
Abstract
Forty plant species and groups of related species were identified as potential food sources in the alpine tundra and subalpine forest of the Colorado Front Range. Emergence, flowering, and ripening schedules were monitored from 1994 to 2004, a period that included years with marked differences in precipitation, growing-season temperature, and snowbankmeltout dates. Late-lying snow is a critical limiting factor in the high-mountain environment, delaying the emergence of edible greens and the blooming of root-food species by as much as 2½ to 3 months in extreme years. Fruit and seed production are unreliable at high altitude because favorable moisture and temperature conditions rarely exist at all stages in the developmental process. Ethnographic data from arctic and subpolar regions with ET (effective temperature) values similar to those of the study area suggest that plant foods comprised less than 20% of the high-altitude summer diets of Front Range hunter-gatherers. Percentages would have been lower during cold, snowy intervals such as the Little Ice Age. Visitors to the alpine region, however, spent much of the year in warmer environments. On an annual basis they were not plant-food deprived.
Introduction
The Colorado Front Range is a north-south trending massif at the eastern margin of the Southern Rocky Mountains, in the interior of the North American continent. Centered on the 40th parallel of latitude, the range includes more than 50 named summits higher than 4000 m, many of them located along the Continental Divide in Rocky Mountain National Park and the Indian Peaks and James Peak Wilderness Areas.
Traditionally regarded by Plains and Colorado Plateau archaeologists as “peripheral,” the Rocky Mountain crest and foothills were anything but peripheral to the people who incorporated them into their seasonal rounds (Fowler and Fowler 1971; Goss 1972; Petersen 1977). Evidence for Clovis and Folsom occupation remains sparse (Benedict 1992a; Brunswig 2003). But by 10,000 BP, the users of Agate Basin, Hell Gap, Cody, and Jimmy Allen projectile points had become regular visitors to the Front Range crest (Benedict 1992a, 2005a; Pitblado 2000). Seasonal occupation continued with varying intensity during the Archaic and Late Prehistoric stages. The most recent native people to utilize the high mountains were the Arapahos and Utes.
Few regions in North America are as ecologically diverse as the Colorado Front Range. In contrast to the Central Canadian Arctic (where Inuit hunters might live their entire lives without seeing a tree), or the eastern United States (where it was difficult to escape the shadows of the forest), the extreme compression of Front Range environments made it possible to walk from alpine tundra to plains grassland in several days, crossing two forest ecosystems along the way. Each environment had its appropriate season and unique resources, encouraging a lifeway of seasonal transhumance (sensu Davis 1963). Carefully scheduled movement through a landscape with more than 2500 m of relief was the key to successful use of the region (Benedict 1992b). Late summer and early autumn were the optimum seasons to be in the high country. Winter and spring were the seasons to avoid it.
Good winter campsites were essential. The best are near permanent streams in the hogback zone along the east flank of the Front Range. Here, steeply dipping sandstone outcrops provide sheltering overhangs. Groves of floodplain cottonwoods supply fuel. Because the hogback zone is in the winter precipitation shadow of the mountains, snowfall is typically light. Chinook winds, warmed by their descent from the Continental Divide, keep temperatures moderate, streams open, and the ground largely free of snow in winter. Migratory game animals such as bighorn sheep, elk, mule deer, and bison regularly wintered in the vicinity (Armstrong 1987). Even in the lower foothills, however, severe springtime snowstorms could bring activities to a halt, causing massive game-animal mortality and human suffering (Benedict 1999).
Native people are unlikely to have wintered in Middle Park, the intermontane basin west of the Indian Peaks (Figure 1). This basin registers some of the lowest air temperatures in the United States, and remains deeply snow-covered in many winters. But it is rich in resources, and was visited at other seasons.
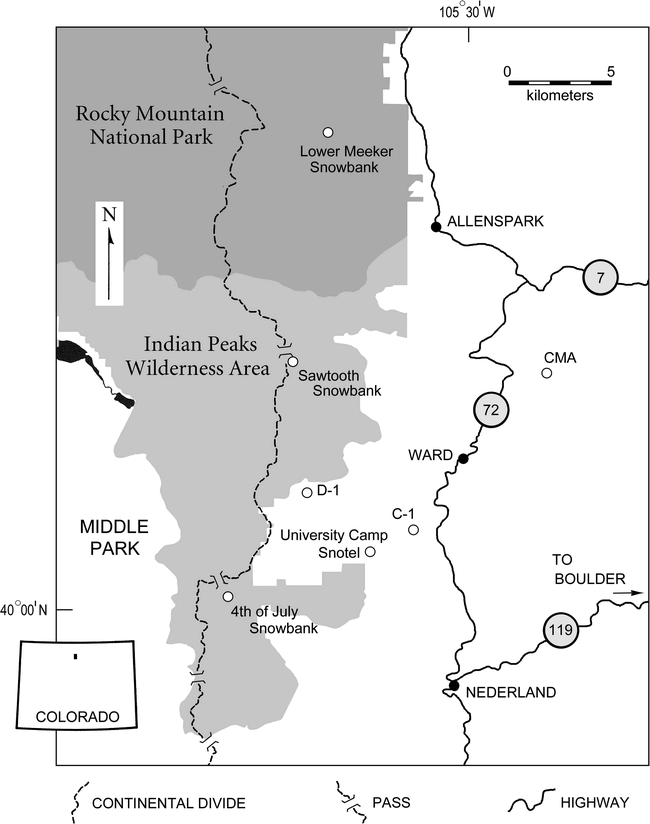
Map of the study area, showing the locations of weather stations and of snow patches with long meltout records. Illustration by James Benedict.
Three seasonal transhumance systems have been tentatively identified in the region (Benedict 1992b, 2000a). The two simplest systems involved up-and-down seasonal migrations, one originating at winter camps in the eastern foothills of the Front Range and the other in sheltered canyons and pinyon-juniper woodlands in western Colorado or adjacent Wyoming. A more elaborate system involved a 300 – 400 km counterclockwise migration in which people who wintered in the eastern foothills traveled northward along the mountain front in spring, then crossed the Divide westward through mountain passes that become snowfree several months earlier than those in the Indian Peaks and Rocky Mountain National Park. In summer, these groups drifted southward through North Park and Middle Park, where toolstone supplies could be replenished at traditional quarries, and where game animals and wild plant foods were plentiful. In late summer or early autumn, the people entered the high mountains from the west, hunting at alpine-tundra game-drive systems. As the first snows of autumn began to accumulate, they abandoned the mountains to re-establish winter camps in the eastern Front Range foothills.
Accurate population estimates are not available for the timberline region. Numbers undoubtedly fluctuated with changes in climate, as indicated by high frequencies of archeological radiocarbon dates during intervals thought to have been warmer and drier than the present (Benedict 1979), and low frequencies during periods of lingering snow (Benedict 1999). The existence of elaborate game-drive systems with numerous blinds (Benedict 1996; Cassells 2000) implies seasonal aggregation of people into relatively large bands (Benedict 1992b).
Hunting was the region's principal attraction. More than 50 stone game-drive systems have been recorded in the alpine tundra and forest-tundra ecotone east of Middle Park. Rock walls and cairn lines were constructed at these sites to maneuver large game animals to within range of hunters concealed in natural or manmade blinds (Benedict 2005b). First used at high altitude during the Late Paleoindian Period (Benedict 2000b), the technique remained popular until the coming of the horse, when it became rapidly obsolete (Benedict 1992b). Drive systems were probably also used in the subalpine forest, but would have been constructed of wood rather than stone (Hearne 1911; Honigmann 1954; McFee 1981), leaving no long-term archaeological record.
Gathering of wild plant foods is not nearly as well documented as hunting. This paper describes a study designed to assess the importance of plant foods in the diets of native people at high altitude, and to develop a basis for predicting the effects of climatic change on plant-food availability. It does so by (a) identifying the principal edible plants in an alpine-tundra and subalpine-forest study area; (b) monitoring their emergence, blooming, and ripening during a time period long enough to include years with markedly different growing conditions; (c) using these data to identify the principal climatic factors that influence plant-food availability in the region; and (d) estimating the overall contribution of plant foods to the diets of high-altitude hunter-gatherers, based on ethnographic analogy with human groups living in comparable environments at the beginning of the contact period.
The Research Area
The research area (Figure 1) slopes eastward from the Continental Divide, through the Indian Peaks Wilderness and Rocky Mountain National Park, to ranchland and non-wilderness portions of Arapaho-Roosevelt National Forest. Two major vegetation zones and two ecotones are represented.
Above tree limit, at altitudes of 4200 – 3500 m, remnants of a gently rolling early-Tertiary landscape still survive. Uplifted during Miocene and Pliocene time, the landscape was deeply dissected by Pleistocene valley glaciers (Benedict 1991). Most of the snow that falls on the tundra uplands is redeposited by wind in sheltered depressions, forming glaciers and late-lying snowbanks that supply meltwater to high-altitude wetlands throughout the summer. Despite episodic disturbance by frost processes (Benedict 1970), the alpine tundra supports more than 250 species of grasses, sedges, and herbs (Komárková 1979; Marr 1961; Willard 1979), with a high proportion of arctic-alpine and circumpolar taxa. It provides summer forage for bighorn sheep and elk, a major attraction to native hunters (Benedict 2005b). Bison also utilized the tundra uplands (Lee et al. 2006), although details of their hunting are uncertain.
Tree islands of climatically stunted krummholz (Holtmeier 2003), set in a matrix of alpine-tundra vegetation, dominate the landscape at altitudes of 3500–3350 m. This is the forest-tundra ecotone—a preferred summer camping location due to its expansive views of the surrounding countryside, shelter from the wind, and convenient access to firewood, water, and the food resources of two major vegetation zones (Benedict 1975).
The subalpine forest, dominated by Engelmann Spruce (Picea engelmannii [Parry] Engelmann) and Subalpine Fir (Abies bifolia A. Murray), occurs at altitudes of 3350 – 2850 m. The upper margin of the forest serves as a broad snow fence, accumulating large quantities of snow blown eastward from the tundra in winter. Beyond the zone of wind-enhanced precipitation, conditions become increasingly xeric, and the forest is subject to stand-replacement wildfires. Lodgepole Pine (Pinus contorta Douglas) and Quaking Aspen (Populus tremuloides Michaux) are successional after fire and other disturbances. Groves of Limber Pine (Pinus flexilis James) occupy exposed, windy locations. The subalpine forest can be uncomfortably hot in summer, with limited air movement, voracious mosquitos, tangles of deadfall timber, and a shortage of smooth, level ground. Evidence for prehistoric human use is concentrated in meadows along Pleistocene outwash terraces in valleys that lead to and from high mountain passes.
Large terminal moraines mark the outer limit of Pleistocene valley glaciation and the approximate upper limit of a diffuse ecotone between subalpine- and montane-forest vegetation. Within this ecotone, at altitudes of 2550 – 2850 m, Engelmann Spruce and Subalpine Fir are restricted to cool, moist microenvironments. Montane-forest species such as Ponderosa Pine (Pinus ponderosa Douglas) and Douglas-fir (Pseudotsuga menziesii [Mirbel] Franco) occur on drier slopes, with lodgepole pine and aspen in successional contexts. The change from subalpine to montane-forest vegetation coincides with a shift from the winter-spring precipitation maximum of the high mountains to the spring-summer maximum of the lower foothills and plains. Archeological sites are less common in the ecotone than might be expected from its biological diversity. They are most numerous along prehistoric travel corridors.
Climatological data (Figure 2) are available for four weather stations in the study area (Figure 1). The highest station, operated continuously since 1951 by the University of Colorado's Institute of Arctic and Alpine Research, is on a tundra ridgecrest east of Navajo Peak (D–1, 3750 m). A short distance to the southeast, on the floor of a glaciated valley in the upper part of the subalpine forest, the University Camp Snotel (UCMP, 3140 m) is maintained by the National Water and Climate Center, USDA. Winter precipitation at this location is augmented by windblown snow scoured from surrounding uplands. The Institute of Arctic and Alpine Research operates a second subalpine-forest station (C–1, 3050 m). This station receives less windblown snow than University Camp due to its ridgetop location and greater distance from the Continental Divide. A shielded rain gauge maintained by the Center for Mountain Archeology on an outwash terrace of South St. Vrain Creek (CMA, 2565 m) provides precipitation data for the lower part of the research area. Daily air temperature measurements were discontinued at this locality at the end of 1996, so NOAA/National Weather Service data for Boulder (1670 m) are included to illustrate the relationship of temperature to altitude on the east slope of the Front Range (Figure 2, c).

Climatological data for the study area. Stations D–1 (3750 m) and C–1 (3050 m) are operated by the Institute of Arctic and Alpine Research (University of Colorado 2006). The University Camp Snotel (UCMP, 3140 m) is operated by the U. S. Department of Agriculture (National Water and Climate Center 2005a, 2005b). Data for the Center for Mountain Archeology (CMA, 2565 m) are on file at the Center, together with meltout dates for the 4th of July and Sawtooth Snowbanks. Meltout dates for the Lower Meeker Snowbank were provided by Dr. Willam C. Rense. Temperature data for Boulder (1670 m) are from the NOAA-CIRES Climate Diagnostics Center (2005).
Meltout dates for three high-altitude snowbanks (Figure 2, d) provide an integrated measure of snowfall and growing-season temperature. The snowbanks occupy topographic recesses where windblown snow accumulates deeply. The 4th of July Snowbank is located in the forest-tundra ecotone on the floor of a valley east of Mount Neva. The Sawtooth and Lower Meeker Snowbanks are located above tree limit on the lee (east) faces of prominent summits. Because these snowbanks rarely experience carryover snow from year to year, their meltout dates closely reflect conditions during individual water years.
Sources of Information on Wild-plant Use
Few people can accurately describe the diets of their ancestors six generations ago, when Native Americans last practiced a hunting-gathering lifestyle in the study area. Northern Cheyenne elders interviewed by Hart (1981) in the 1970s often remembered the medicinal and ceremonial uses of plants, but rarely recalled details of their dietary use. Loss of traditional plant-food knowledge was hastened in the Front Range by the relocation of native peoples to warmer, drier environments with different assemblages of edible plants. Ethnobotanical studies during the early reservation period provide useful information on species such as serviceberries, chokecherries, and wild plums (whose ranges extend upward into the lowest parts of the study area), but little or no information on the alpine and subalpine species utilized during earlier years.
In 1914 the Colorado Mountain Club brought two Arapaho elders, Sherman Sage and Gun Griswold, to participate in a two-week pack trip through the region destined to become Rocky Mountain National Park. Both men had spent summers in the area when they were young. They were accompanied on the trip by Tom Crispin (a bilingual Arapaho), Shep Husted (a knowledgable outfitter and guide), and Oliver Toll (a Williams College undergraduate who served as impromptu ethnographer) (Benedict 2001). No Arapaho women were included in the party, and Toll's published account of the trip (Toll 1962) predictably emphasized place names, hunting successes, trails, battles, and legends. Although Toll did not mention plant-food use in his publication, he provided Alfred L. Kroeber with a brief typescript report listing Arapaho plant names, and occasionally uses (Toll 1914). The list identified four species as having edible berries, two species with edible roots (one described only as “? little white flat-top flower”), and four species whose leaves or bark were brewed for tea. Of the one-hundred geographic locations whose Arapaho names were identified in dictaphone recordings made in Fort Collins after the pack trip had ended (transcribed and translated by Andy Cowell and Alonzo Moss), only two were associated with edible plants: a mountain where “white turnips” were gathered, and a place where buffaloberries (soapberries) were plentiful. To the best of my knowledge, this is the full extent of the high-altitude Front Range ethnobotanical record.
To supplement this meager information, I turned to British Columbia, Alaska, and the Canadian Arctic, which share many plant genera and species with the Front Range subalpine forest and alpine tundra. Unlike the former native inhabitants of the Southern Rocky Mountains, First Nations people still commonly live in their traditional homelands. They continue to use wild plant foods today, recall their former uses, or have learned of their uses from tribal elders. The ethnobotany of these regions has been studied intensively, particularly by Turner (1995, 1997), Turner et al. (1990), and Kuhnlein and Turner (1991). Many of the plant-food species discussed in these studies also occur in the Colorado mountains. It would be surprising if they were not used for similar purposes, and prepared with similar techniques, in both regions. Table l lists what I believe were the most important food plants available to native people at altitudes above 2550 m in the Front Range study area. Color photographs of the plants are available on compact disk, together with details of their collection and preparation (Benedict 2007). Representative species are illustrated in Figures 3 – 5. Species names follow Weber and Wittmann (1992, 1996). Some Front Range taxa have recently been differentiated from their boreal-forest and arctic-tundra counterparts based on differences in chromosome numbers (Weber and Wittmann 1996). But from a practical standpoint, the species are so similar in appearance and taste that a forager would find such differences irrelevant.
Edible plants and lichens of the Front Range study area.


Edible greens and root foods in the study area: (a) Oxyria digyna; (b) Heracleum sphondylium; (c) Cardamine cordifolia; (d) Equisetum arvense; (e) peeled Erythronium grandiflorum bulbs; (f) Erythronium grandiflorum. Photographs by James Benedict.

Edible root foods in the study area: (a) Bistorta bistortoides; (b) Calochortus gunnisonii; (c) Bistorta vivipara, (d) Claytonia megarhiza, (e) Oreobroma pygmaea, (f) O. pygmaea roots (n = 887) cached in the subnivean tunnels of a montane vole. Photographs by James Benedict.

Edible fruits and seeds in the study area: (a) Vaccinium cespitosum, (b) Gaultheria humifusa, (c) Ribes inerme; (d) Pinus flexilis; (e) Helianthella quinquenervis; (f) Luzula parviflora. Photographs by James Benedict.
In cases in which names have recently been changed, I include their earlier, more familiar names in Table 1. Several species of greens are included because of their flavor and abundance, even though there is little or no ethnographic evidence of their consumption by native peoples. Designation of plants as “emergency foods” or “starvation foods” (Table 1) is subjective because species reserved for times of hardship by some groups were eaten routinely by others (Turner and Davis 1993). Medicinal plants, plants with ceremonial significance, dye plants, and plants used for basketry, cordage, utensils, weaponry, and other non-food purposes were not included in the study.
Seasonal and Annual Availability of Wild Plant Foods
Introduction
Over the course of eleven growing seasons (1994 – 2004), I monitored the emergence, blooming, fruiting, and ripening dates of edible greens, root foods, seeds, and fruits. I made regular observations at all altitudes in the research area. My list of edible plants had not fully evolved by the summer of 1994, and several species were not monitored until the following year. I did not discover scarce Front Range stands of Claytonia lanceolata until 2000, but have included five years of data because the species is so important in northerly regions. Figures 6 – 8 summarize my observations on the phenology of selected edible plants. Emergency foods are excluded because they are available throughout the year (Amerosedum lanceolatum, Bryoria fremontii, Umbilicaria spp.) or because they are most suitable for use after they have overwintered beneath snow (Arctostaphylos uva-ursi, Juniperus communis).

Graphs showing the emergence dates of greens (a–c), the seasonal availability of fertile horsetail shoots, and the bloom periods of important root-food species above timberline (e–h).
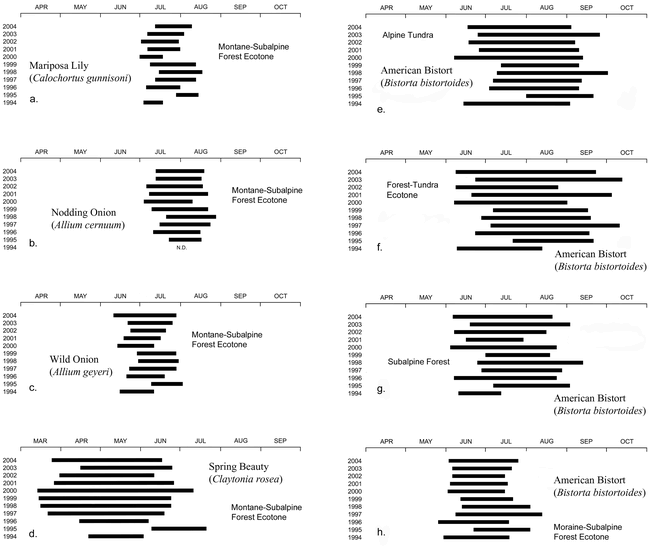
Graphs showing the bloom periods of root foods in the lower part of the study area (a–d) and of Bistorta bistortoides throughout the altitudinal range of the species (e–h). Note that the record for Claytonia rosea (d) begins in March, a month earlier than the other recordws.
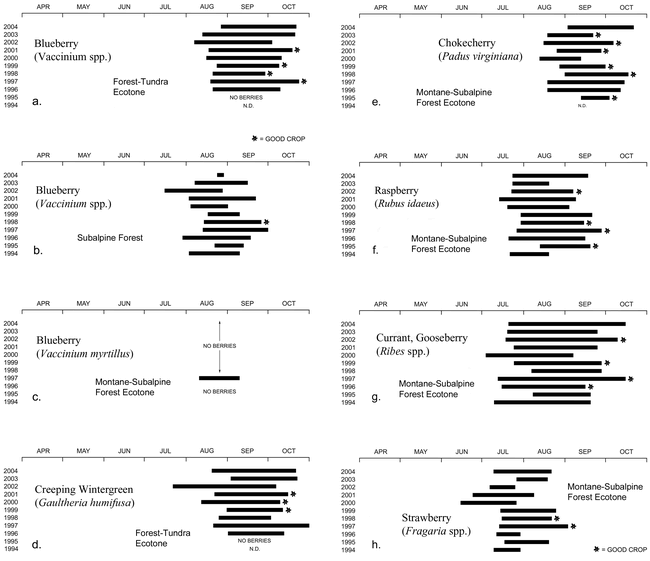
Graphs showing the seasonal availability and productivity of ripe Vaccinium spp. berries in three ecosystems (a–c), ripe Gaultheria humifusa berries in the forest-tundra ecotone (d), and the fruits of four species that grow near the lower boundary of the study area (e–h).
Climatological data from study-area weather stations are summarized in Figure 2. The summaries are based upon annual water years (October–September), which are more relevant than calendar years to plant growth. September is treated as a growing season month because of its importance to the ripening of high-altitude fruits, though it does not meet Binford's (2001) temperature-based definition, and in some years experiences hard frosts and early snows. The general trend between 1994 and 2004 was one of decreasing winter snowfall (Figure 2, a) and increasing growing-season rainfall (Figure 2, b). Altitudinal differences in precipitation were greater during the snow-accumulation season than during the growing season (Figure 2, a and b). Growing-season temperatures (Figure 2, c) showed similar trends at all altitudes. Snowbank meltout dates (Figure 2, d) were correlated more strongly with winter precipitation than with summer temperature, though both factors were involved.
The study period proved ideal due to the extremes of climate represented. The winter of 1994 – 1995 was one of the snowiest in recent memory and was followed by a cool, moist summer unfavorable for melting. The winter of 2001–02 produced relatively little snow and was followed by a hot, dry summer. Differences in plant-food availability during these contrasting water years provide a basis for estimating the availability of edible plants during former climatic intervals. Simple climatological data such as these, however, fail to predict the productivity of fruit and seed crops in many years.
Greens
Green vegetables are the first plant foods to become available in spring. Their leaves and stems can be gathered and eaten within a few days of emergence. Most species remain edible throughout the summer, though they become tougher and less palatable with age. Bittercress (Cardamine cordifolia), Alpine Sorrel (Oxyria digyna), and Lovage (Ligusticum tenuifolium) are still flavorful in October. How late in the season a green vegetable continues to be eaten depends mainly on how hungry the gatherer is, and on the availability of other foods. For this reason I have plotted in Figure 6, a–c, only the dates when shoots first become large enough to warrant harvesting: e.g., reach a height of about 5 cm. Horsetail (Equisetum arvense) is not strictly a green vegetable, but was included in this category because its brown, fertile shoots were prepared similarly to greens. Bar graphs are used for this species (Figure 6, d) because the shoots wither and dry a few weeks after emergence.
Greens become available much earlier in the lower part of the study area than above timberline (Figure 6, a–c). Annual differences correlate closely with snowbank meltout dates (Figure 2, d). The influence of snow is illustrated by comparing the snowy water year of 1995 to the dry water years of 2000 and 2002. In comparison to the drought years, heavy snowfall delayed the emergence of greens by at least 1 month in the montane/subalpine forest ecotone and 1½ – 2½ months above timberline (Figure 6, a–c).
Root Foods
Edible corms, bulbs, tuberous roots, taproots, and rhizomes are known collectively as “root foods.” Some root-food species, such as the Yellow Avalanche Lily (Erythronium grandiflorum), Mariposa Lily (Calochortus gunnisoni), and Nodding Onion (Allium cernuum), have distinctive seed stalks that remain visible until they are crushed by autumn snows. They can be located and harvested until the ground freezes or the plants become hidden beneath deep snow. Other species, such as the Indian Potato (Claytonia lanceolata), Spring Beauty (Claytonia rosea), and Pygmy Bitterroot (Oreobroma pygmaea), have succulent leaves that wither and die within a few days of blooming, making them virtually invisible. Their roots were traditionally harvested during the flowering season (Turner et al. 1990). Because all Front Range root-food species are conspicuous bloomers, and can be gathered most efficiently when in flower, I have chosen the bloom period as the most consistent measure of year-to-year variability (Figures 6, e–h; 7, a–h).
Claytonia rosea, which grows in the lower part of the study area, is the first of the root foods to blossom. It does so at a season when heavy snows still occur frequently. Snow puts the maturation process temporarily on hold, extending the bloom period to as much as 4 months (Figure 7, d). The plants, however, often are hidden from view. During the spring of 1995, snow was so deep and persistent that C. rosea did not emerge and flower until early June, almost 3 months later than it did in 2000 (Figure 7, d).
Three important high-altitude Front Range root foods—Erythronium grandiflorum (Figure 3, e–f), Claytonia lanceolata, and Oreobroma pygmaea (Figure 4, e)—are restricted to areas of late-lying snow, where they bloom within a few days of emergence. The speed with which these species complete the aboveground portions of their life cycles may be a strategy (a) to avoid competition with more-aggressive, slower-maturing species, (b) to take advantage of the flush of soil moisture that accompanies meltout, or (c) to ensure long-term survival in an environment where growing seasons much shorter than those of today can be expected to occur. When small mammals such as Montane Voles (Microtus montanus nanus) overwinter in snowbed plant communities (Benedict and Benedict 2001), they commonly decimate root-food supplies (Figure 4, f). The blooming schedules of all three snowbed species show a predictable relationship to annual snowfall (Figure 2, a) and to snowbank meltout dates (Figure 2, d). In 1995, their bloom was delayed approximately 1½ months relative to 2002 (Figure 6, e–g).
The alpine spring beauty, Claytonia megarhiza (Figure 4, d), can tolerate an unusually broad range of snow-cover conditions, so is found blooming at one place or another for much of the summer (Figure 6, h). Conspicuous leaves make the plants easy to locate even when blooms are absent. The large, fibrous taproots of this species can be gathered throughout the growing season.
American Bistort, Bistorta bistortoides (Figure 4, a), is the most widely distributed Front Range root food. It occurs at all altitudes in the study area, and in environments as diverse as windswept tundra ridges, protected recesses containing moderately late-lying snow, and montane wetlands. Stands are largest, and rhizomes shallowest and easiest to dig, above tree limit—an advantage partially offset by the decrease in rhizome size that occurs with increasing altitude (Figure 9). Bloom begins in the alpine tundra only a few weeks later than it does in the lower part of the study area (Figure 7, e–h), reflecting the near-synchronous exposure of meadows that are blown free of snow in winter, regardless of their altitude. Bloom persists longest above timberline (Figure 7, e–f) due to the species’ ability to grow and to flower in areas where snow disappears late in the summer.
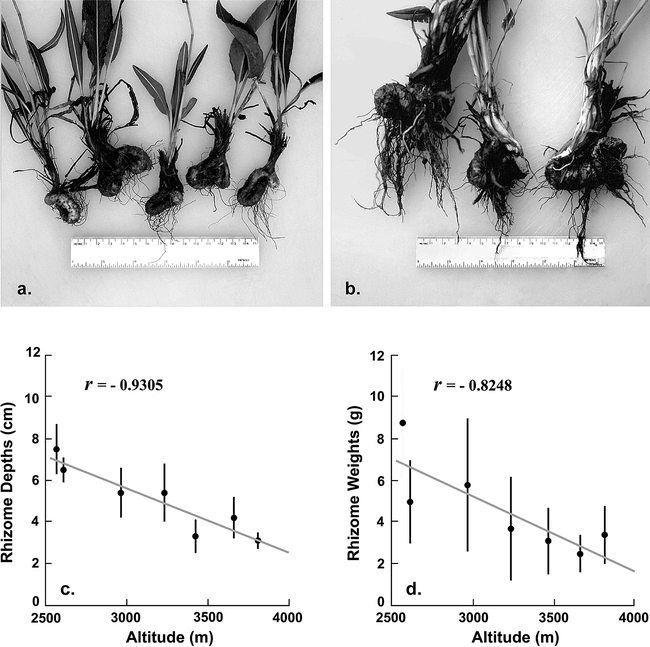
Photographs of Bistorta bistortoides rhizomes collected at altitudes of 3810 m (a) and 2560 m (b). Graphs show the relationship of rhizome depths and weights to altitude (c–d). Each data point is the average of 20 or more measurements. Vertical bars represent one standard deviation (not available for weight at the lowest collecting locality). Photographs by James Benedict.
The blooming seasons of Calochortus gunnisoni, Allium cernuum, and Allium geyeri correlate closely with snowmelt patterns, and are completed within 1½ months of emergence (Figure 7, a–c).
Fruits
It would be difficult to overemphasize the importance of fruits to native peoples. Prime berry patches “were named, owned, managed, and celebrated as places” in Southeast Alaska, where fruits such as soapberries (Shepherdia canadensis) played a symbolic role in Tlingit ceremonial feasts (Thornton 1999). Elaborate technologies were developed to prepare berries for storage (Mack and McClure 2002). Year-to-year differences in the availability of ripe fruits in the study area are summarized in Figure 8, together with subjective assessments of their productivity.
Blueberries (Figure 5, a) illustrate the challenges associated with fruit production at high altitude in the Colorado Front Range (Figure 8, a–c). Wild blueberries have been studied in detail elsewhere, due to their commercial value (Northwest Berry and Grape Information Network 2005; University of Maine Cooperative Extension Service 2005). The studies show that favorable temperature and moisture conditions must persist long enough during the preceding autumn for primordial flower buds to develop in large numbers. Because blueberries are pollinated primarily by bees (which are inactive during wet weather), the bloom period must be relatively dry. The adverse effect of drought during fruit development and ripening is a critical limiting factor. Near the lower altitudinal limit of blueberries in the study area, ripe fruits were available in only one of eleven years (1997), and in that year were present in token quantities. In the Front Range subalpine forest, where blueberries are the dominant understory species, ripe berries were bountiful only in the wet summer of 1998. Good blueberry years were more frequent in the forest-tundra ecotone, due at least partly to the influence of snowmelt seepage, which ameliorates the effects of summer drought without hindering pollination. Ripe blueberries were locally abundant in the ecotone during the 1997, 1998, 1999, and 2001 water years. All four years experienced above-average precipitation during fruit set and ripening (Figure 2, b). But in 2004, the wettest summer of the study period, crops were poor. A hard frost occurred in late August of that year, but by this time the berry crop already appeared destined for failure. The problem in 2004 appears to have been the distribution of summer moisture, most of which fell during the pollination season. Unusually short growing seasons can also curtail blueberry production. An example was the summer of 1995, when a promising (but delayed) crop of green blueberries did not have time to ripen in the forest-tundra ecotone. With so many opportunities for failure, it is small wonder that good blueberry years are uncommon in the Front Range.
Like blueberries, other fruit-bearing species need favorable conditions at many stages in their seasonal cycles. The factors that cause them to succeed or fail are not always apparent. Fruit productivity in the lower part of the study area (Figure 8, e–h) showed little consistency from species to species, and no clear correlation with the simple climatic data available for the study period (Figure 2). My notes on fruits are rife with statements such as “withered before ripening” (Amelanchier alnifolia; 2000, 2002), “very abundant but soon frost damaged” (Padus virginiana; 1995), “scarce after profuse bloom” (Ribes spp.; 2000), and “poor crop due to summer drought and galls” (Padus virginiana; 1996). Compared to the berry-friendly Pacific Northwest, the Front Range is a difficult environment for fruits. Successful production depends on disparate factors that cannot be anticipated even in years with good bloom. It would have been difficult for native people to predict the quality and abundance of a fruit crop in advance, and to use this information in scheduling their seasonal travels. Like many root foods, blueberries benefit from a regimen with frequent fires. The fact that fires are no longer set deliberately for crop improvement (see Turner 1991) may be one reason that blueberries are so rarely productive in the subalpine-forest zone today.
Seeds
The seeds of most high-altitude plants are too small and too heavily predated by birds and mammals to be important food sources. Limber Pine (Pinus flexilis), which grows abundantly in the study area (Figure 5, d), is an occasional exception. Its large and nutritious seeds are aggressively harvested by Clark's Nutcrackers (Nucifraga columbiana). Humans, however, could minimize the effects of avian competition by picking the cones in their ripe (but still closed) state, and opening them artificially with the aid of heat. Elsewhere in the West, this strategy was used to harvest the seeds of Whitebark Pine (Kuhnlein and Turner 1991) and Piñon Pine (Bettinger 1991). Limber Pine nuts require two consecutive favorable years for cones to form and seeds to ripen. Like fruits, they are an unreliable food source at high altitude. Only the 1998 and 2003 harvest years were productive in the subalpine forest and forest-tundra ecotone. Drought in the summer of 2000 (Figure 2, b) left trees covered with sterile, aborted cones that had formed in profusion during the preceding year.
Three other species with edible seeds are Bistorta vivipara (Figure 4, c), Helianthella quinquenervis (Figure 5, e) and Luzula parviflora (Figure 5, f). These were undoubtedly eaten as snacks, but are unlikely to have been an important food source.
Role of Wild Plant Foods in the High-altitude Front Range Diet
How important were wild plant foods to the hunter-gatherers who utilized the study area during warm seasons of the year? Figure 10, based on tabular data in Binford (2001), provides an approximate answer. It shows the relationship between (a) estimated use of terrestrial plant foods by 265 hunter-gatherer groups at the time of initial European contact and (b) the effective temperature of the environment in which each group resided. Effective temperature (ET) was chosen as a measure of warmth because it is thought to have greater biological significance than mean annual temperature (Bailey 1960). It can be calculated from weather records using the equation in Table 2. Dietary estimates (Binford 2001:Table 5.01) are based upon data in Murdock (1967), supplemented by Binford's own summaries of ethnographic monographs, reports, and other records. Some of the human groups included in the study had already begun to use domesticated plants by the time of first direct European contact (Binford 2001:117), but this was not true of groups living in regions with ET values as low as those in the Front Range study area. The relationship between plant-food consumption and warmth is unmistakable (Figure 10).
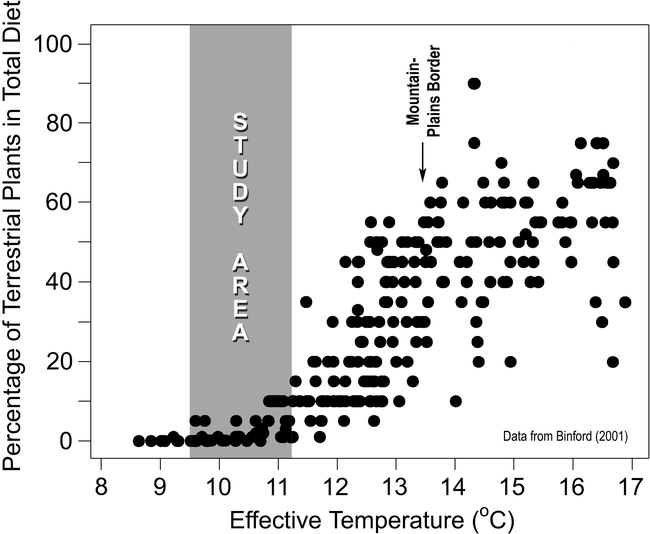
Role of terrestrial plant foods in the annual diets of 265 hunter-gatherer groups in North and South America, Eurasia, Africa, and Australia at the time of first European contact, plotted as a function of effective temperature. The graph is based upon data presented in tabular form by Binford (2001:Tables 4.01, 5.01). The range of ET values within the shaded area reflects temperature data from study-area weather stations. Extrapolation to higher altitudes suggests values as low as 9.0°C along the Continental Divide.
Effective Temperatures and Growing-season Lengths.

Based on weather-station temperature data, modern ET values in the study area range from 9.5°C to 11.2°C (Table 2). Extrapolation suggests that values as low as 9.0°C should be expected at altitudes above 4000 m. Sixty-one of the groups summarized by Binford (2001) lived where ET values fall within this range. The groups were distributed throughout the North American and Eurasian Arctic, the Subarctic, and Tierra del Fuego. Although some groups were migratory, their seasonal travels were limited to cold environments, with access to fresh, terrestrial plant foods for only 2 – 3 months of the year. Subsistence was heavily biased toward hunting and fishing, with plant foods accounting for a scant 0 – 10% of annual diet (Figure 10).
Assuming an average ET value of 10°C for the study area, the data in Figure 10 suggest that wild plant foods would have constituted only 0 – 5% of annual diet. This, of course, is an underestimate due to the close proximity of warmer environments. There was no need in the Front Range for people to conserve a proportion of their high-altitude harvest for use in winter and spring, as was the case in the arctic and subarctic. If all of the greens, roots, berries, and seeds gathered in the study area during the 3-month growing season were consumed in the high country, the estimate based on Binford's data would need to be quadrupled. Thus plant foods might contribute as much as 20% to summer diet along the crest of the Front Range. To the extent that high-altitude plant foods were preserved for winter use, this percentage would need to be lowered.
On an annual basis, Front Range hunter-gatherers clearly were not plant-food deprived. During the course of their seasonal travels, users of the alpine region visited a much broader range of environments than were available to year-round residents of the arctic and subarctic. Each environment had its appropriate season and plant-food resources. The annual rounds of people who visited the crest of the Colorado Front Range in summer led them at other seasons to regions with ET values as high as 13 – 14°C (Table 2). Here plant productivity was greater, and the growing season longer, than in the study area. Because we do not know exactly how Front Range people apportioned their time altitudinally, it is not possible to estimate plant-food consumption on an annual basis. Clearly, however, the ability to migrate to warmer environments extended the effective growing season to at least seven months, greatly increasing opportunities for gathering. People could spend spring and early summer in the mountain parks, where Balsam-root (Balsamorhiza sagittata [Pursh] Nuttall), Bitterroot (Lewisia rediviva Pursh), Yampa (Perideridia gairdneri [Hooker & Arnott] Mathias), Avalanche Lily (Erythronium grandiflorum), and Indian Potato (Claytonia lanceolata) begin to bloom in late May or early June. From there they could move to the high mountains, enjoying a second harvest of Avalanche Lily bulbs and Spring Beauty corms, and gathering root foods, fruits, and medicinal plants that were available only at high altitude. On their return to lower, milder environments in autumn, they had access to ripe Chokecherries (Padus virginiana), Serviceberries (Amelanchier alnifolia), and wild plums (Prunus americana), whose ranges barely extend into the study area from below. Thus hunter-gatherers in the Southern Rocky Mountains could increase plant-food availability in ways that were impractical for year-round residents of arctic and subarctic latitudes, where distances were prohibitive.
Important root foods such as the Nodding Onion (Allium cernuum) and Yellow Avalanche Lily (Erythronium grandiflorum) require lengthy pit cooking to break down inulin (a nondigestible polysaccharide) into its constituent fructose units (Peacock 1998). In regions where roots of these species were abundant, they were processed in earth ovens or on stone cooking platforms, then stored in cache pits for winter and spring consumption, or dried for transport or trade. The ovens were typically 1.5 to 4.5 m in diameter, with depths of 10 to 100 cm (Alexander and Matson 1987; Francis 2000; Matson and Alexander 1990; Peacock 1998; Pokotylo and Froese 1983; Smith 1974; Thoms 1989; Vanags 2000; Wissler 1910). No ovens approaching these dimensions, and no cache pits whatsoever, have been found within the study area. Twelve stone-filled earth ovens have been excavated at high-altitude campsites (Benedict 1985, 1990, 2000a, 2002), but all were small features, ranging from 30 to 68 cm in maximum diameter and 6 to 20 cm in maximum depth. They suggest use by single-family groups to prepare food for immediate consumption (Figure 11). If roots were collected in the mountains for storage or trade, the lack of archaeological evidence for large-scale processing suggests that they were dried (leaving no visible trace) or were carried to lower altitudes for processing in ovens.

Plan and cross-sectional views of a Late Archaic oven in a wetland at site 5BL82, Middle St. Vrain Valley, Colorado Front Range (Benedict 2002). The feature was floored with charcoal and filled with charcoal-stained earth and cobble-sized stones (n = 24). Like other excavated earth ovens in the study area, it appears to have been too small to use in large-scale root-food processing. Photographs by James Benedict.
The list of forty edible plants and groups of closely related plants discussed in this paper (Table 1) is by no means comprehensive, but is thought to include all of the most important potential food species. Some taxa, such as Viviparous Bistort (Bistorta vivipara, Figure 4, c), served multiple purposes. Thirteen species, however, would have been gathered primarily for their roots, nine for their fruits, eight for their leaves and/or stems, and four for their seeds. Six were probably reserved mainly for times of food scarcity.
How do these figures compare with estimates from other environments? Grinnell (1923a: 250) reported that Cheyenne women interviewed during the reservation era “speak of about 35 or 40 plants that serve them as food, of which 16 or more are fruits, and eight or ten roots.” Carlson and Jones (1940) list 34 food plants used by the Comanches in Oklahoma. Fifteen were fruits, 14 were root foods, and five were seeds or nuts. No green vegetables were reported, perhaps as an oversight. According to Julian Steward (1938), the Northern Paiutes of Owens Valley, in southern California, used at least 55 food plants—41 for their seeds, five for their roots, five for their berries, and four for their greens. And the Lehmi River Shoshonis of mountainous central Idaho harvested at least 38 edible plant species—20 for roots, 13 for seeds, three for berries, and two as green vegetables (Steward 1938).
Summary and Conclusions
The seasonal availability of potential wild-plant foods at high altitude in the Front Range of north-central Colorado was monitored during an eleven-year period. Regular observations were made in alpine tundra along the Continental Divide, and eastward as far as the lower limit of subalpine-forest vegetation. Medicinal and ceremonial plants were not studied, though they were doubtlessly an incentive for visiting the high mountains. The study area contains abundant evidence of prehistoric human use, including game-drive structures above timberline and campsites in the forest-tundra ecotone. Native people were removed from this part of Colorado before their lifeways could be studied, but analogy with similar environments—particularly mountain and arctic regions of Canada and Alaska—suggests that at least 40 plant species, or groups of closely related species, could have been used for food.
Emergence of greens and flowering of root-food species in the Front Range study area are influenced most strongly by annual snowmelt dates. Thus native people on the Plains and in Middle Park could predict their availability by scanning the mountain snowpack from a distance. Greens can be collected throughout the summer growing season, but would probably not in themselves have attracted people to the high country. Root foods are reliably available, though traditional favorites such as Erythronium grandiflorum and Claytonia lanceolata are smaller and less common at high altitude than in aspen forests along the margins of Middle and North Parks. Only two of the thirteen “root-food” species considered in this report are abundant enough to be likely candidates for large-scale harvesting. The most promising is Bistorta bistortoides, the rhizomes of which were traditionally dried for storage (Hart 1981:32), so would have left no archaeological record of their preparation. The other is Erythronium grandiflorum, which is abundant in some valleys, but not in all. The apparent lack of large roasting pits in the study area suggests that the bulbs of this species, if gathered for storage or trade, were processed at lower altitudes. More likely the bulbs were dug and prepared for immediate consumption in the mountains.
Fruit and pine-nut crops rarely are bountiful in the study area because they require favorable conditions at several stages in their development, beginning the preceding year. Dry summers are the most common cause of failure, but other causes exist. Blueberries did not ripen at high altitude in 1995 due to the short, cold, snowfree growing season. They failed to set fruit in 2004 due to untimely rains that inhibited insect pollination. Poor crops can also occur when conditions during the preceding autumn discourage development of premordial fruit buds. If I were a Front Range hunter-gatherer under conditions similar to those of the present, my philosophy would be to enjoy berries and pine nuts whenever I found them, but never to count on their availability.
The eleven-year study period included one water year with unusually late-lying snow (1995), and another with early-melting snow and summer drought (2002). These years represent opposite extremes in the range of climatological variation likely to have occurred in this area of Colorado during the past century and a half. During colder climatic intervals, such as the Little Ice Age, plant foods would have been even less abundant at high altitude. During warmer intervals, they would have been more abundant. Periods of summer drought or increased winter–spring snowfall would have impacted plant-food availability regardless of temperature. There is no shortage of limiting factors in the high-altitude environment.
Ethnographic and climatological data summarized by Binford (2001) suggest that game animals, rather than wild plant foods, were the study area's principal attraction. This is consistent with the abundance of stone hunting structures above timberline (Benedict 2005b) and the apparent absence of large root-processing ovens. Plants probably acounted for less than 20% of summer diet along the crest of the Front Range. But people spent much of the year in warmer, low-altitude environments, where fruits and seeds are more productive, and the growing season is longer. On an annual basis, plant foods were an important element in their diets.
Footnotes
Acknowledgments
William C. Rense provided meltout data for the Lower Meeker Snowbank (Figure 2, d). Andy Cowell alerted me to unpublished information gathered by Oliver Toll concerning Arapaho plant-food use in the region that would later become Rocky Mountain National Park. Transcriptions and translations by Cowell and Alonzo Moss of dictaphone recordings made at the conclusion of Toll's 1914 pack trip also were useful. Audrey Benedict, Andy Cowell, and an anonymous reviewer made helpful comments on the manuscript. When guidance was needed, as was often the case, I was fortunate to be married to one of the region's premier naturalists.