while those of P. coccineus are red and those of the hydridized forms are light red. Seed color is typically straw to cream in P. dumosus and purple with dark speckles in P. coccineus, while it is cream, light purple or light red with speckles of various colours in hybrid forms. For the cacha beans resulting from introgression, only some examples among the numerous colors are given.
The monte (a general Spanish term encompassing a wide range of unmanaged secondary vegetation) is not a well-delimited category of space. The term is mainly used to denote a non-intensive management of space. It applies to fallows, roadsides, or riversides.
Species Studied
We selected this study area because three species of the genus Phaseolus are present: the common bean and the two bean species locally known as cacha, namely the year-bean (P. dumosus) and the scarlet runner bean (P. coccineus), shown in Figure 3. Five Phaseolus species have been domesticated worldwide (Bitocchi et al. 2017), and two of them, P. lunatus and P. acutifolius, are not present in our study area. Of these five species, the common bean has received by far the most attention from agronomists. For example, although breeding programs have used P. dumosus as a source of resistance genes for the common bean, no program has been dedicated to breeding it directly. However, cacha beans have interesting characteristics that may be useful in small-scale agriculture. Unless stated otherwise, the term “cacha bean” applies to both P. dumosus and P. coccineus, following Yanacona naming.
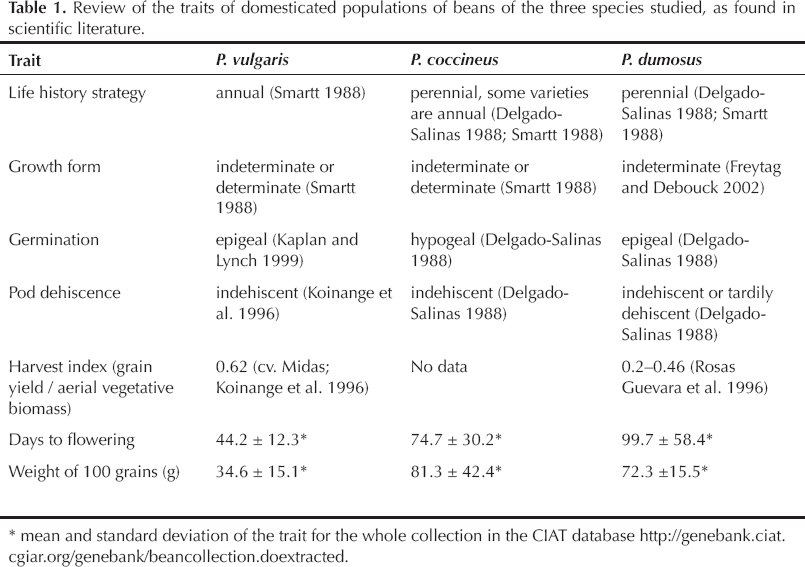
Table 1 compares some of the principal traits that differ among the three species studied here. The common bean is an annual, whereas both cacha beans are perennial, although some annual culti-vars of P. coccineus exist (Delgado-Salinas 1988). Among the five Phaseolus cultigens, P. dumosus is considered to have “a higher unrealized evolutionary potential” than the other four (Schmit and Debouck 1991:359). Its domestication affected seed size and, possibly, seed dormancy and pod dehiscence, but cultivated populations retained the same ecological habit as the wild populations of the rainforests of Guatemala: a vigorous climbing habit and a perennial life history (Schmit and Debouck 1991). More precisely, the roots and the base of the stem (up to 1–2 m) are perennial. These characteristics give it a great capacity to maintain itself in clearings and even to become feral in the secondary vegetation of neotropical mountains, from northern Puebla (Mexico) to Ayacucho (Peru) (Schmit and Debouck 1991). Phaseolus coccineus is characterized by its hypogeal germination and its fleshy root, able to resprout every growing season (stems are also able to resprout, like those of P. dumosus). Both P. dumosus and P. coccineus are predominantly allogamous, whereas P. vulgaris is predominantly autogamous. Both cacha species were domesticated in Mesoamerica but their migration towards the northern Andes is poorly documented. Moreover, taxonomic uncertainty has long placed P. dumosus as a subspecies of P. coccineus; therefore, the former species is absent from archaeological records (e.g., Kaplan and Lynch 1999), making its history difficult to trace. Phaseolus dumosus may have spread southwards in the Andes in the late pre-Columbian period (Schmit and Debouck 1991). In southwestern Colombia, given the ubiquity of this species in mountain agroecosystems and its importance as a marker of Indigenous cultures (Sanabria and Argueta 2015), it has probably been an important species for several centuries. Phaseolus coccineus may have arrived later in the region, the main argument for this contention being that, although the two species can cross-breed easily, hybrid populations are not widespread. In Pancitará, P. coccineus was unknown to farmers until the late 1970s.
The more or less recent admixture of both cacha species in the same environments in southwestern Colombia generated variability in the population of cacha by introgression, as was noticed by Schmit and Debouck (1991) and described by Molano (2006). Whether these hybrids exhibit different ecological characteristics has not yet been investigated. Some studies mention cacha bean (both species) as a sometimes cultivated, sometimes subspontaneous component of the agroecosystems of Latin America, from Mexico to Peru (Basurto Peña 2000:42–45; Debouck and Schmit 1989; Debouck et al. 1990; Sanabria et al. 2005), but little research has been conducted on their uses and the agricultural practices associated with them. Cacha beans are, however, good examples of resource plants for agroecology, their perennial character allowing the generation of an extensive root network that contributes to soil conservation and maintenance of soil biota. Furthermore, their cultivation is compatible with the inclusion of other perennial plants in the agroecosystem, because their vigorous lianescent character allows them to climb on trees.
Study Methods and Ethics
We used an ethnographic approach to document the knowledge and practices of local farmers from an emic perspective (i.e., the farmers' point of view). The first author undertook participant observation over four months (November 2019–March 2020). During this time, the first author conducted 60 semi-directive individual interviews of 1–2 hours each, with 24 female and 36 male respondents, their ages ranging from 31 to 87 (Table S2). These interviews focused on agriculture, botany, food, landscape, and the management of different bean species. They were done at the farm and complemented with visits to the cultivation plots, 20 full days of agricultural work, 15 commented walks with respondents on trail paths, and everyday discussions in the village center, where the first author lived for 110 days. The study was made possible by authorizations from the resguardo's cabildo (council) after community approval during a meeting organized on December 8, 2019, to present the objectives of the study. Interviews were recorded, transcribed, and anonymized by replacing their name by another local name if they did not wish to have their name cited.
Differences Perceived by Yanaconas among Phaseolus Types
In the recent past, Yanaconas named cacha bean “torto” or “plancho,” and sometimes “pipejegua” (literally “mare's belly,” referring to the shape of the seed). This bean, previously disparaged by the non-Indigenous population, came to be valued by lowlanders as a “healthy food” at the same time as Indigenous cultures began to be recognized at the national level. As a result, the word “cacha,” the term used by lowlanders as well as by other Amerindians in southwestern Colombia, has been used by the Yanacona since the 1990s. For convenience, we use “cacha” here.
When asked what the main differences between the common bean (“frijol de año” or “frijol varero,” literally “year bean”—for its annual habit—and “pole bean,” respectively) and the cacha beans are, the Yanacona predominantly mention five traits of ecological importance (nearly always cited by respondents). The first trait mentioned is the lifespan. The common bean is an annual and has to be sown every September. In contrast, the cacha beans are short-lived perennials that resprout at the same period of the year. They are considered to be short-lived in comparison to shrubs and trees. However, in contrast to previous estimations of P. dumosus lifespan ranging from three to six years in agronomic trials (D. Debouck, pers. comm., December 6, 2019), Yanaconas mention a much longer lifespan for this species, which they say can reach more than 30 years. Personal measurements made by the first author show that stem diameter at the base can reach 3 cm for the oldest individuals mentioned, while for individuals indicated by farmers to be around five years old, it never reached 2 cm. Few local observations on lifespan were collected for P. coccineus, but personal observations suggest it is comparable to that of P. dumosus.
The second difference recognized by Yanaconas is phenological (Figure 4). At this elevation, pods of common bean can be harvested five to eight months after sowing, depending on the variety. In contrast, fruits of cacha bean are mature eight to 11 months after sowing, depending on the species (P. coccineus being slightly earlier than P. dumosus), but also on the microenvironment. However, when the plant resprouts in the second year, maturity is attained much faster, in about four to five months, and, if the weather is favorable (alternance of sunny and rainy days), plants can subsequently produce pods every three months. The ability of cacha to yield at various seasons, especially in the second and subsequent years, is evoked in another nickname of the cacha bean (referring mostly to P. dumosus) among the Yanacona, the “frijol de todo el tiempo” (i.e., “all-time bean”). We can relate it to the name “todo-el-año” (“all-year”) given to P. dumosus by farmers of the Altiplano Cundiboyacense, Colombia (J. Locqueville, pers. obs., May 2019). Moreover, cacha beans are usually harvested as green pods but can also be harvested when dry. As dry pods are tardily dehiscent (strongly indehiscent in humid conditions but dehiscent under hot and dry conditions) and do not abscise, they can still be harvested several months after maturity. Combined with the fact that the phenology of cacha varies slightly with elevation, it is possible to harvest cacha pods at virtually all times of the year. This characteristic has an important consequence; while the common bean is harvested as dry pods once a year, thus, must be conserved in the granary during a whole year, with a risk of attack by bruchids (gorgojo, Chrysomelidae), the cacha bean pods are generally harvested every day in small amounts. This also makes the cacha bean a useful food for periods of scarcity. “Before, we always said April was the month of hunger, but I didn't understand, as there were plenty of cacha and arracacha” (Siempre deciamos antes que Abril era el mes del hambre, pero yo no entendía, pues había mucho fríjol cacha y arracacha [Nidia, July 3, 2020]).
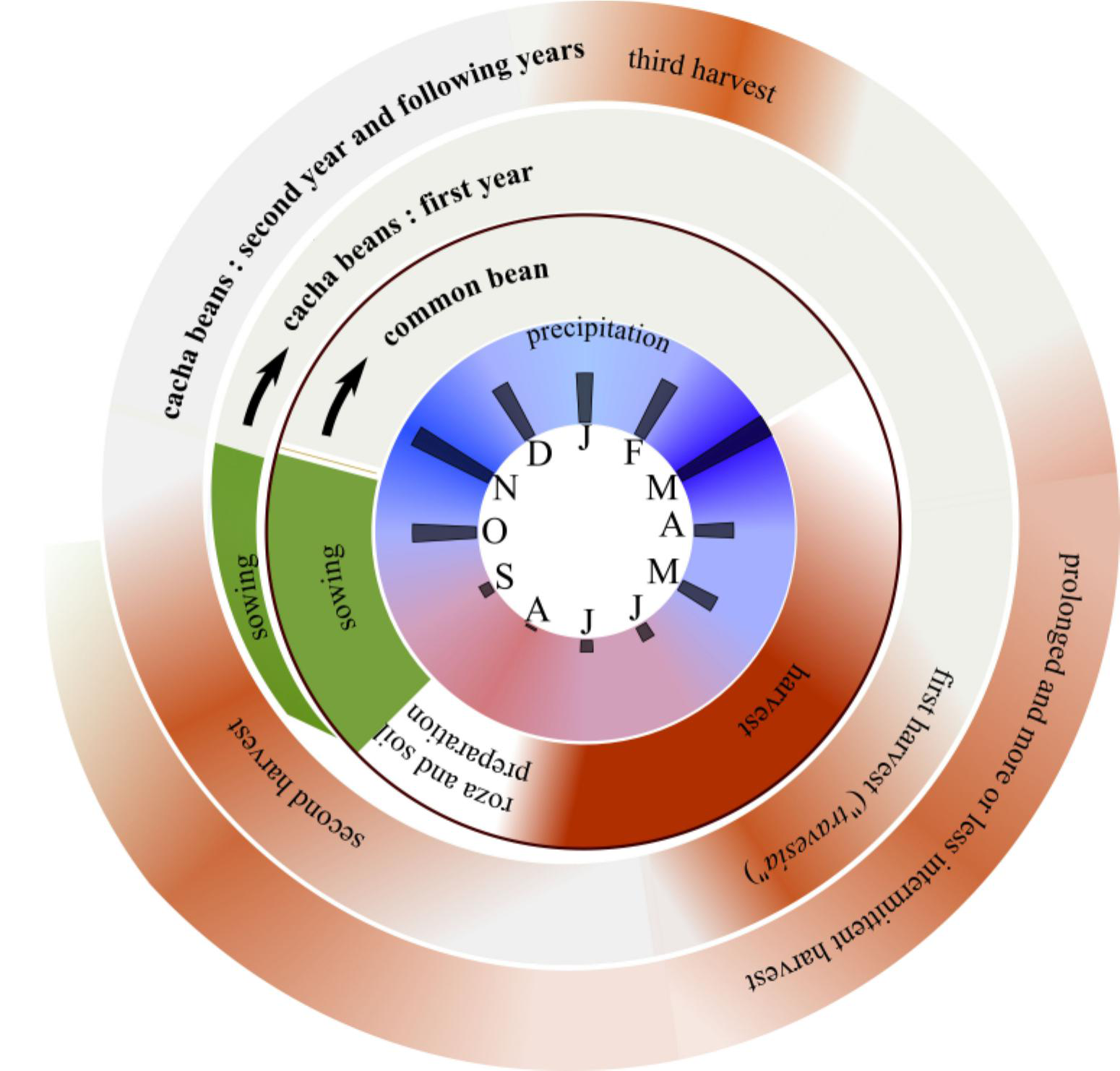
Local calendar for common bean and cacha beans. Monthly precipitation is given at the center, with a maximum at 300 mm/month. Second from the center is the yearly cycle for the sowing and harvesting of common bean. Third is the calendar for the two first years of the cacha bean cycle. From the third year onwards, the cycle is similar to the second year. Intensity of the color is an indicator of the availability of the resource at a given time. However, for cacha beans, the annual distribution may vary strongly from one year to the next.
The third trait often mentioned by farmers is the level of attack by bruchids. When seeds are stored in granaries, the cacha beans are said to be much less attacked than the common bean. However, farmers state that young pods of all three species are occasionally oviposited upon by insects. The fourth difference (possibly related to the third) is in the plants' resistance to other pests and diseases, especially in humid conditions. The common bean is seen by Yanacona as more prone than cacha bean to rain-driven fungal infections. “The rainy season spoils [the common bean], the leaves get rotten” (el invierno lo perjudica, lo daña, se hiela la hoja [Simeon, November 29, 2019]) but “it may be rainy, or summer, it doesn't affect much [the cacha bean]” (Puede llover, puede haber verano, la estación climática, los climas poco lo afectan [Juliana, November 29, 2019]). This is in concordance with the fact that both species of cacha are mentioned by breeders as a useful source of resistance against anthracnose and Ascochyta blight (Porch et al. 2013).
The fifth trait farmers mention, perhaps the most important in the context of our study, is the capacity of cacha to live and develop without human intervention, thanks to its highly competitive climbing behavior. “It has the advantage that it is not necessary to cultivate it” (tiene la ventaja que no hay necesidad de cultivarlo [Nestor, December 9, 2019]). A single individual is able to explore the nearby vegetation, as well as to produce a thick “mattress” (colchón) on the tops of small trees and shrubs. This trait gives the cacha bean a great capacity to maintain itself in secondary vegetation without human intervention.
Two other characteristics, less frequently spontaneously mentioned (probably because of the nature of the question), mitigate the ecological advantage of cacha and explain why the common bean remains an essential element of the system. The first is ease of harvest. The cacha produces pods that are sometimes difficult to reach, at several meters high, whereas the common bean is much easier and quicker to harvest. The second is culinary. Cacha, especially when beans are mature and dry, is sometimes perceived as more difficult to digest and is less appreciated by the younger generation. Some people prefer to throw away the first cooking water after a short boil. The two classes of bean are often perceived as complementary for cooking: “The common bean is better for rice, frijolada. Cacha is better for corn soup. It gives a flavor like meat, either chicken or beef” (El frijol de año es más para el arroz, la frijolada. El cacha es mejor para la sopa de maiz. Da un sabor como a gallina, a carne, sea de gallina o de res. [Eudilia, December 3, 2019]).
Differences among Intraspecific and Hybridization-Driven Variation
In this community, several landraces and commercial varieties of common bean are cultivated, often in mixture. Both climbing (indeterminate growth) and bush (determinate growth) varieties of common bean are present, the latter being cultivated only in small amounts and only since relatively recently (probably around the first decade of the 2000s). Yanaconas mention few variations in traits of the common bean.
As both cacha species are cross-compatible and predominantly allogamous, the introduction of P. coccineus has led to creation of a large pool of hybridization-driven variation (Figure 3). Farmers are aware of this mechanism of formation (“se casan,” literally, “they get married”). Of the different cacha bean types they distinguish, two are known as the “proper cacha” and are typical P. dumosus, and only distinguished by their seed color. They both have a wide hilum, white flowers, and epigeal germination, and differ by their seed coat color (cream- and straw-colored). A third variety is close to true P. coccineus, with scarlet flowers, hypogeal germination, white or violet seeds, sometimes with black speckles, and a narrow hilum. The other varieties are less well-defined and are named according to the seed color. They exhibit attributes of the two species (e.g., both hypogeal and epigeal germination can be observed within the same seed color) or intermediate characters (e.g., pale red flowers).
Although this P. coccineus-driven variation (resulting from hybridization or not) has been present in Pancitará for less than 40 years, Yanaconas have experimented with it and mention two functional traits in which the different cacha types differ. First, P. coccineus and several hybrid types are known by farmers to resprout more easily after burning. This trait is likely associated with the fleshy, tuberous nature of P. coccineus roots and with its hypogeal germination (Delgado-Salinas 1988). When germination is hypogeal, there are meristems both on the epicotyl (partly aboveground) and at the cotyledonar node (belowground), thus, in case of damage to the tip of the epicotyl, a second growth is still possible (fueled by the reserves of the cotyledons), and this capacity is maintained at later stages. This trait has been suggested to increase tolerance to several kinds of disturbance (fire, herbivory) in wild relatives of manioc, while manioc itself is characterized by epigeal germination, perhaps an adaptation to cultivated environments (Pujol et al. 2005). In contrast, Phaseolus dumosus-like types have epigeal germination (Delgado-Salinas 1988). Thus, if the aboveground meristem is damaged, the seedling is not able to resprout and, at later stages, its fibrous roots, because of the absence of meristems, may not as easily resprout after a burn or other disturbance. The second functional trait mentioned by farmers is the fact that P. coccineus or cacha types showing introgression from P. coccineus are less competitive in secondary vegetation and disappear more quickly, suggesting a lower rusticity of these bean types in this environment.

The three species of bean cultivated in Pancitará and the range of spaces where they grow in the landscape. The line is dotted when the situation is seen occasionally.
Local Management Practices of Beans
Because they drive particular plant-plant interactions, functional differences between bean types may have direct consequences on how they can be cultivated and on the types of spaces in which they can be managed. Therefore, practices of the Yanaconas, with respect to beans, must be understood at the landscape level and within the complexity of interspecific relationships (Figure 5).
In Pancitará, the common bean is always sown in c5ltivated plots, whether in homegardens or in rozas. One of its nicknames is frijol del maíz (“maize bean”), as it is without exception sown along with maize. Three maize seeds with one or two bean seeds are sown within a hole made with a stick (barretón). This practice, an essential part of Mesoamerican milpa, is a good example of a practice favoring a mutualistic relationship between the two plants (Lopez-Ridaura et al. 2021). But the positive effects of intercropping depend on the relative growth of the maize and bean when grown together and are, therefore, highly dependent on the varieties or landraces used. A common problem arising when both species are sown at the same time is the faster development of beans, which shade maize and decrease its production. In Pancitará, common bean can be sown with the local, tall-growing (but late-maturing) maize landraces. These maize landraces (capio, labura, blanquillo, amarillo, susunguero) were formerly the only ones cultivated. Although early-maturing commercial varieties are now common, they have not completely replaced landraces, which are considered to be more resistant to the pathogens that are an important problem in humid conditions. Cacha beans, in contrast, are rarely sown along with maize, as they are too competitive for light capture. Debouck (1994) reports that, in Guatemala and Costa Rica, the adoption of early-maturing varieties of maize with soft stems (replacing late-maturing landraces grown previously) led to the abandonment of cacha bean (in this case P. coccineus).
Review of the traits of domesticated populations of beans of the three species studied, as found in scientific literature.
The rusticity of the cacha bean allowed Yanacona farmers to develop a different agroecological system based on positive interactions of this bean with trees. There are three main management regimes of the cacha beans, each located in different places in the landscape: swidden-fallow systems, homegardens, and hedgerows. We use the term “regime” rather than “system” to emphasize the idea that it is not a fully designed and controlled assemblage. Each management regime has a different set of tree species that influence the management practices applied to the cacha. A special case is the absence of management, which will be discussed later.
In swidden plots, the cacha bean is sown at the same time as maize and common bean in the same plot, at the foot of tree poles (frijoleros) that were conserved during the slashing of vegetation. Selected trees are cut with a machete at 2–3 m high, and branches are pruned to convert the trees into poles that will support the vigorous cacha vine. Trees usually survive this treatment and resprout quickly. This system has been reported in other parts of Colombia (D. Debouck, pers. comm., December 6, 2019), as well as in the highlands of Mexico (Basurto Peña 2000:66), but its ecological implications have not been discussed extensively. In Pancitará, when cacha bean is sown, the density of poles can reach 1000 per ha. No special protection is put in place to protect trees from fire, so, depending on fire intensity, cut trees will die or be able to produce new branches during the following season. During the first year, weeding is done with machetes on average three days per ha per month. By comparison, the time dedicated to the weeding of maize and common bean in homegardens can reach 15 days per ha per month during the beginning of the growing season. This difference of time investment is partly driven by the fact that weed abundance is decreased by adequate fallow periods in rozas compared to homegardens. In a roza, tree branches may be pruned if they limit cacha growth.
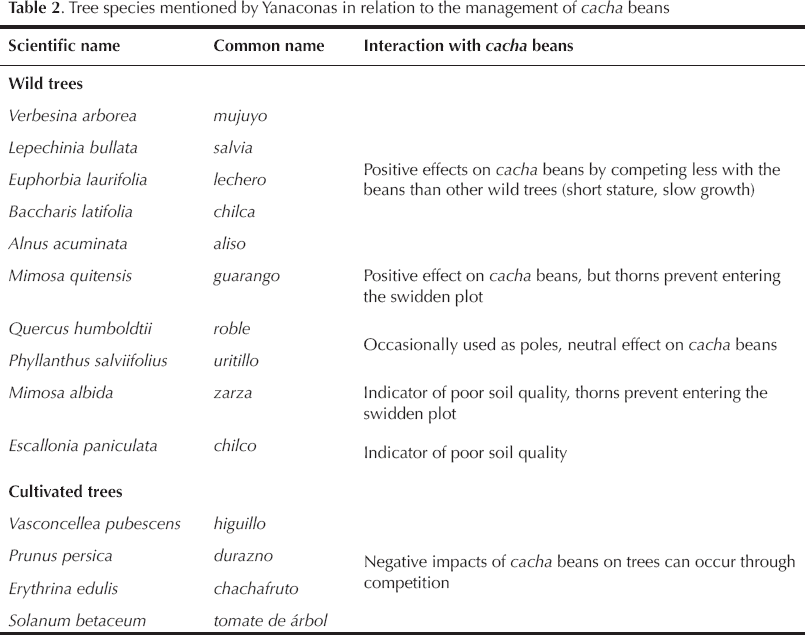
An important feature of this management regime is the omnipresence of wild trees. Thus, Yanacona have developed a knowledge of tree-cacha interactions, summarized in Table 2. Their knowledge leads to: (1) a selection of swidden plots (for example, if Mimosa spp. bushes are present, farmers will prefer to choose another plot, if one is available, because of these trees' thorns); (2) a selective conservation of tree poles to support cacha plants, as trees are chosen that are likely to sprout again; and (3) a selection of tree seedlings when weeding to favor the most desirable tree species. For example, aliso (Alnus acuminata) is often left alive in the swidden plot in anticipation of the plot's conversion to pasture, as this tree is known to fertilize the soil and “maintain it fresh” (mantiene fresca la tierra [Joel, January 1, 2020]).
After the roza stage of the swidden plot (in which maize and common bean are harvested), for two years, the fallow is named frijolar (literally, “bean grove”) as cacha becomes the dominant species. No intervention is carried out and cacha is harvested regularly, thus extending the productivity of the swidden plot to several successive years. Over time, trees progressively outgrow the cacha plants, which become less productive. To maintain cacha for a longer time, some farmers coppice the trees (and harvest the beans climbing on the trees at the same time), reducing the shade. Because swidden agriculture in the Colombian Andes is practiced on steep slopes that are prone to erosion, it is often criticized by environmentalists, as evidenced by the “land use vocation map” edited by the Instituto Geográfico Agustín Codazzi (IGAC 2019). We calculated that this map assesses only 12% of the Pancitará resguardo area as being suitable for agriculture, the rest of it being mentioned as being suited only for “forest and [watershed] protection.” However, thanks to the rusticity of the cacha bean, Yanacona fallows remain productive for at least 3–4 years without intervention. This bean also contributes to soil regeneration through nitrogen fixation and litter production, and through the stabilization of soil against erosion by its perennial root system and by those of the numerous trees associated with it and on which its cultivation depends.
Tree species mentioned by Yanaconas in relation to the management of cacha beans
In homegardens, the management of cacha beans differs in two main ways from its management in swidden plots. First, the cacha is often fertilized with chicken and cuy (guinea pig, Cavia porcellus) manure and weeded regularly (relatively labor-intensive hoe tillage), resulting in more regular yields, especially during the dry season, when the pods of cacha beans from low-altitude zones are less abundant. Cacha plants in homegardens are also pruned at 30 cm height at the end of the dry season, as farmers think that this prevents them from exhausting their resources, thereby increasing the plant's lifespan. Second, in homegardens, farmers sow the cacha bean at the bases of both wild and cultivated trees. Wild trees, such as mujuyo (Verbesina arborea), uritillo (Phyllanthus salviifolius), and roble (Quercus humboldtii), are conserved as supports for cacha and are pollarded for firewood. But farmers also plant trees that produce edible fruits (all cultivated trees in Table 2) and they worry about the negative impact on these trees in competition with beans. “If the cacha grows a lot, I cut it down, otherwise it kills the other plants with its shadow. The trees also, it covers them and kills them” (Si hay mucho la pelamos, sino me mata las maticas, por la sombra. Los arbolitos también, los cubra y los mata [Luis, December 12, 2019]). Therefore, there is a high level of direct intervention on cacha plants (trimming plants and occasionally pulling them out). Such examples show that trees are not seen merely as a support for beans and that many plant-plant interactions are subject to day-to-day regulation by farmers.
Hedgerows are a key element of the Yanacona landscape, preventing cattle from escaping pastures and entering fields, thereby protecting crops. Nearly all tracks, pastures, and homegardens in Pancitará are bordered by hedgerows. We estimated their length to be 172 ± 32 m per ha of land (see Methods) in the 2000–2800 masl zone. They are also children's preferred space for collecting wild fruits, such as those of mora (Rubus urticifolius), chimbalo (Solanum caripense), and various species of Passiflora (P. mollissima, P. x mixta, P. ligularis), and for collecting cacha beans. “When I was a child, my father told me: ‘go to harvest some beans in the hedgerows.’ I climbed in the trees and harvested a small bag of them” (Cuando yo era muchacho mi papá me decía ‘va a coger frijol en las ciercas’, subía a los palos y cosechaba una bolsita [Rolando, December 30, 2019]). These hedgerows are an ideal habitat for the cacha bean because they are pruned regularly. Cacha seeds are simply scattered (thrown without being buried) at the foot of trees (mainly Euphorbia laurifolia and Phyllanthus salviifolius) when people pass by, and cacha plants in hedgerows are not given special attention. Despite this lack of attention, owing to the ubiquity of cacha along frequently traveled trails, hedgerows contribute a large proportion to the daily supply of cacha bean. In the recent past (until the 1990s), most Yanacona sowed cacha in hedgerows, but this practice is gradually decreasing in frequency. One explanation for this tendency is the transition from sheep to cow raising. Cows more readily consume the cacha plant in the hedges, greatly decreasing its abundance, especially as stocking intensity has increased.
How Do Cacha Beans Confer Stability and Resilience to the Social-Ecological System?
Cacha functional traits, associated with the diversity of management regimes we describe, confer stability to the food system in various ways. In this part, we will discuss five main ways, involving both ecological and social processes, by which this stability is conferred.
First, even without external chemical inputs, cacha resistance to diseases and humid conditions is such that, according to the Yanacona, there is no year without cacha, while many times, the common bean will give no harvest if no fungicide is applied. “I don't remember a year without cacha, whereas sometimes the [common bean] yield nothing” (El cacha, no recuerdo un año que él no haya granado, mientras que el otro a veces no grana [Dolores, November 26, 2019]). Second, the flowering of cacha bean spread across the year allows a continuous supply of pods and decreases the risks associated with storing large amounts of grain (in particular, destruction of the stock by bruchids). Third, for the same reason, seeds are available at any time of the year. Because dry pods show little dehiscence, Yanaconas can simply collect dry pods on the side of a track if they need cacha seeds to sow in their garden or swidden plot, rather than keep seeds from one year to another. “If I don't have it in the garden, I'm going to take a look over there, maybe there is some on the side of the track, I find one or two pods, well, I take them home and I sow them right away” (si yo no tengo aquí en mi casa, voy por allá, de pronto está en los caminos, me encuentro unas vainitas, pues yo me las cosecho, yo me las llevo para mi casa y las siembro [Monica, March 11, 2020]). This has an important consequence: it generates a constant seed flow between cultivated and unmanaged spaces, thus maintaining strong natural selection for competitive ability and for resistance to pests and diseases. Conversely, seeds from managed cacha plants are often dispersed in hedgerows and rastrojos and easily grow and reproduce in the secondary vegetation.
A fourth way is that its semi-domesticated habit gives cacha more resilience to all kinds of social and ecological perturbations. For example, in the 1990s in Pancitará, many local crops were abandoned to cultivate poppy, which gave Yanacona the financial resources to buy food from stores. This lasted until the 2000s, when the Colombian army began aerial fumigation of glyphosate to destroy the illegal poppy fields (Tokatlian 2003), destroying other crops as well. According to the Yanacona, several local landraces of common bean were lost during this period. The cacha bean, by contrast, never disappeared, as it was scattered in the landscape, including non-cultivated areas preserved from glyphosate treatments. Even though its cultivation was nearly abandoned, it maintained itself in the secondary vegetation and was an important food crop during the crisis at the end of poppy cultivation.
Fifth, the varying management intensities devoted to cacha also give more flexibility to access rights, thus stabilizing food supply through social, and not only ecological, mechanisms. While the common bean is unequivocally a domesticated plant under individual property in the Yanacona agroecosystem, ownership status of the cacha bean is much less clear. We have seen that cacha can be managed in a more or less intensive way, ranging from simple “assisted dispersal” to cultivation with tillage and organic inputs. These different management intensities determine various access rights. Nearly all land in Pancitará is under a regime of private property and all Yanacona agree that, in theory, one could not harvest cacha beans outside of one's own land in Pancitará, except along some riversides. But in practice, the type and intensity of management actions determine the degree of tolerance there is for harvesting of cacha on private land. Soil tillage, swidden plot preparation, and subsequent weeding are time-consuming actions that determine a private access to beans in rozas and homegardens. Sowing is an important management action and usually implies private access, but in some spaces, such as hedges, sowing is often done by scattering cacha seeds collected along the pathways, so it does not represent a heavy investment. In this case, there is a certain tolerance of landowners for harvesting by others. Moreover, these access rights can change with time for the same plants, especially in the fallows where the access rights gradually change as the intensity of fallows management decreases. It is customary for a landowner with a large cacha fallow to allow a neighbor who requests it to harvest pods for personal consumption or, for example, to feed farm workers during a working day nearby. “In a way, if you have a lot in the rastrojos, you are somewhat obliged to give some away, to allow whoever asks to pick up some” (De cierta manera, si usted de pronto tiene harto en algun rastrojo por allí y no lo cosecha todo, si alguién le pide el permiso está un poco obligado a dejar que coseche un poquito [Rodrigo, March 1, 2020]). This “courtesy” around cacha is an emergent social possibility derived from the ability of cacha to sustain itself without intervention. As it produces pods regularly over the year, the cacha bean encourages a more collective use of this resource, especially in spaces remote from the dwellings. In addition to this, some Yanacona now work and live in Popayán, the nearest large city, while still owning land in Pancitará. There is also an implicit tolerance for collecting unmanaged cacha in their land, as it would otherwise not be harvested.
Discussion
A structuring idea in agroecology is that increasing biological diversity (species diversity, phenotypic trait variation, or both) leads to greater stability of the agroecosystem (Isbell et al. 2017; Jackson et al. 2010; Tilman 1999). However, it is less well recognized that a single species can, through the diversity of management regimes it allows, also confer a form of stability. In our study case, the common bean has functional traits that allow only a reduced set of mutually beneficial interspecific plant-plant interactions (with maize only) and, thus, can only be seen in homegardens and roza plots, while the cacha bean, with its set of traits associated with a semi-domesticated habit, allows a wide variety of management regimes and places. This invites us to consider the stabilizing role not only of biological diversity (specific or intraspecific), but also of the diversity of human-plant and human-landscape-plant couplings.
The various ways in which cacha is managed, particularly in hedgerows and fallows, perfectly illustrate the concept of an agronomy of “piloting,” as theorized by Larrère and Larrère (2018), i.e., management viewed as an inflection of natural processes rather than the establishment of an artificial environment. It demonstrates the need to rely on local ecological knowledge to create and maintain agroecological systems. Indeed, the benefits of cacha do not stem from the cacha plant itself but emerge from the combination of environmental management practices, social values, and interactions of cacha with other living beings, such as trees.
The generalized bidirectional flow of cacha seeds between gardens and the wild has also been noted in other Indigenous communities of the southwestern Colombian Andes (Sanabria and Argueta 2015) and contrasts with many systems in which farmers strongly select against such gene flow, e.g., in Sorghum bicolor (Barnaud et al. 2009) and common bean (Papa and Gepts 2003). An issue to be further investigated is whether the current management practices associated with the cacha bean are domesticatory—i.e., exert selection towards a greater dependence on human actions. As Beebe et al. (1997) noted in the case of weedy populations of common bean, the preservation of a tamed but rustic habitat is a key factor that allows the long-term preservation of a semi-domesticated population. This is the case in Pancitará, where cacha bean is either feral or under low-intensity management in a high proportion of spaces. A common hypothesis in the domestication of herbaceous perennial grains is that frequent habitat disturbance (tilling) likely leads to selection favoring annual lifeforms (Van Tassel et al. 2010), because of the tradeoff between investment in vegetative and reproductive organs in the first year. In Pancitará, cacha bean is managed intensively in some spaces, but as farmers do not segregate cacha seeds based on their wild vs. cultivated status, nor on the type of space where they dwell, a rapid domestication process is unlikely. Moreover, because both cacha species are predominantly allogamous, pollen-mediated gene flow is likely, making attempts to closely manage reproduction of cacha in this environment much more difficult.
However, it is difficult to predict how gene flow between P. dumosus and P. coccineus could affect traits in the population. Such gene flow probably does not imply a risk of extinction by hybridization, as it has been described for wild-weed-crop complexes within a species (Ellstrand et al. 1999). But as Yanacona consider P. coccineus as being less able to maintain itself in secondary vegetation, outcrossing could drive the population towards greater dependence on human action. However, outcrossing could also bring more diversity; for example, in the root or seedling traits evoked above. The evolutionary trajectories depend on both ecological and social factors; thus, these must be studied together.
Actions may be taken to promote a wider use of cacha beans, such as integrating them in breeding programs. However, integrating perennial semi-domesticated plants within agroforestry systems is needed (Barot et al. 2021), and species like cacha should be regarded not only as a donor of resistance genes for the common bean, but also for its useful combination of traits in complex environments. Cultivated as a monoculture and as an annual for commercial gain, cacha bean would have few advantages over the common bean. Rather, a breeding scheme is called for that takes into account not only the performance of beans in experimental stations but also in interaction with the practices of farmers and the environments in which they cultivate their plants.
Our results suggest that the common bean and cacha beans are complementary in terms of food provisioning. Cacha bean, however, produces relationships of a special kind, as it is in some situations considered as something between a private and a common good. This study also allowed us to see how thin the line is between “wild” and “cultivated” spaces and plants. Some individuals of cacha may, at one point in their lives, be considered cultivated, and then wild. It is interesting to note that rather than being linked to the species as such, cultural norms and access rights may be linked to the diversity of cultural-ecological niches in which the plant's traits allow it to be grown. Such interactions between functional traits of plant species and the diversity of their roles in agroecosystems are still to be explored in ethnoecology.
Footnotes
Acknowledgments
We gratefully acknowledge the French Ministère de l'Enseignement Supérieur et de la Recherche for a PhD grant to the first author, and the University of Montpellier Soutien à la Recherche 2020 for funding part of the research presented here. We are above all indebted to the Yanacona resguardo of Pancitará and the people of La Vega for their precious contributions and help, and for granting us with research authorizations. We also thank Diego Macias for plant species determinations and other faculty members of the University of Cauca for providing us with much relevant scientific information. Finally, we thank the three anonymous reviewers for their many suggestions for improving this paper.