
Abstract
Dendroarchaeology became established with A.E. Douglass’ (1929) classic paper in National Geographic Magazine, dating approximately 45 ruins throughout the southwestern United States. Since then, dendrochronology has established the occupation sequences of numerous archaeological sites and has been used to describe past climatic conditions experienced by native and historical populations. The technique of dendrochronology has continued to expand and new methods are being developed that can provide ecological records that may help answer questions concerning anthropogenic ecology and resource availability in an archaeological context. The applications discussed in this paper include fire histories, stand-age structure, climate reconstructions, insect outbreak reconstructions, and mast (synchronous fruiting of trees) reconstructions. Native Americans’ effect on natural fire regimes is a debated issue that we can now explore through numerous fire histories extending back to the 1600s in the southwestern United States. Historical fire regimes can be examined in the eastern United States back to the 1800s. Climate reconstructions in the southwestern United States extend back more than 2,000 years in multiple locations. Native Americans have used Pandora Moth larvae as a food source, and outbreak reconstructions now extend to the 1300s in south-central Oregon. Furthermore, new techniques in dendrochronology have been developed to reconstruct masting in oak trees in the southeastern United States over the last century. These dendroecological records can provide useful information for resource availability if specific studies can be conducted that combine archaeological findings with these long-term reconstructions.
Introduction
Dendrochronology has gained wide acceptance in archaeology as an accurate tool for chronological control. Dendrochronologists have used tree rings to date the construction of archaeological structures (Dean 1978; Dean et al. 1985; Douglass 1929; Haury 1962), peel bark scars (a.k.a. culturally modified trees) from Native American use of the inner bark of pine trees (Kaye and Swetnam 1999), and even to verify the dating of historical works of art (Lavier and Lambert 1996) such as the panels in Dutch paintings (Eckstein et al. 1986) or the wood in violins (Grissino-Mayer et al. 2002). The use of dendrochronology to reconstruct environmental variables is becoming more common around the world and is not dependent upon preservation conditions in archaeological sites. The prevalence of these ecological records can extend the benefit of dendrochronology to archaeologists where preservation of wood in archaeological sites is rare. Dendroecology is a sub-discipline (established in the 1970s) of dendrochronology that uses tree rings to reconstruct environmental records (Fritts 1971; Fritts and Swetnam 1989). The products of dendroecological research have not been used to their full capacity in archaeological research. Archaeologists have recognized the applicability of dendroclimatic reconstructions of temperature and precipitation to explain environmental resource limitations and associated migration patterns (Ahlstrom et al. 1995; Dean et al. 1985; Grissino-Mayer 1995; Salzer 2000b; Salzer and Kipmueller 2005; Van West and Dean 2000) and have used stream flow reconstructions to provide paleoenvironmental information for prehistoric irrigation systems (Nials et al. 1989). Dendroecology can be used to develop further environmental records of fire history (Swetnam and Baisan 1996), insect outbreaks (Speer et al. 2001; Swetnam et al. 1985), and acorn production (Speer 2001). Researchers are developing long records of these variables that can be useful to archaeologists interested in anthropogenic ecology and resource availability.
Anthropogenic Ecology
Anthropogenic ecology is the study of the extent to which people have affected their landscape (Adams 2004; Minnis 2003; Wagner 2003). Resource availability is used in this paper to mean the natural resources that are accessible to human populations and is a logical extension of the concept of anthropogenic ecology. At the same time, a feedback exists between these two processes where human action on the landscape can modify resource availability. Human/environmental interactions encompass both anthropogenic ecology and resource availability as well as allowing for interactions such as human affects on resource availability (e.g., Hammett 1997; Nabhan et al. 1982; Rea 1979; Wagner 2003). In this paper, we will describe the records available from dendroecology and demonstrate how dendroecological records can examine landscape change due to human activity, be used to understand management processes (such as fire), and provide information on resource availability. These alternate lines of evidence may help in the interpretation of how humans have interacted with their environment and in the exploration of native peoples’ behavior (e.g., Dean 1997).
Dendroecology: Scope and Limitations
Data and Techniques
Dendrochronology is the general term for the study of annual growth rings in trees to provide chronological information or long-term records of environmental variables that affect tree growth. Depending upon the variables that are being recorded, sub-disciplines are identified, including dendroclimatology for the reconstruction of climatic variables or dendroecology for the reconstruction of ecological variables such as fire, insect outbreaks, or masting events (synchronous fruiting of trees). The key tenet of dendrochronology is the principle of crossdating where researchers match the patterns of wide and narrow rings between living trees in a modern chronology with remnant or archaeological wood to develop absolutely dated time series. Crossdating enables researchers to identify ring anomalies (such as locally absent rings or false intra-annual bands) in some individuals from a given location. The resultant chronologies provide annual and sometimes sub-annual resolution. Trees also incorporate everything that affects their growth so that they become recorders of the natural environment. Dendrochronologists can reconstruct these environmental variables (such as precipitation) given a calibration data set from which statistical models (using regression analysis) of the tree's response can be developed. We can then use that statistical model or a distinctive response of the tree (such as reduced growth due to the defoliation from an insect) to develop long-term reconstructions of that environmental variable.
Limitations
Reconstruction of environmental variables is restricted by the topographic position of sensitive trees (Fritts 1971), the availability of calibration datasets (Speer 2001), the age of the available trees, and the preservation of remnant wood. Trees are not ubiquitous on the landscape and even when they are present, dendrochronologists must choose specific trees that are sensitive to the environmental variable that they wish to reconstruct. Trees mainly respond to the variable that most limits their growth. Some sites will have adequate precipitation, for example, so that those trees are not limited by that particular variable and will not record annual changes in precipitation. Furthermore, many reconstructions depend upon calibration datasets such as modern fruiting records from the species of tree where mast reconstructions are desired. These calibration datasets are very rare, but without them, we cannot determine the response of the tree to a biological forcing factor and develop the resultant reconstruction (Speer 2001).
Age of trees and wood preservation can also limit the extent to which a reconstruction is possible. Most tree species can live to be hundreds of years old, whereas only 17 tree species throughout the world have been found with individuals that live to be over one thousand years old (Brown 1996). Tree-ring chronologies can be extended back in time by hundreds or thousands of years by incorporating dead wood from the same site through crossdating of remnant samples of fallen yet undecayed trees or from wood incorporated into dwellings. This enables dendrochronologists to develop chronologies longer than the lifetime of individual trees, but success of this process depends upon local preservation of wood. In the Eastern Deciduous Forest, for example, wood decays quickly on the forest floor, so that remnant wood does not remain on the site for more than 50 years in most cases. In comparison, remnant pieces of Bristlecone Pine wood in the White Mountains of California have been found to be preserved for many thousands of years (Hallman et al. 2006). Because of these constraints, dendroecological reconstructions are limited in length.
These dendrochronological tools can potentially develop very useful datasets for ethnobiologists, but much time, patience, and understanding of the tree as a biological recorder is needed to develop accurate chronologies. Not all sites record the variables of interest; considerable time must be invested to determine the drivers that affect growth at a particular site. The methods of dendrochronology are very exacting. Most of the researcher's time is spent looking through a microscope repeatedly analyzing rings and matching their patterns to other trees on the site. This pattern matching, or crossdating, is of utmost importance in dendrochronology because without accurate dating, the final chronology may be imprecise and misinterpretation of the data would be possible. We recommend collaboration between an archaeologist and a dendrochronologist in the development of these records so that a full understanding of the culture and of trees as environmental recorders can be brought to the study question.
Types of Tree-Ring Evidence
Dendroecological applications can provide information through different kinds of evidence and through different levels of analysis. Direct evidence can be obtained from scars present on the tree, such as fire scars or peel bark scars on culturally modified trees. Indirect evidence of environmental variables is gathered through the comparison of an affected population (such as a host tree species for insect outbreaks) versus an unaffected population (such as a non-host tree species) that is used as a control that represents the background climate signal. Some reconstruction techniques require a calibration dataset, where a known record of some environmental phenomenon is documented for a period of time and is used to understand the response of a tree species to that phenomenon. A regression analysis is performed to develop a statistical model that can then be used to reconstruct that phenomenon further back in time. Climate reconstructions, mast reconstructions, stand-age structure for anthropogenic affects, and some insect outbreak reconstructions require this calibration dataset. Stand-age structure analyses need a general forest control population in order to interpret anomalies in a given study location.
Even though fire scars are direct evidence, the interpretation of a fire history for a stand or the affect of anthropogenic use of fire relies on the interpretation of indirect evidence. Fire history can be obtained at the tree level, but is usually reported at the stand level where 20-30 trees have been sampled and the composite fire history for the stand is reported based on scars that are recorded on at least two trees. This enables the reconstruction of spreading fires that have affected multiple trees in the stand and excludes lightning strikes on individual trees, or possibly other scars that were mistakenly classified as a fire scar and only affected one tree. An interpretation of a fire as being of anthropogenic origin can usually only be made by observing a change in fire season, comparison with climate records, proximity to archaeological sites, and/or comparison with fire histories from throughout the region. A change in fire behavior on the fine scale may result in the interpretation of Native American use of fire near a habitation site and is another example of indirect evidence.
Teasing Apart Multiple Signals
Trees record anything that affects their growth, which makes them convenient recorders of environmental variables. This means, however, that trees may be recording climatic variations, mast events, and insect outbreaks all at the same time. Dendrochronologists use several techniques to differentiate these signals.
The

Where:
Rt is ring width at year t.
Gt is the age related growth trend.
Ct is climate at year t.
D1t is the endogenous disturbance within the stand.
D2t is the exogenous disturbance from outside the stand.
Et is the error term incorporating all of the signal that is not controlled for by the above variables.
This model simply states that ring width is a function of the age related growth trend, climate variability, endogenous disturbances (such as competition within the stand), exogenous disturbances (such as logging events), and the error term that incorporates all other signals recorded by the tree. Through mathematical treatment of the data, a dendrochronologist can control for the signal from these other factors to enhance the signal of interest. One example is the use of a mathematical model to remove the age-related growth trend. All other factors being equal, the width of rings will decrease from pith to bark. A pine tree growing without competition, for example, will put on a smaller ring each subsequent year because the tree is putting the same volume of wood on an ever-increasing cylinder. In order to remove the age related growth trend, the ring widths are standardized by fitting a negative exponential curve to the ring-width data and dividing the ring-width value by the model value. The result is an index chronology, which is a dimensionless measure of growth in each year. Then we can remove another signal, such as climate, by conducting a regression analysis where a climatic variable is the independent variable and the ring width is the dependent variable. The residual chronology is the value that is left after the climate data has been removed from the index chronology. This residual chronology can then be examined for a response to some variable such as fruit production. Through these procedures, we can peel away the different layers of information that are recorded in the ring widths and analyze more obscure signals in the trees (such as mast events; Speer 2001).
Different species of trees have different responses to some variables and we can use that individual response to examine specific environmental variables. For example, trees are often attacked by defoliating insects that are host specific, meaning that they only affect certain species or genera of tree. By comparing the growth of different tree genera, we can tease apart the effect of a pathogen on a tree.
Aggregation of data at different spatial scales can also be used to average out extraneous responses of individual trees. A climate reconstruction, for example, is conducted at the stand level with at least 20 trees (Fritts 2001). We generally take two cores per tree and the ring width chronologies from these 40 well-dated cores are averaged together to develop a climate reconstruction. By averaging the individual core chronologies together, we compensate for events that affect the growth of one tree or one side of the tree. If the signal of interest occurs on a stand or regional level, individual tree competition for light and nutrients can be offset by calculating their average in order to strengthen the broad scale signal.
Examples of Dendroecological Reconstructions
Anthropogenic Ecology and Resource Availability
Dendroecological records can be used to examine anthropogenic ecology and resource availability for prehistoric through historic time periods. Some examples of the wide range of applications include examination of fire history, tree establishment and death dates, culturally modified trees, climate reconstructions, insect outbreak reconstructions, and mast histories (Table 1). In the following section we describe examples from each one of these applications and suggest how future collaborations can enhance our understanding of the resources that were available for native peoples and how they may have modified their landscapes.
Summary table of the usefulness of dendrochronological applications to archaeology, including advantages and difficulties with the techniques.

Fire in the Southwestern United States
Native American use of fire is an issue that has been debated for the last 50 years (Adams 2004; Agee 1993; Pyne 1982; Swetnam 1990; Vale 2002; Wagner 2003). Native Americans may have used fire to aid in hunting, to improve grasslands, clear land for agriculture, promote growth of particular vegetation, or as a strategy in warfare (Adams 2004; Pyne 1982; Shinn 1978; Stewart 1936). Lightning ignition of fires is also a common phenomenon and produces a natural background of fire occurrence (Swetnam and Baisan 1996). Swetnam and Baisan (1996) argue, however, that appropriate fuel and climatic conditions, not ignition sources (i.e., lightning strikes), are the limiting factor in the occurrence of fire.
Dendrochronologists have developed fire histories to inform us about spatial and temporal patterns of fire on the landscape. Fire scars can indicate the frequency, intensity, and seasonality of fires. For example, examination of the earlywood and latewood of the fire-scarred tree ring can specify what time of year the fire occurred (Swetnam et al. 1999). Fire histories can also be used to examine anthropogenic effects on the fire regime (Adams 2004). These records easily extend before the late 1800s, which was a time of heavy grazing by sheep and cattle and was followed by fire suppression by the Forest Service (Savage and Swetnam 1990). Fire histories can be used to determine the pre-settlement fire regime so that forest managers can use prescribed burning to return the forests to a pre-European condition and improve forest health (Morgan et al. 1994; Swanson et al. 1994; Swetnam et al. 1999). Swetnam and Betancourt (1990) have shown that fire occurrence in the southwest can often be explained by climate patterns, especially at the broad scale. They examined fire histories that extended back to the 1600s from 55 sites throughout the southwestern U.S. (Swetnam et al. 1999) (Figure 1) and found that regional fire occurrence was driven by long-term climate fluctuations such as El Niño. These fire histories show the occurrence of past fire in the southwestern U.S., but the recurrent question is how Native Americans affected the fire regime.

Fire history reconstruction for a network of sites throughout the southwestern United States. Each horizontal line in the top part of the graphic represents an individual site while each tic mark on that line represents a separate fire event. The bottom part of the graph is a composite of fire occurrence throughout the western United States and northern Mexico. These 55 fire histories extend back to AD 1600 (from Swetnam et al. 1999).
Dendroecological work by Wilkinson (1997) extending back to 1652 has shown some effect from Native American burning as demonstrated by an increase in fire occurrence during times of Spanish pressure on Native American encampments. In the Sacramento Mountains of New Mexico, she found that broad-scale disturbance from anthropogenic sources did not occur until the introduction of grazing in the 1880s and fire suppression in the early 1900s. In the local area of Dog Canyon, however, she did observe a short-term change in fire frequency and season of fire associated with Native American use of the area. Deviations in fire frequency or seasonality from the natural regime may indicate anthropogenic use of fire (see Adams 2004). This local effect on the fire regime was identified by comparing fire occurrence over a broad area and in a specific forest type to individual sites histories. This approach has been successfully used in other studies as well (Kaib et al. 1996; Morino 1996; Seklecki et al. 1996), and compares the broad-scale pattern as the background signal to the local pattern to identify irregularities in the fire histories. Knowledge of habitation site locations and the timing of migrations would enhance the dendroecological interpretation of fire histories and help to understand the anthropogenic use of fire.
Fire in the Eastern United States
In the eastern U.S., the role of fire in shaping the vegetation communities is not as clear as in the western U.S., but anthropogenic use of fire is well documented (Wagner 2003). Fire is becoming recognized as a natural and important factor in maintaining oak woodlands (Abrams 1985, 1992, 2000). Some interpretations of dendroecological fire reconstructions, however, argue that Native American and Euro-American burning is responsible for many of the past fires in the Eastern U.S. (Buell et al. 1954; Day 1953; Jenkins et al. 1997; Sutherland 1997). Buell et al. (1954) examined an oak cross-section and ascribed six fires from 1627 to 1711 as being caused by Native American burning. Buell et al. cite Day (1953) as support that fires before Euro-American contact were ignited by Native Americans. Day (1953) discusses Native American modification of the local landscape as reported by early explorers’ writings about Native American behavior as well as observations of their villages. The fire history from Buell et al. (1954) is one of the longer fire records in the eastern United States but it is limited by coming from one individual tree that was not crossdated. Only a few fire histories have been developed in the eastern U.S. and most of those do not extend beyond the 19th century. More regional dendroecological work with a longer time perspective on specific sites of known habitation could provide the means to test hypotheses of Native American burning and help to differentiate between a natural fire regime and one that has been modified by human interactions.
Tree Establishment and Death Dates
Landscape modification by humans is one focus of anthropogenic ecology. Examples of extensive landscape modification for the purpose of prehistoric agriculture can be identified in the American Southwest. Some of these examples include the extensive irrigation systems of the Hohokam (Woodbury 1961), trincheras, or checkdam fields to catch soil and water resulting in a terraced landscape, or rock mulching to retain soil moisture (Adams 2004; Fish 2000; Minnis 2000). Landscape modification due to agricultural practices is also apparent in the Eastern Woodlands as ridged and hilled fields, mounds, and areas of cleared vegetation (Wagner 2003). Not all landscape modifications are as extreme as the above examples; neither are all associated with agriculture. A non-agricultural example of extensive landscape modification is the deforestation associated with the Mississippian period town of Cahokia. Here, wood was harvested not only for fuel, but for construction of houses, public buildings, woodhenges, and palisades. Approximately 15,000-20,000 logs would have been required for each palisade, and an estimated 800,000 posts for all the houses in the town (Lopinot and Woods 1993). Large and small landscape changes can be recorded by trees that lived during or were established after these landscape-altering practices, and composition of the forest can give further insight into broad scale vegetation changes.
Although dendrochronological stand-age structure analysis of forests has not been an area of collaboration between archaeologists and dendrochronologists, this technique can potentially be used to examine how Native American groups may have affected local vegetation patterns. Dendrochronological stand-age structure may be particularly well suited to addressing historic anthropogenic landscape changes. Stand-age structures quantify the current forest composition and recent changes in forest composition through determining the dates of tree establishment and dates of tree mortality (Abrams et al. 1995). All living and dead trees within a plot must be sampled and age dated to provide a complete picture of the establishment and mortality of all tree species. This technique then yields a history of forest dynamics for the lifetime of the trees. In conjunction with a complete inventory of the tree species living on the site, we can extrapolate early site conditions and determine what tree species were encouraged to grow on the site. Stand-age structure analysis has been used to document forest succession on old agricultural fields after they have been abandoned (Golley et al. 1994) and can be used to study successional processes in recovering forests. Forest composition that has been modified by Native American use in the past is likely to propagate that modification through time because early management controls the boundary (or starting) conditions of that forest.
One limitation of this application of dendroecology is the age of the trees still existing on the landscape. In the Eastern Deciduous Forest, wood decomposes quickly on the forest floor limiting the length of record available in this biome. Even given this temporal constraint, it would be a productive line of research to compare stand-age structure results from in and around Native American habitation sites to general forest composition data to examine the continuing affect of past Native American land use on local vegetation patterns over the past several hundred years.
Pollen analysis is another method that can be used to examine vegetation change due to Native American and historical land use (e.g., Dumayne-Peaty 2001; Hall 1985; Wycoff 1977). Davis et al. (2002) is an excellent example of the use of palynology to examine questions of anthropogenic ecology. They documented the conversion of six sites from agricultural fields to wetlands associated with charcoal decline and an increase in woody taxa due to Spanish control of the Sonoran Desert wetlands (Davis et al. 2002). Palynology records vegetation change by the occurrence of pollen in a lake core and can also inform us of climate change that may have forced this vegetation change. Dendrochronology can record vegetation change over a shorter time period but with annual resolution through stand-age structure analysis, resulting in establishment and death dates of trees for as long as the trees are preserved on the site. Dendrochronology can further record climate and fire events on these sites with a high enough temporal accuracy that timing can be assigned to the events, enabling researchers to examine the mechanisms for the change. Although the applications of dendrochronology and palynology have rarely been combined, the partnership between them can potentially be a useful collaboration because dendrochronology can examine a record with annual resolution while palynology can examine the same system with greater time depth. Together these two tools can provide a more complete picture of ecological modification of the landscape aiding in archaeological interpretation of human activities.
Culturally Modified Trees
Culturally modified trees (CMT; Figure 2) provide direct evidence of Native American activity on the landscape from Mexico, across the continental United States, through British Columbia, to the southern coast of Alaska (Lewis 2002; Mobley and Eldridge 2002; Swetnam 1984; Towner et al. 1999; Wilkinson 1997). Peel bark trees are a subset of CMTs and researchers can identify the year and season of Native American occupation in an area from the occurrence of these distinctive scars. Native Americans were thought to peel the bark from these trees in the spring time and eat the inner cambium as a starvation food; dates of these events have been related to social forcing factors of the time (Swetnam 1984). Other hypotheses that have been proposed for why trees were peeled include uses for housing, clothing, medicinal purposes, and wood crafts (Towner et al. 1999). Peeled trees generally occur in clusters of about 20 individuals (personal observation) and the scar left on the trees can be dated to the year of damage and sometimes the season (Swetnam 1984; Wilkinson 1997). Analysis of CMTs has been done mainly due to chance location of these sites rather than based on a hypothetical framework developed to determine the driving factors that caused Native Americans to use these trees. Mobley and Eldridge (1992) report on the rare circumstance of systematic studies of CMTs in the Pacific Northwest, reporting on 967 tree scars with the oldest dating to AD 1467. Mobley and Eldridge's work on CMTs was often conducted in the field from ring counts on freshly cut samples. More accurate dates can be provided if all CMTs were dated by dendrochronologists with well-sanded cross sectional surfaces employing standard crossdating techniques. Collaborative work examining the timing of peel events along with archaeological surveys to determine the proximity of habitation sites and an understanding of the social stressors of the time could enable us to test hypotheses and better understand this phenomenon (Andersson 2005; Eldridge 1997; Mobley and Eldridge 1992).

A peel bark tree or culturally modified tree. Notice the scar is wide and starts at about knee height, extending to just above head height. Axe cuts can be found in the wood at the top and the bottom of the scar, helping to identify this distinctly as a culturally modified tree. This particular tree is a Ponderosa Pine tree that is growing on a site that has not experienced fire for about 100 years, resulting in the large pile of needles and sloughed off bark at the base of the tree. Photograph by James Speer.
Climate
Climate reconstruction from tree rings is probably the most recognized application of dendrochronology. Dendrochronologists reconstruct past temperature and precipitation records that can inform investigators of climate conditions during Native American habitation (Ahlstrom et al. 1995; Dean et al. 1985; Grissino-Mayer 1995; Kaye and Swetnam 1999; Salzer 2000a, 2000b; Stahle et al. 1998; Van West and Dean 2000). Grissino-Mayer (1995), for example, developed a climate reconstruction for El Malpais, New Mexico, that extends back to 100 BC This 2000-year long precipitation record delineates drought and moisture episodes for much of the southwestern U.S. Grissino-Mayer compared this climate reconstruction to major changes in Native American settlement patterns. Stahle et al. (1998) examined Bald Cypress (Taxodium distichum) growth in southern Virginia and found that the Roanoke and Jamestown colonies were established during two of the most severe droughts recorded in their 800-year record. Both of these colonies foundered soon after they were established, which demonstrates communities’ reliance upon the resources provided by a temperate climate.
Most dendroclimatic studies across North America reconstruct precipitation because the majority of sampled trees are located at lower elevations where trees are stressed by lack of moisture. Temperature reconstructions are possible, but they need to be conducted at high latitude or high elevation where the trees are stressed by low temperatures. Salzer and Kipfmueller (2005) created a 2,262-year temperature reconstruction from Rocky Mountain Bristlecone Pine (Pinus aristata) in the San Francisco Peaks, Arizona, at 3,536 m elevation. They found that the post-1976 warm/wet period is unprecedented throughout the entire record. During the Medieval Warm Period and Little Ice Age, Salzer and Kipfmueller (2005) record periods of high variability with major shifts between warm or cold and wet or dry conditions. This type of variability creates an unstable climatic condition that is hard to cope with, making subsistence agriculture unpredictable, and leading to possible social strife and collapse of ancient cultures (Dean et al. 1985). Salzer (2000a) compared the Anasazi settlement patterns in northern Arizona to his 2,262-year long temperature reconstruction. Salzer (2000a) discovered three distinctly cold periods in the 1200s around the time of the Anasazi abandonment of their settlements in northern Arizona, suggesting that this climatic variability may have been a factor in Anasazi relocation.
Dendrochronological records can be used to reconstruct factors affecting resource availability (such as precipitation and temperatures warm enough to grow crops), but correlation to these records should not be used naïvely to explain population dynamics without a model explaining the complex interactions and the mechanisms responsible for such changes (Ahlstrom et al. 1995; Salzer 2000a; Van West and Dean 2000).
Insect Outbreaks
Insects have been used as a food resource by native populations around the world (e.g., Blake and Wagner 1987; Chagnon 1997) and trees record insect outbreaks when the insect damages the tree through defoliation or stem damage (Swetnam et al. 1985). Insect outbreak reconstructions can be obtained by examination of the rings of a tree species that is a host for a specific insect. The insect may eat the leaves of a tree reducing its photosynthetic potential, e.g., Pandora Moth (Coloradia pandora), Spruce Budworm (Choristoneura sp.), Tussock Moth (Orgyia sp.), and Gypsy Moth (Lymantria dispar); can affect the cambium of the tree, e.g., bark beetles (Ips sp., Dendroctonus sp.); or be a root parasite, e.g., periodical cicadas (Magicicada sp.). Outbreak damage as recorded in tree rings is often a cryptic signal that may be confounded by a climate signal such as drought. Therefore, the climate signal must be controlled or subtracted, and the most common way to do this is to use a non-host tree species from a similar site to measure the variability in climate through time. Once climate and other disturbances (such as fire or other insects) are accounted for in the chronology, then insect outbreaks can be inferred into the past. This process requires considerable understanding of the ecosystem that is being studied and the effects of various factors on the host trees.
Speer et al. (2001) developed a record of Pandora Moth outbreaks that extends back 622 years in south-central Oregon (Figure 3). Pandora Moth is a phytophagous insect that defoliates Ponderosa Pine (Pinus ponderosa), Jeffrey Pine (Pinus jeffreyi), and Lodgepole Pine (Pinus contorta) in the western U.S. The Klamath and Piute Indians used the Pandora Moth larvae and pupae as a traditional food source (Blake and Wagner 1987). The Speer et al. (2001) record can now be used to examine Pandora Moth as a food resource for the native populations in south-central Oregon. At the same time, knowledge of how intensively Native Americans used Pandora Moth as a resource could help ecologists understand how humans have acted as predators of these insects in the past, possibly modifying their outbreak dynamics. This knowledge can be gained from identification of habitation sites and population estimates made by archaeologists for those sites. It has been suggested that the Native Americans would move their dwellings to the site where the Pandora Moth outbreak was occurring (Blake and Wagner 1987). Speer et al. (2001) found that on an individual site, recurrence of an outbreak may be as long as 156 years, but regional return intervals were around 20 years. Therefore, Pandora Moth could be a driver for settlement patterns and humans could have controlled Pandora Moth populations in the past. Further work is needed to test such hypotheses.
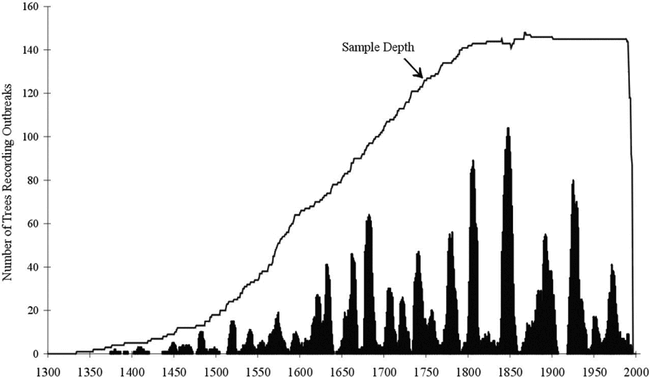
Pandora Moth reconstruction over 622 years from south-central Oregon. The top line represents the number of trees recording outbreaks through time. The sample depth decreases further back in time with 20 trees still recording outbreaks at AD 1500. The dark area shows the number of trees recording outbreaks through time throughout the entire south-central region of Oregon (from Speer et al. 2001).
Mast
Recent work in dendroecology has produced a new technique for developing mast (massive fruit production in trees) reconstructions from tree rings (Speer 2001) (Figure 4). Koenig and Knops (1998) compared previously archived tree-ring chronologies to known records of fruit production in seven genera of coniferous trees (Abies, Larix, Picea, Pinus, Pseudotsuga, Thuja, and Tsuga) and found a slight trade-off between incremental growth and reproductive effort on the landscape scale. To date, Speer (2001) is the only person to have conducted a mast reconstruction using dendrochronology, and these records exist for only five oak species in the southern Appalachian region. Masting is a biological drain on the resources that a tree may apply to incremental growth each year. Climate also controls these resources and is, generally, a more significant limitation on tree growth. To conduct a mast reconstruction, similar to insect outbreak reconstructions, the dendrochronologist needs to control for the climate signal in order to separate its effect from that of masting. This can be achieved with modern climate records or from dendrochronological climate reconstructions. Modern climate records can be obtain for the entire U.S. back to 1895 from the U.S. Historical Climatology Network (USHCN; Karl et al. 1990), but this limits the time depth for the present reconstructions. Another limiting factor is that a set of known mast records needs to be obtained for the species of tree that is being studied so that a statistical model can be developed using regression analysis to determine the response of the trees to masting. Future mast reconstructions will have to be conducted where mast records exist and will need to take advantage of long-term dendroclimatic reconstructions to control for climate.

White Oak regional mast reconstruction from the southern Appalachian Region. The reconstruction is based on 165 White Oak trees from Tennessee, North Carolina, and northern Georgia. Mast years are shown as z-scores with numbers larger than 1.2 and less than −1.2, considered extremely good or poor mast years respectively (from Speer 2001).
Speer's (2001) mast reconstruction used a spectral analysis of White Oak (Quercus alba) mast crop (Figure 5) to identify a distinctive six-year periodicity in the variability of acorn production across the southeastern U.S. (Tennessee, North Carolina, and Georgia) where these trees were sampled. Acorns would have been an important food source for Native Americans and then trickle down resources in the ecosystem would have produced a pulse of regeneration in mice and deer (Ostfeld et al. 1996). Mast reconstruction from dendrochronology could be used to determine the dependability of mast as a human food source in prehistoric times and as livestock feed in historic times.

Spectral analysis of the White Oak regional mast chronology (from Speer 2001) showing the significant cycles in the mast data. Fischer's test of significance is generally used when the system is recording one cyclical pattern while Siegel's test of significance works better in a system that records multiple periodicities (Walden 1992).
Native Americans have long used nuts as a food source, probably from as early as Paleo-Indian times through historic periods. Acorns were a staple of the prehistoric inhabitants of California for thousands of years (Jones 1996). Along with hickory, pecan, and walnut, acorns were an important part of the diet of Archaic cultures (approximately 8000-1000 BC) in the Eastern Woodlands (Asch et al. 1972; Gremillion 1997). Paleoethnobotanical evidence from Cloudsplitter Rockshelter and Cold Oak Shelter, located along the western escarpment of the Cumberland Plateau in eastern Kentucky, showed that acorns were a common food source during the early and late Holocene (9,000 – 8,000 years BP and 3,000 – 2,000 years BP). Nutshells from acorns, hickories, black walnut, butternut, and chestnut are the most common archaeobotanical remains found at these sites during these two periods, suggesting their importance as a food source (Delcourt et al. 1998). The late archaic site of Lamoka in New York State has evidence of intensive acorn use in the form of acorn-roasting pits that are 15 m long (Ritchie 1969). Acorns, hickories, and pecans continued to be used throughout the Eastern Woodlands even into Mississippian times (approximately AD 1000–1500) although with less intensity than domestic grain crops such as maize. Smith (1929) stated that the human race may have eaten more acorns than it has wheat over the span of human evolution. This may be an overstatement, but acorns have definitely been intensively used, and were an important part of the diets of many prehistoric people prior to the domestication of cereals such as wheat and maize. The nutritional content of acorns and the timing of acorn production just before the lean winter months made them an important food source for Eastern Woodlands cultures (Gardner 1997).
Conclusions
The paucity of long-term environmental reconstructions necessitates a central archive where these reconstructions can be stored and indexed for easy access by archaeologists. Currently the International Tree Ring Data Bank (ITRDB) stores tree-ring chronologies from all over the world, although the vast majority of these chronologies were produced for climate reconstruction purposes. Recently the International Multiproxy Paleofire Database has also been established to archive fire history data. Both of these datasets are operated by the National Oceanic and Atmospheric Administration (NOAA) World Data Center (WDC) and free data can be downloaded from their websites. Hopefully, in the future, we can add databases on insect outbreak histories, mast reconstructions, and peel bark dates to facilitate collaboration among scientific disciplines. Whether or not a new database for dendroecological data is developed, joint research between archaeologists and dendrochronologists should be pursued to develop new hypotheses and to further both fields of study.
Archaeology initially benefited from dating techniques of dendrochronology and later from the climate reconstructions developed through analysis of tree rings. Dendroecology has benefited from a better understanding of Native American occupation patterns and the timing of social issues between differing tribes and with European settlers. Both dendroecologists and archaeologists may be aided in the future by working together to develop records of anthropogenic ecology and resource availability. An understanding of the past interactions of humans and the environment can generate new hypotheses of landscape change, ecological dynamics, and cultural change helping us understand humans’ niche in the world ecosystem.
Footnotes
Acknowledgments
We would like to acknowledge our funding sources of the U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, Cooperative Agreement Number PNW 95-0748, NSF STAR grant, and the National Science Foundation Doctoral Dissertation Improvement grant (BCS 0000281). Thanks also to one anonymous reviewer and Karen R. Adams and Rex K. Adams for very helpful comments that have improved this paper.