
Abstract
Variation in folk knowledge of biological kinds among members of a single ethno-linguistic community has long been a focus of ethnobiological attention and interest. Concentrating on variable local understandings of animal names, this paper discusses three forms of binary names applied to animal categories among the Nage of Flores Island, eastern Indonesia. It is shown that while some people, the “lumpers,” regard such names as unproductive binomials designating terminal taxa (folk-generics or folk-specifics), others, the “splitters,” construe the same names as composite terms comprising separate names of two distinct subtaxa. In two of the three cases evidence from the animal nomenclatures of related languages supports the splitters. Also discussed are factors possibly accounting for this variation, including age, gender, and variable familiarity with local fauna bound up with ecological change.
Keywords
In the study of ethnobiological classification it has long been recognized that ethno-linguistically defined groups can vary internally in regard to knowledge of local flora and fauna. Variation can exist, for example, in how informants label specimens as instances of local categories (see e.g., Bulmer and Tyler 1968; Ellen 1993a) or in the results of free-listing tasks, in regard to how many instances of a life-form people can name (see e.g., Brewer 1995; Nolan 2001). Other possibilities concern differences among informant responses to various cognitive tests involving folk taxa (Atran and Medin 2008; Coley et al. 1999). Factors proposed to account for such variation within single ethno-linguistically defined populations have predominantly included informant age and gender (see e.g., Berlin 1992; Berlin et al. 1981; Ellen 1993a), whereas other studies—mostly involving American subjects—have focused on significant differences between informants distinguishable as “experts” and “novices” (e.g., Boster and Johnson 1989; Nolan 2001).
Although bearing on the same general topic, the present paper takes a different approach. My interest is in variable interpretations of names applied to certain categories of insects, frogs, and lizards revealed by ethnographic investigations of the folk zoology of the Nage, a people inhabiting the central region of Flores Island in eastern Indonesia. More specifically, I demonstrate how different folk classifiers construe named taxa as either monotypic (and thus terminal) or polytypic categories—or, using the analytical terminology developed by Berlin, whether or not they interpret particular “folk generics“ as comprising “folk specifics“ and in one case a further division into two “folk varietals.“ As will become clear, in all instances variable interpretation of the same names is facilitated by syntactic features of Nage biological nomenclature. Employing nonrandom stratified samples that included both male and female and older and younger informants, I consider how age and gender may affect interpretations of animal categories. At the same time, evidence from related languages indicates that Nage splitters’ interpretations of particular frog and insect names—those positing taxonomic contrasts within categories—were more common in the past; thus, I further consider factors that would account for both apparent changes and current intracultural variation.
The Cultural and Linguistic Context of Nage Classification
My specific subjects are a community I have previously distinguished as central Nage (e.g., Forth 2011). At present, central Nage comprises six modern administrative districts (desa) with a combined population of 11,652 (August 2014); formerly, the territory covered by the six desa included just three, popularly known as the “three Nage desa,“ referring in part to the inclusion of “Nage“ in each of their names. Nage are speakers of a Central-Malayo-Polynesian language. Their language is most closely related to those of the Ngadha, located to the west, and the Lio and Endenese peoples, residing to the east. Together, these compose a group labeled “Ngadha-Lio“ (Verheijen 1990), and this group, together with the languages of Manggarai (spoken in the western third of Flores Island) and the neighboring island of Sumba, form a complex of languages within Esser's (1938) “Bima-Sumba group.“ Blust (2008) has questioned the validity of Bima-Sumba, particularly in view of a lack of evidence linking Bimanese (western half of Sumbawa island) with other languages in the notional group. But he affirms the connection between Sumbanese languages and those of west and central Flores, thus both Manggarai and the Ngadha-Lio languages. I mention these related languages as the partly historical interpretation which follows will draw—crucially in some instances—on cognate terms contained in their ethnobiological terminologies.
It should go without saying that the nomenclature through which any ethnobiological classification is largely expressed consists of a series of names for individual folk taxa consisting of one or two or more lexemes. In determining lexical units (or “word boundaries”), it is important to note that, in Nage, single lexemes of all kinds are always either monosyllabic (e.g., wa, ‘wind’) or bisyllabic (e.g., wawi, ‘pig’), with the stress in bisyllabic words falling on the first syllable. Accordingly, foreign words of three syllables adopted into Nage are treated as two words; thus Indonesian patola (‘decorated silk textile’), where the stress falls on the second syllable, is replicated in Nage as pa tola, with the stress placed equally on pa (treated like a separate word) and the first syllable of tola. Unlike English, for example, stress does not vary in Nage bisyllables in a way that affects sense (see for example the different pronunciations of ‘insult’ when used as a noun or a verb). Like other members of the Ngadha-Lio group, Nage is also a vocalic language, so all syllables must end in a vowel.
Nage Animal Taxonomy
In the study of folk biological classification, component categories can be fully understood only in relation to formal features of the entire taxonomy. As shown in several previous discussions of their taxonomy of animals, Nage possess a unique beginner, ana wa (‘animal, creature’, Forth 2004) as well as terms for the life-forms ‘snake’ (nipa), ‘fish’ (ika), and ‘bird’ (ana wa ta'a co, ‘animals that fly’). Nage discourse further reveals covert life-forms comprising ‘mammals’ and ‘lizards’ respectively (Forth 2011). By contrast, folk-generics (sensu Berlin 1992) pertaining to other vertebrates and all invertebrates are unaffiliated, that is, they are not incorporated in any named or unnamed life-form taxa and so fall directly under ana wa (‘animal’). In addition to generics, Nage name a number of folk-specifics, and with few exceptions do so by adding a modifier to the generic name—as in English ‘House sparrow’ (Passer domesticus Linnaeus) in relation to the folk-generic ‘sparrow’.
Any folk taxonomy is founded on a principle of inclusion, whereby taxa at lower levels are represented as instances of taxa at higher levels. As discussed elsewhere (Forth 1995), Nage have several ways of expressing class inclusion. The most common is bhia ko'o, ‘kind of’ (bhia, ‘form, manner’), even though, much like English “kind of,“ this can express resemblance as well as inclusion (see “X is a kind of Y,“ meaning that X resembles Y rather than strictly constituting a subclass of Y). On the other hand, Nage possess no nominal form corresponding to English ‘kind, type, sort’; as a result, ‘three kinds of snakes’ can only be expressed as ‘three (snakes)’, a phrase which, though context can indicate that ‘kinds’ is intended, also and more usually refers to three specimens. Nevertheless, nowadays, when virtually all Nage speakers are bilingual, they regularly employ the national language term jenis (‘kind, sort’; also ‘race, species’) when referring to or enumerating kinds (or subclasses within a class). In addition, in ethnobiological discourse they often use me'a (‘alone’, ‘different’, ‘separate’) to identify two categories (or names) as pertaining to different kinds (or folk taxa), as when one says X me'aY me'a, a frequently heard expression meaning that X and Y are categorically ‘separate’ or ‘independent’ entities, or different from one another. By these means, Nage can be directly questioned as to whether animals belonging to a single named class comprise just one or more than one kind (or subclass), and whether two names denote different kinds, as opposed to being, for example, synonymous names for a single kind.
Three Kinds of Names and Three Examples
In what follows I analyze three instances of binary names with reference to three distinguishable forms of binary naming. All three forms can be called “binomials“ in the usual sense of a name comprising two separate lexemes (words), provided it is understood that I refer to folk names and not Latinate binomials (incorporating the name of a genus and a species). Employing animal names as illustrations, the three kinds of Nage binomials are the following:
Productive binomials—terms corresponding to Conklin's “composite lexemes“ (1962) and Berlin's “primary productive lexemes“ (1992)—incorporate the name of a folk zoological taxon followed by a modifier and refer to a taxon subordinate to the one designated by the first name. These occur mostly as labels for folk-specifics, as in goka leo, denoting one of two kinds of goka (python), specifically the Timor python Python timorensis Peters, whose bright yellow and black skin patterning is described by the modifier leo (also the name of the Black-naped oriole Oriolus chinensis Linnaeus). Such productive names are further applied to folk-generics and folk-intermediates, where modified terms denote instances of the named life-forms nipa (snake) and ika (fish), the only two life-forms Nage denominate with single lexemes. Examples of folk-generics thus named include nipa ‘e'e, Wolf snake Lycodon spp., and ika hepa (literally ‘broom fish’), an unidentified species of freshwater fish. Occurring only in reference to ‘fish’, binomials designating folk-intermediates include ika lowo (‘river fish’) and ika mesi (‘sea fish’). In all instances, these terms function as true names and are therefore not to be confused with descriptive usages of identical form (cf. English ‘blackbird’ contrasting with ‘black bird’).
Unproductive binomials are also numerous among Nage animal names. These correspond to what Berlin (1992) designates as “primary analyzable, unproductive lexemes,“ and before him, Conklin's (1962) “unitary complex“ lexemes. Also called “pseudo-binomials“ (Hunn and Brown 2011), they are “unproductive“ because the modified term does not denote a superordinate taxon, and indeed often not an animal at all. In Nage, unproductive names apply mostly to folk-generics and usually describe some distinctive physical feature of an animal. One example is bele teka, ‘sharp wing’, denoting the Peregrine falcon Falco peregrinus Tunstall; another is éko teko, ‘ascending tail’, referring to scorpions. With some names of this sort, the description is metaphorical, as in lako lizu, ‘sky dog’, denoting a Night heron Nycticorax sp., and lema la, ‘protruding tongue’, the name for slugs. Yet another variation is illustrated by kua méze (‘big kua’), denoting species of eagles, which is discussed later on.
What can be distinguished as binary composites are conventionalized combinations of two animal names—typically the names of folk-generics or at any rate taxa of the same rank—which designate a pair of animals. In this sort of usage the names are conjoined in a way implying the English conjunction ‘and’. One example discussed below is pegi ngozo, a general term for ‘cockroach’, comprising what most Nage regard as the names of two different kinds of cockroaches. Another is bhada ja, ‘buffalo [and] horse(s)’, a phrase referring to the largest and most valued of livestock. In fact, the second example is more exemplary of this sort of construction as by far the majority of binary composites do not denote a folk taxon (as does pegi ngozo, cockroach) but special-purpose utilitarian categories; thus bhada ja is employed mostly in reference to bridewealth and other goods exchanged between affines. Despite the necessary insertion of “and“ to produce intelligible English glosses, it is important to note that bhada ja, for example, is not identical to bhada ne'e ja, ‘buffalo and horse(s)’ (where ne'e is the explicit conjunction). For the second phrase refers to a specific buffalo (or buffaloes) associated with a specific horse (or number of horses), whereas bhada ja expresses a collective sense that does not allow counting or enumeration. Mutatis mutandis, the same applies to pegi ngozo and pegi ne'e ngozo, although the first term, as this does name a taxon (‘cockroach’), is indeed numerable.
Occurring in other Malayo-Polynesian languages, and seemingly reflecting a feature widespread in this language group (cf. Ellen 1993a:56-58), binary usages exemplified by bhada ja are especially common in Nage. While prominent in discourse concerning animals and plants (see bu bea, ‘casuarina tree’ Casuarina spp., a taxonomic name understood as comprising two separately named sub-taxa, bu and bea), they are by no means restricted to biological kinds. Illustrating nonbiological usages are ine ame, ‘mother [and] father’, ‘parent(s)’; ulu éko, ‘head [and] tail’, denoting a settlement or region conceived as a territorial and socio-political unity; and uta tua, ‘vegetables [and] palm wine’, the most basic kinds of food and drink. In what follows I deal exclusively with binary names used to label folk zoological taxa: the aforementioned pegi ngozo, ‘cockroach’; tu gea, a frog taxon mostly identified with the Brown bullfrog Kaloula baleata Müller; and mapa bonga, ‘skink’ (Scincidae). Some Nage interpret one or more of the three terms as unproductive names labeling monotypic folk-generics (pegi ngozo, mapa bonga) or folk-specifics (tu gea); however, each is alternatively interpreted by other Nage speakers as a binary composite conjoining two contrasting animal names, thus designating a polytypic taxon comprising two subtaxa.
Before discussing each case in detail, attention must be given to field methods. Basic understanding of Nage knowledge and classification of frogs, roaches, and lizards (skinks) derives from general ethnography conducted over a period of thirty years (1984–2014) while investigating a variety of topics, by no means all ethnobiological. In all instances, general ethnography was complemented by directive and semi-directive questioning, including free-listing—for example, asking informants to list all kinds of lizards (see Forth 2011) they knew. After I became aware that people differed in whether they understood pegi ngozo (‘cockroach’) or tu gea (‘kind of frog’) as labeling a single, undivided (or monotypic) category of animal or a divided (polytypic) category incorporating more than one named kind, I began questioning two sets of informants to discover the extent of this variation. Although each included males and females and people of varying ages, the samples were for a large part “opportunistic,” comprising people who were available at the time and were willing to assist, and in some instances people who had been recommended by others. Although I tried to ensure an equal representation of men and women, for various reasons men proved more accessible and ready to participate than women, so whereas equivalence was obtained in the sample of cockroach informants (18 men and 18 women), in the sample for frogs there were 50% more men (24, as opposed to 16 women). Differences between the ‘roach’ and ‘frog’ samples in regard to gender distribution are incidental, reflecting the fact that questioning concerning the two sorts of creatures mostly took place at different times, when different people were available. Difficulty in accessing especially older female informants explains why, in both samples, women were on average younger than male informants. Nevertheless, the information is sufficient to reveal differences in responses among people distinguished either by sex or age, and thus offers ways of accounting for variable local interpretations of named categories as either monotypic or polytypic—or in other words factors distinguishing what may be called “lumpers“ and “splitters“ (cf. Berlin 1992:225).
In enquiring both about pegi ngozo (cockroach) and tu gea (kind of frog), informants were asked whether there was more than one kind of each and, if so, how they distinguished these, both nominally and with respect to physical appearance and behavior. In addition, people were asked to identify photographs of specimens, a procedure which also assisted in matching Nage categories with international scientific taxa. Investigating different interpretations of mapa bonga (skink) required less detailed questioning than did the other two categories, for it quickly became clear that an understanding of this as a polytypic category was restricted to a very small number of Nage men.
Pegi Ngozo, ‘Cockroach’
I first encountered pegi ngozo as a common Nage translation of Indonesian (Bahasa Indonesia) kacoa (‘cockroach’), subsequently observing it in use as a general term for cockroaches. However, as I later discovered, many Nage interpret pegi and ngozo as separately denoting two different kinds of cockroaches. Of a total of 36 informants (18 male and 18 female) questioned directly on the matter, 24 (14 men and ten women) characterized pegi ngozo as a category comprising two similar yet distinct kinds of creatures. One who did not—a man of 50 who has assisted me in both general ethnographic and ethnozoological enquiries for over ten years—explained pegi ngozo as constructed of two synonyms, and the entire expression as simply “a more complete name.“ Coming from a mature informant and one relatively knowledgeable of local folk zoology, this response shows that a representation of pegi ngozo as a monotypic category is a definite view among Nage, not just a use of a more general term where specific terms are equally known to apply. Nevertheless, it is obviously a minority view.
The majority of people who construed pegi ngozo as subsuming two kinds of cockroaches identified these respectively as pegi and ngozo. Three men and one woman first specified the term contrasting to pegi as pegi ngozo, thereby implying a polysemous use of pegi ngozo as the name for both the superordinate and a subordinate category. As the name of the subordinate taxon, pegi ngozo is interpretable as a productive binomial, with ngozo modifying pegi. Consistent with this is further evidence for pegi being the more prototypical of the two categories, as well as the fact that all four splitters recognized pegi ngozo, in the more specific sense, as equivalent to ngozo.
A large majority of splitters describe pegi as a smaller kind of cockroach, and ngozo (or pegi ngozo where this was used in contrast to pegi) as a larger kind. Indeed, size is the main difference Nage articulate between the two kinds, expressing this with the frequently heard formula pegi so éno, ngozo so méze, ‘pegi are smaller, ngozo are larger’. Differences of color are also regularly remarked. For example, informants describe pegi as lighter (‘white, light’, bha; or ‘dark yellowish, orange’, kune) and ngozo as darker (‘reddish, dark red, brown’, to; or occasionally ‘black, dark’, mite) but showing light-colored markings on the upper back near the head. Consistent with informant descriptions, all evidence, including the results of photo-elicitation (Figures 1–3), indicates species of Blattella (notably the German cockroach B. germanica Linnaeus and the Asian cockroach B. asahinai Mizukubo1) as the usual referents of pegi and the Australian cockroach Periplaneta australasiae Fabricius as the main referent of ngozo. At the same time, seven people further applied ngozo to specimens of the Oriental cockroach Blatta orientalis Linnaeus, another dark-colored roach smaller than the Australian cockroach but considerably larger than local species of Blattella (= pegi). 1
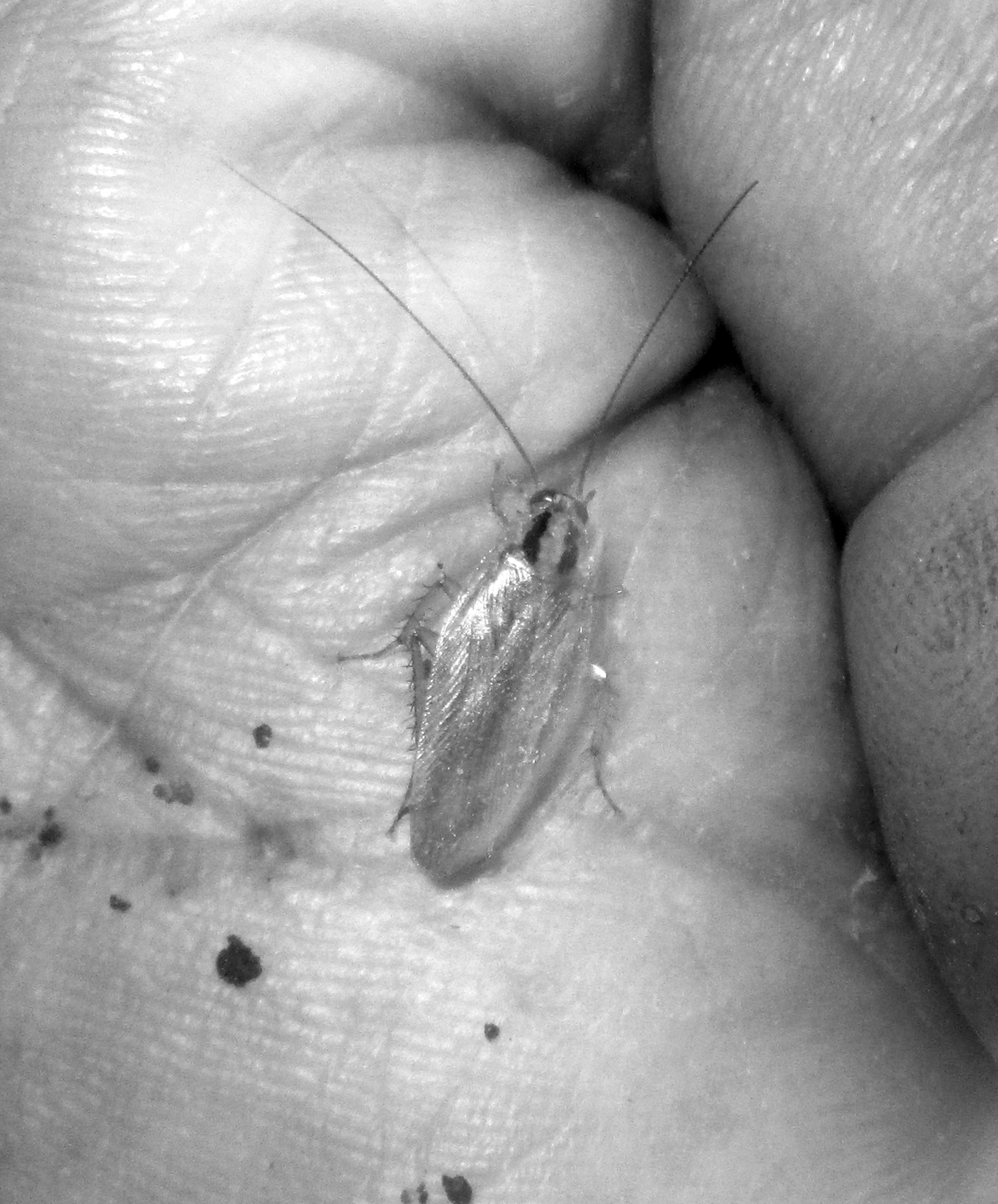
Blattella sp., probably the Asian cockroach B. asahinai, identified by most informants as pegi [1629].

The Australian cockroach Periplaneta australasiae, identified by most informants as ngozo [1655].

Two specimens of the Oriental cockroach Blatta orientalis, identified by most informants as ngozo [1631].
While proportionally more women than men recognized just one kind of cockroach, four of ten women who distinguished two kinds reversed the names, applying ngozo to the smaller and pegi to the larger, and their photo identifications were consistent with this reversal. Other distinctions splitters recognized between pegi and ngozo concerned habitat, odor, and whether they were winged. Most people described both kinds as winged. Attributions of winglessness are mostly accounted for by the fact that Blattella germanicus, although winged, is incapable of sustained flight (Bell et al. 2007:26). Similarly, the common idea that pegi occur only in human spaces while ngozo occur in both dwellings and forests seems not to be contradicted by international entomology.
The contrast of pegi and ngozo is further attested in metaphorical usages. Pegi ni'o kaka meo, ‘like a cockroach accidentally slamming (ramming) into a spider (or spider's web)’ can describe two people colliding. However, since ni'o (‘to slam, ram into’) is a crude reference to a man's thrusting motion in sexual intercourse, one regular informant explained the phrase as alluding to an illegitimate union between persons of different ranks (where pegi apparently refers more particularly to a lower-ranking man). Connected with cockroaches’ habit of consuming the relatively soft interiors of blackened gourd-shells (kula) used as dishes, ngozo wiwi kula, ‘cockroach on the lip (edge) of a dish’, describes a dull, inarticulate person who remains silent whenever others speak and never contributes to public discussion. Some Nage splitters will allude to this phrase in support of their position; yet oddly enough many attribute feeding on gourd vessels mainly or exclusively to pegi—a curiosity I can only explain by species of Blattella (pegi) being more commonly encountered in Nage dwellings, where gourd-shell vessels are typically stored. One male lumper suggested ngozo may be used alone in the metaphor simply as a “better sounding“ abbreviation of pegi ngozo. Yet none of this affects the majority opinion that ngozo and pegi should be understood as, in effect, designating two contrasting folk-specifics subsumed in the folk-generic labeled with the composite name pegi ngozo.
Further support for the splitters’ interpretation is found in comparative linguistic evidence. The occurrence of two names for cockroaches, in some instances reportedly distinguishing different entomological species, appears to be widespread in the languages of Flores and an even wider area of eastern Indonesia. In addition, many of the names are evidently cognates of Nage pegi (Table 1).
Eastern Indonesian terms for cockroaches.
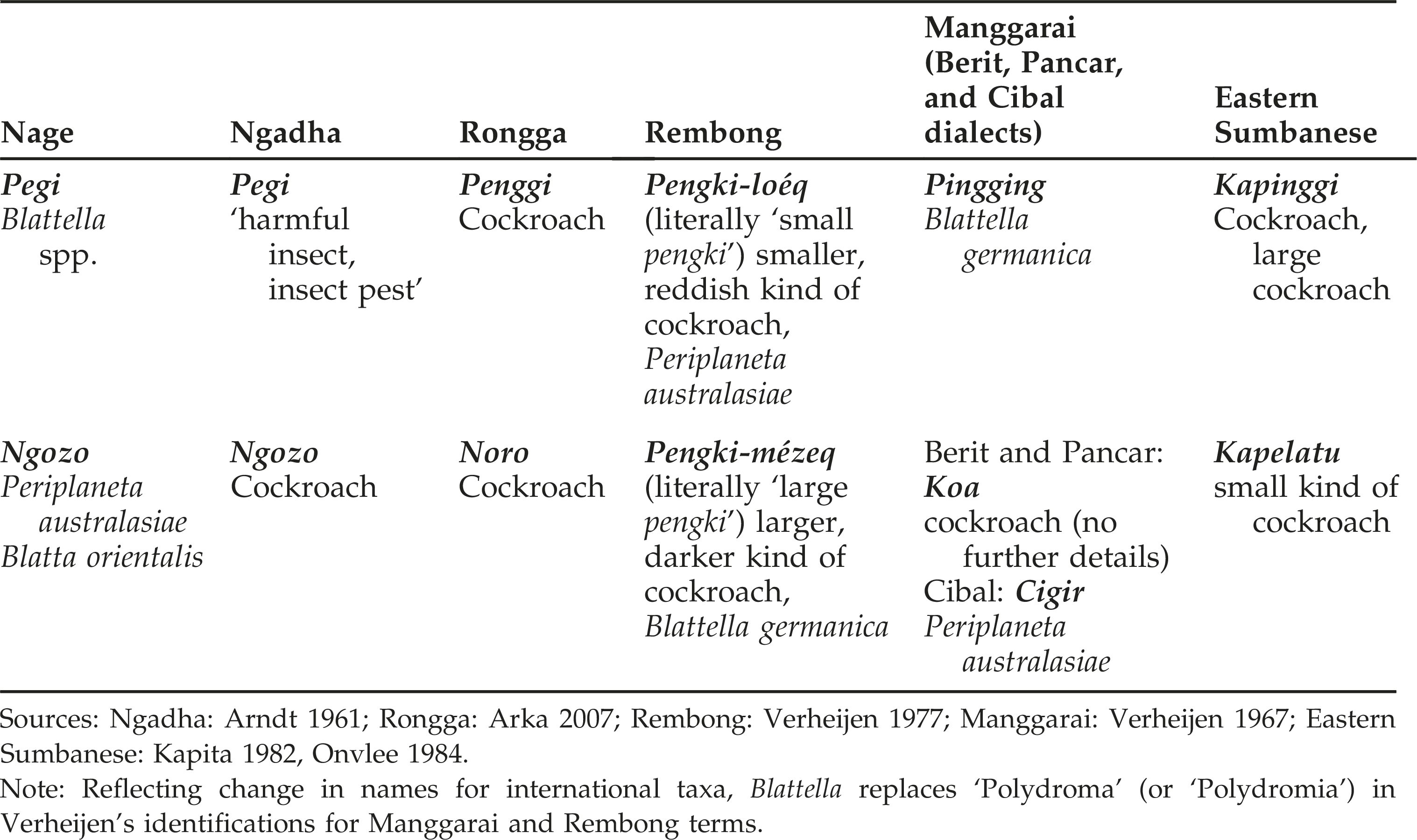
In addition to the evidence given in Table 1, the regional tendency to distinguish two kinds of cockroaches finds further support in the report of a Ngadha woman who stated that, in her native dialect of Mangu Léwa, ngozo méze (‘large ngozo’) and ngozo kedi (‘small ngozo’) respectively distinguish a larger and a smaller kind of roach (Forth fieldnotes 2014). How far this might coincide with the entomological distinctions reported for other languages I do not know. Yet the informant described the smaller kind as occurring inside houses and the larger as mainly infesting modern outhouses and toilets, and the Mangu Léwa contrast of ‘big’ and ‘small’ subcategories is semantically identical to that found in Rembong (Table 1). Questions put to two Ngadha informants failed to illuminate Arndt's gloss of Ngadha pegi; one was unfamiliar with pegi as an insect name, and the other's response was ambiguous. Like Nage, the languages of Ngadha and Rongga—both spoken in more westerly parts of Flores Island—belong to the previously-mentioned ‘Ngadha-Lio group’ of languages. Pegi is also recorded as the single term for cockroach in Lio and Endenese, two other members of this group, spoken to the east of Nage, and for Lio at least recent enquiries conducted by the author confirm this. In addition, in some dialects of Sikkanese, a member of a distinct ‘Timor area’ group of languages spoken to the east of Lio (Wurm and Hattori 1983), pi'i, ‘cockroach’, suggests yet another cognate of pegi.
For both Manggarai, spoken in the far western part of Flores, and Rembong, a language spoken in the extreme northeastern part of the Manggarai district, entomological identifications provided by Verheijen, an accomplished natural historian as well as a highly productive linguist and lexicographer, are likely to be reliable. Even so, he has evidently reversed the identifications for the two Rembong terms (which explicitly distinguish smaller and larger species of roaches) since Periplaneta australasiae grows up to 4 or 5 cm in length whereas the Blattella species are less than 2 cm. For Blattella germanica, the German cockroach, Verheijen gives the now defunct Polydroma germanica, and I have accordingly replaced this with the former in Table 1.
As noted, both Esser (1938) and Blust (2008) recognize connections between the languages of Sumba Island, located immediately south of Flores, and those of central and western Flores. For eastern Sumbanese, Onvlee (1984) identifies both kapinggi and kapelatu with Periplaneta americana, but this appears to reflect a misinterpretation of Dammerman (1926), and it is furthermore curious that, contrary to the general pattern, Onvlee describes kapinggi as denoting not a small but a large cockroach. Since both Sumbanese terms incorporate the fused prefix ka, ubiquitous in Sumbanese languages, kapinggi is obviously cognate with Manggarai pingging and Nage pegi. Kapelatu is curious insofar as kapelatu hapi (hapi denotes Bos spp.) names a large fly, apparently Tabanus sp., which plagues cattle. But this can probably be understood as an unproductive name, and is in any case somewhat clarified by Kapita (1982:95), himself Sumbanese, who remarks that the fly first appeared on Sumba with the introduction of cattle in 1915.
Contrasting to pegi, cognates of ngozo are confined to languages of the Ngadha-Lio group. Manggarai koa, listed for an unspecified cockroach kind in two dialects, is reminiscent of Indonesian kacoa (cockroach). Verheijen (1967) compares Manggarai pingging to Rotinese hihinggi or hihingi and Sasak belinggi. If these terms are indeed related, the evidence would indicate a derivation of the Flores reflexes from a more inclusive group of languages than Central-Malayo-Polynesian (of which the languages Flores, Sumba, and Roti are all members), since Sasak belongs to the Western-Malayo-Polynesian group. Whatever the case, cognates of Nage pegi appear quite widespread in eastern Indonesia. And more widespread still is the labelling of different kinds of cockroaches with two distinct names. Although neither is reflected in any term listed in Table 1, Blust (2002) has identified two prototerms for these insects in Proto-Austronesian: *qatabang, ‘cockroach sp.’, and *Sipes, ‘cockroach’. Indonesian and Malay similarly include two names for cockroaches, kacoa (or kacoak) and lipas (a reflex of Proto-Austronesian *Sipes), but these appear to apply, without distinction, to cockroaches of all kinds. The Nuaulu of the Moluccas employ two terms for cockroaches (matamai putie and matamai reunosu), evidently marking a contrast of folk-specifics within a single generic, although according to Ellen (1993b) these denote different developmental stages rather than entomological species.
The foregoing evidence all points to pegi ngozo as the product of a combining of two names which, originally, simply denoted two contrasting folk taxa. Conjoining them to produce a composite name has thus entailed the creation of a higher-order category; as a result, the original contrast has become lost for some Nage (12 of 36 respondents), but obviously the process has not gone very far. In this scenario, moreover, ngozo and pegi would initially have represented two folk-generics—and more specifically, unaffiliated generics, as roaches are not subsumed in any recognized life-form. Since the emergence of pegi ngozo as a standard term, the component categories have in effect been transformed into folk-specifics, a development which might suggest that the binary term could alternatively be considered a folk-intermediate. But in the absence of any superordinate life-form this raises a logical problem, and on substantive grounds as well it would be difficult to justify treating pegi ngozo as itself a named life-form (which would then contain just two folk-generics).
If pegi ngozo were not a composite term subsuming two originally separate taxonomic names, one would have to imagine it as the original, unitary designation for cockroaches, with a separate application of the two component lexemes to morphologically distinct entomological species then developing later. But this appears ruled out by the comparable use of cognates of pegi and a contrasting term to distinguish cockroach kinds in several related languages. Even so, it is worth underlining the unitary character of pegi ngozo as the name of a single animal taxon. In this respect the term contrasts with non-taxonomic composites—like bhada ja, ‘buffalo [and] horse(s)’—which, despite the utilitarian or symbolic unity of their components, ultimately refer to pairings of two quite different animals. When speaking of cockroaches, including creatures that are visibly present, Nage will thus often refer simply to pegi ngozo, and as is typical with folk-specifics they—or rather the splitters among them—will specify ngozo or pegi only when there is a particular need to do so (for example, when identifying single specimens). In addition, although one can speak for example of ‘four pegi’ (pegi éko wutu, where éko is the numeral classifier for all animals), the composite term can similarly be employed when enumerating cockroaches, as in pegi ngozo éko wutu.
Especially when pegi and ngozo are considered as contrasting taxa, most evidence shows pegi to be the prototypical category, or the “better” example of the pair. Of 18 informants asked to provide a Nage gloss of Indonesian kacoa (cockroach), eight thus gave pegi while another five said this could be either pegi or pegi ngozo. Just two gave pegi or ngozo (indicating these were equally correct) and three responded with ngozo alone. Not surprisingly, all of the latter were splitters. Nevertheless, those answering pegi were evenly divided between splitters and lumpers (four each), as were those responding with the other two alternatives. Thus splitters accounted for six of 18, or fully one-third, of respondents giving only pegi as the translation of the Indonesian name.
Although consistent with pegi reflecting a more widespread protoform than ngozo, this pattern is curious, as the majority of splitters apply pegi to the smaller of the pair (see Hunn 1999). Still, the apparent prototypical status of this category accords with Nage descriptions of pegi occurring only inside houses, and ngozo both inside and outside, as it would then follow that people encounter pegi more often than ngozo—especially women, who spend considerably more time inside dwellings than do men. As the species which splitters distinguish as pegi and ngozo are both found inside houses, the pattern could also explain why more women than men (44.5 as opposed to 22.3%) are lumpers, the conjoining of two names in a unitary reference then paralleling their spatial coincidence in places more often occupied by women. And it should also not be forgotten that, contrary to the more common view, 40% of female splitters apply pegi to the larger kind of roach. At the same time, other evidence suggests that Nage women generally are more often lumpers than are men, a phenomenon demonstrated elsewhere for the Nage classification of rats and mice (Forth 2012), so the predominance of men among informants interpreting pegi ngozo as incorporating the names of two distinct taxa would also reflect this more general tendency.
The predominance of pegi suggested by its greater association with Indonesian kacoa (cockroach) agrees, of course, with its placement before ngozo in the binary term. By the same token, pegi, as a single name corresponding to the single Indonesian name, might be understood as an abbreviation of pegi ngozo. But this only begs the question of why this order and not ‘ngozo pegi’ (which is never heard). Since Nage regularly explain word order in binary composites with reference to the idea of one “sounding better” than the other, one possibility is that the lexical precedence of pegi reflects prosodic considerations, for pegi, especially by virtue of the initial schwa (/e/), has shorter syllables than does ngozo.
Tu Gea, ‘Kind of Frog’, especially Kaloula baleata
Nage pake, ‘frog’, primarily labels a folk-generic. Nage recognize two kinds of pake: one also called pake and denoting riparian frogs (or pake lowo, ‘river frogs’, as they are sometimes described) and another named tu gea (described accordingly as pake wolo, ‘hill, dry land frogs’). Pake in the more inclusive sense includes only frogs; there are no other amphibians on Flores, and the category does not include non-amphibians—unlike the ‘frog’ taxon of the Kalam of New Guinea, for example, which further subsumes eight folk taxa of small mammals (Bulmer and Tyler 1968). Defined in this way, the contrast of pake and tu gea can be called universal among Nage zoologists. And so can the representation of tu gea as a subclass of folk-generic pake—a relationship evidenced in such statements as ‘tu gea are also (a) pake’ (tu gea pake mogha) and occasional references to the second sort as pake tu gea (although this is not the usual way of naming the category). At the same time, a sizeable minority—11 of a sample of 40 people questioned directly on the matter—additionally interpreted tu and gea as respectively designating another pair of frog taxa subordinate to the first. Here, then, we have another example of some Nage interpreting as a polytypic taxon what others understand as a monotypic taxon, and it will be noticed straightaway that, according to the splitters, tu gea is a combination of two folk taxa of precisely the same form as pegi ngozo (cockroach). The difference, of course, is that, insofar as pake in the most inclusive sense can be considered a folk-generic, the composite term in this instance conjoins folk-varietals rather than folk-specifics.
Actually, Nage frog taxonomy is somewhat more complex than this, as a minority of people recognize an additional kind named pake bago (Figure 4). Nevertheless, it is certainly simpler than the classification of frogs reported by Bulmer and Tyler (1968) for the Kalam of New Guinea, who employ 31 terms to name 22 terminal taxa corresponding to 18 or 19 herpetological species. The difference partly reflects the number of species available for Nage to classify. Monk et al. (1997) list just nine frog species for Flores, and it is unlikely that all nine are represented in the relatively small highland region covered by the present study. If pake (frog) can be understood as an unaffiliated folk-generic subsumed only by the unique beginner category ana wa (animal), then at a lower level Nage further apply pake (pake 2 in Figure 4) to a folk-specific taxon comprising larger frogs encountered in or near streams or other bodies of water. International scientific referents likely include one or both of the two species of Occidozyga which occur on Flores. In contrast, tu gea denotes a frog which Nage generally describe as smaller and thinner than pake, but with a large, distended belly and a loud and distinctive vocalization heard especially after rain and most noticeable after dark. As questioning with photographs affirmed, the name primarily denotes the Brown bullfrog Kaloula baleata, a member of the Mycrophylidae (Narrow-mouthed frogs).
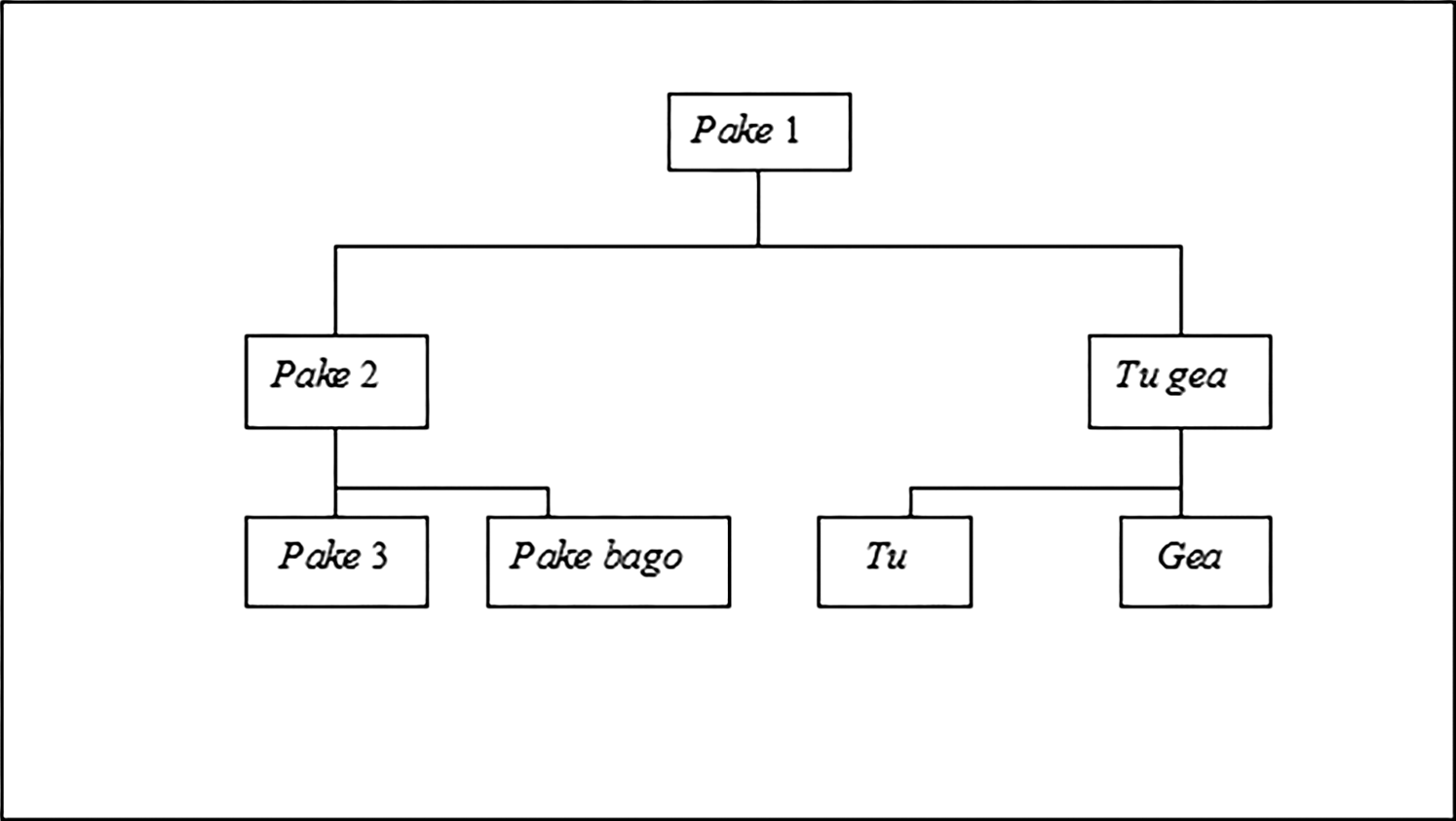
A Nage splitters’ classification of frogs (pake).
Besides the fact that they are typically not found in bodies of water, among the most distinctive features of tu gea for Nage is their nocturnal booming, a circumstance that may account for some people describing frogs named pake, incorrectly, as mute. Yet their generally unitary representation of tu gea raises the question of how splitters distinguish tu from gea. All evidence indicates that tu, an onomatopoeic name reproducing the booming call which largely distinguishes tu gea from pake—and which Nage often imitate as “tung, tung“—refers simply to Kaloula baleata. In other words, tu and tu gea are synonyms. On the other hand, for splitters, gea can separately denote a small tree frog, a kind occasionally referred to as pake gea and most likely the Golden flying frog Polypedates leucomystax Gravenhorst, one of the Old World tree frogs (Rhacophoridae). Some lumpers too are familiar with small tree frogs of the sort splitters designate as gea. However, they describe these as nameless, thus recognizing them only as a covert category and furthermore one that is not subsumed in tu gea.
What then would account for this variation? Both tu and gea have other, non-zoological senses. Tu additionally means ‘to bring, take (along)’, ‘to accompany, lead, guide’, and ‘to hold, maintain’, while gea can mean ‘open’, ‘loose, hanging loosely’. Gea was also given as the name of an (unidentified) plant. Yet there is no indication that tu gea exemplifies an unproductive name, like éko teko, ‘ascending tail’ (scorpion). Another possibility is suggested by Nage zoological names like kua méze (eagle), where méze, ‘big’, modifies unanalyzable kua without altering the referent. Since kua, a term with several other senses (including the name of the large vine Flagellaria indica), is also employed alone to refer to eagles, and since tu and tu gea are similarly synonymous, this suggests that gea may serve a similarly disambiguating function—a possibility I return to below. But this still leaves the question of the derivation of gea, and hence tu gea. One splitter claimed that gea imitates the frog's call, with which it replies to the other creature's tu. Similarly, a lumper suggested that tu and gea may represent different vocalizations of males and females of Kaloula baleata. However, both statements seemed speculative, and once again comparative linguistic evidence provides a far more definitive answer.
More specifically, dialects of Manggarai include cognates of tu and gea apparently denoting the same frog species as are labeled by the Nage terms. Comparable to Nage tu, central Manggarai ntung, refers to a ‘large-bellied’ and ‘very noisy’ frog which Verheijen (1967) identifies as Kaloula baleata, the narrow-mouthed frog known for its noctural booming, while in the dialects of Lamba-Léda and Riwu, ngéar, comparable to Nage gea, denotes a ‘tree frog’. Ngéar is also listed as the name of ‘a kind of small frog’, also specified as pake ngéar, in Rembong. In addition, Kaloula baleata is named tung in Rembong, described by Verheijen (1977:v) as a separate language but displaying features of both Manggarai and the Ngadha-Lio group, and the frog is called by the same name in Rongga, a member of the second group.
In Ngadha, thus in the region immediately to the west of Nage, Kaloula baleata is called tu te—apparently a fully onomatopoeic name (cf. Rembong tung-teng, glossed by Verheijen as the sound made by Kaloula baleata), while in Lio, to the east of Nage, a similarly described species is named tu tumba (Forth fieldnotes 2014). In both Lio and Ngadha, therefore, we find tu, as a component of a frog name, combined with different components, thereby further attesting to the independence of tu from gea in Nage tu gea. What Ngadha te and Lio tumba represent in these terms is uncertain; most likely they simply serve a disambiguating function, as I suggested above for tu gea. But with Nage gea, by contrast, terms like Manggarai and Rembong ngéar, denoting a distinct kind of frog, disclose another, more definite explanation. Here it should be recalled that, as Nage and other languages of the Ngadha-Lio group are vocalic, terminal consonants present in apparent cognates in other Flores languages (like Manggarai ntung and ngéar) do not occur—or have been lost insofar as they were likely original to protoforms. In addition, Ngadha-Lio languages contain no sound corresponding to Manggarai /nt/, which is reproduced simply as /t/, while according to the orthography I employ here, /e/ in Nage gea is a long /e/, corresponding to the sound Verheijen writes as /é/ in Manggarai and Rembong ngéar.
Paralleling separate uses of pegi and ngozo in metaphorical expressions, at least one such expression employing tu alone is interpreted by Nage commentators as a reference to the frog most designate as tu gea. This is tu neke ngai, a phrase some Nage interpret as ‘tu (the frog) holds its breath’ and which is used to refer to someone taking a rest, or a ‘breather’. Not everyone I asked was sure that tu in this expression referred to the frog, and indeed the word, especially in its other sense of ‘to hold, maintain’, can be construed differently. Nevertheless, most evidence points to Nage tu gea as a binary composite comprising names which denote, or originally denoted, two distinct and contrasting folk taxa. Insofar as tu, in the usage of splitters, refers to the same kind of frog as is more often designated by tu gea, the first term obviously specifies the prototype of the more inclusive category. Thus tu gea is constructed in essentially the same way as pegi ngozo (cockroach), and the main difference is that whereas most Nage recognize pegi and ngozo as labeling two different kinds of cockroaches, only a minority, albeit a fairly large one (27.5% of a sample of 40 respondents), recognizes gea and tu as naming different kinds of frogs.
From an historical perspective, therefore, it would appear that a process of generalization, whereby a term combining names initially labeling two subtaxa comes to be used to name a monotypic taxon, is noticeably more advanced in the case of tu gea. Details of the sample of 40 informants suggest how the change may be occurring. Of the 11 splitters, eight were men and three were women; in regard to the total male and female informants (24 and 16 respectively) these figures represent 33.33 and 18.75%. Thus it would appear that men are more likely to be splitters than are women. The difference is consistent with the fact that women encounter the amphibians less often than do men. Although frogs generally possess little utilitarian value for Nage—some people will eat riparian frogs (pake) while Kaloula baleata (tu gea, or specifically tu) is consumed to treat breathing disorders—Nage men spend considerably more time than women in forests and other places outside of settlements where frogs chiefly occur, and men are far more engaged in freshwater fishing, where frogs are incidentally caught by methods used to catch fish, eels, and crustaceans.
Differences of age among male and female informants qualify this interpretation, particularly as the female sample was on average younger than the male sample. Whereas 75% of the men were aged 60 or older, for women the figure is just 25%. Also, 25% of women in this age group were splitters, compared to a figure of 38.88% for men; thus the proportion of splitters is increased by fully a third when only older females are considered, but by just 16.56% when this is done for males. (Informants of either gender aged 50 or under were not sufficiently numerous to provide a meaningful comparison; however, of the two men in this category, one was a splitter, whereas all of the six similarly aged women were lumpers.)
The figures thus suggest that age as well as gender influences the extent to which Nage distinguish tu and gea as two different kinds of frogs. And this is important as age is of course the factor more likely to reflect classificatory change, age differences among both men and women suggesting that knowledge of the creatures has generally declined over time. Accompanying this change, and in part probably explaining it, is a recognized decline in frog populations locally attributed to habitat loss, including loss of natural waterways following the construction of a modern dam on the river Ae Sésa about 55 years ago and the introduction of irrigated rice cultivation in the 1930s. Other factors include chemical pollution of water sources (also connected with expanding rice culture) and newer fishing practices—specifically, a deliberate use of chemical agents and electric-shock fishing—which kill not only fish but frogs and other aquatic creatures as well. And linked with an expanding human population, yet another factor is forest clearance for fields and settlements, leading especially to a reduction in non-riparian and arboreal frogs. For these reasons, one would expect a decreasing familiarity with frog morphology and behavior among younger people, both male and female, while older men retain a more detailed knowledge gained in their younger years.
Further supporting this interpretation is variable familiarity with another frog category, pake bago, ‘leaping frog’. Although Nage recognize leaping as a behavior generally characteristic of frogs, a minority employ pake bago as the name of a large arboreal frog, apparently understood as a folk-varietal of pake (or pake 2 in Figure 4). Just six of the 40 informants distinguished pake bago in this way, including four men and two women. All of the four men were over 60, and in fact three were over 70. As one might expect from the generally younger age range of female informants, only one of the two was over 60 (65 to be exact) while the other was 53. Moreover, the younger woman described pake bago in a way more suggestive of the smaller tree frog Polypedates leucomystax, the apparent referent of gea in the usage of Nage splitters.
Nage recognition of three folk-varietals (tu and gea and pake bago) may seem unusual, especially as frogs are both wild animals and of relatively little importance for Nage (see Hunn and Brown 2011:328). Of course, the taxa occur as varietals only in the usage of a minority, and evidently a declining one at that. Nevertheless, the comparative linguistic evidence implies that this relatively elaborate classification was more general in the past. And it further suggests that, in a wider context, pake (‘frog’ in the most inclusive sense) might alternatively be considered not as an unaffiliated folk-generic, as I have treated it here, but as a named life-form—indeed, one coinciding with the herpetological class of Amphibia (since amphibians other than frogs are absent from Flores). In formal respects, this interpretation is not without merit. Yet one could hardly argue that, for Nage, frogs are a sufficiently prominent and distinctive part of local fauna that they compose a life-form comparable to ‘snake’, ‘fish’, or ‘bird’, and it would in any case be a life-form comprising just two folk-generics (pake 2 in Figure 4 and tu gea), both monotypic and terminal in the view of most present-day Nage. What is more, insofar as it is likely to be short-lived, the taxonomy of three ranks illustrated in Figure 4 can be called transitional and even transitory, in the sense that, for most of their history, the taxa labeled tu and gea probably denoted no more than contrasting folk-specifics, which together with pake (pake 2) and pake bago were all immediately superordinate to pake in the more inclusive sense (that is, pake 1), with the composite term tu gea developing relatively recently.
As to how tu and gea came to be conjoined, the combination most likely developed after the “shrinking,“ as it were, of a more complex protoform corresponding more closely to Manggarai ntung, and as a way of disambiguating tu from homonymous terms. In this respect, it is also conceivable that Nage have rarely used tu alone as a name for a frog and that Kaloula baleata has usually been called tu gea, while gea by itself, or perhaps pake gea (see above), has been reserved for small tree frogs. Comparable examples of binary composites where only one of two animals is the contextual referent are not difficult to find. For example, Nage sometimes use kutu bétu (‘porcupine [and] Giant rat’) to refer specifically to porcupines, owing to the homonymous use of kutu (porcupine) for various sorts of lice.
The same interpretation, it should be noted, could hardly apply to the development of pegi ngozo (‘cockroach’), for the minority interpretation of this term as a monotypic, terminal taxa is more readily explained by a declining interest in differentiating the two kinds, somewhat more marked among women than men. Clearly attesting to this change are age differences among informants; thus, of 17 respondents of both sexes 60 or over, 82.35% were splitters. And ecological or economic factors have evidently not played a part in an emerging conception of pegi ngozo as a terminal taxon, as there is no reason to believe local cockroach species are any less abundant than they ever were.
Mapa Bonga, ‘Skink’
The third case of binary naming subject to variable local interpretation is identical to the previous two insofar as it concerns a taxon which some of Nage interpret as comprising the names of two subtaxa. However, people of this view compose an exceedingly small minority. The term is mapa bonga, the name applied to skinks and most often to the Many-lined skink Mabuya multifasciata Kuhl, the commonest species on Flores.
As many as seven skink genera have been recorded for the island (Monk et al. 1997), so there is certainly scope for nominally marking distinctions among the mapa bonga. Yet the claim that mapa denotes one kind of lizard, and bonga another, is suspect on several grounds. To begin with, the interpretation was proffered by just three men inhabiting two adjacent villages, and two of these were questioned together. According to the latter pair, mapa and bonga differ in the shape of their heads, whereas the third said the differences concerned color. Secondly, I never observed a skink being specified spontaneously as either a ‘mapa’ or a ‘bonga.’ Thirdly, and similarly contrasting to what obtains in other cases, the splitters’ view finds no support in related languages, neither from cognates of bonga and mapa nor from different names for skinks. Finally, some evidence suggests mapa bonga originally reflected an unproductive name. Although mapa also occurs in mapa bewa, the name of a bird (possibly the Bare-throated whistler Pachycephala nudigula Hartert) in the more easterly Nage dialect of Wudu, what this component might mean in either context is unclear. By contrast, bonga denotes a bamboo container used as a pig trough—as does puka, in mapa puka, the lizard's name in the dialect of Menge Ruda, spoken to the northwest of central Nage. Nage associate skinks with domestic pigs, not only because they commonly encounter the lizards in pigpens and often in troughs, but also by virtue of the peculiar notion that male skinks can impregnate sows—an idea also encountered on Sumba and Timor as well as other parts of Flores (Forth 2013). That mapa may represent a lexeme originally modified by bonga is further suggested by Ngadha mapa radza which Arndt (1961) translates as ‘winged lizard’, evidently a reference to the Flying lizard Draco volans Linnaeus, 2 and by the same author's gloss of radza in the related Lio language as ‘large bamboo container’ (Arndt 1933). Of course, Flying lizards are not skinks. Yet not only does the Ngadha name denote a lizard of some sort but in this context radza too apparently functions as an adjective, and the Lio cognate has virtually the same referent as the second components of the Nage and Menge Ruda terms for ‘skink’.
The evidence therefore suggests that the minority representation of mapa bonga as a binary name conjoining two subtaxa (folk-specifics) reflects a false etymology involving a reanalysis of its component terms. Most likely, mapa bonga was originally a descriptive term, with bonga either modifying mapa or figuring as the object in a verb phrase. But in the course of time the term's earlier sense, and especially the meaning of mapa, became obscured, thus facilitating its current minority interpretation—an interpretation, possibly formulated quite recently, which likely reflects influence from what appear to be genuine taxonomic composites such as pegi ngozo and tu gea.
Discussion and Conclusions
I have argued that pegi ngozo and tu gea are best explained as representing different stages in a process whereby folk-generics become combined to denote a single generic. In consequence, component categories become reassigned as folk-specifics, but subsequently they come to be decreasingly recognized as distinct taxa at all, a development prospectively hastened by a reduction of folk zoological knowledge owing to factors of social, economic, and environmental change. Variable interpretations of other binary terms suggest a probable starting point in this process. For example, Nage generally regard poi (grasshopper) and cico (cricket) simply as contrasting generics, as once most likely were pegi and ngozo, the two categories of cockroaches. Nevertheless, sometimes they name crickets and grasshoppers together as poi cico, an occasional usage which does not possess any definite taxonomic status. At the same time, a few Nage interpreted poi cico as a productive binomial, designating crickets (cico) as a subcategory of poi (grasshopper)—an interpretation rendered plausible and arguably motivated by the established use of as many as nine binomials to denote grasshopper folk-specifics (e.g., poi fose, a large, thick-bodied grasshopper, and poi waga, denoting locusts, the largest sort of poi). There are, then, two means by which Nage folk classifiers may eventually incorporate crickets (cico) into a single taxon with grasshoppers (poi). Still, in the majority view, poi and cico represent no more than two contrasting categories which, although Nage recognize their physical resemblance, are not related by class inclusion. 3
If the combining of poi and cico suggests an initial development in a process whose later stages are represented by pegi ngozo and tu gea, then the skink generic mapa bonga could be interpreted as an even more advanced stage. However, the lexical evidence points in another direction altogether. For mapa bonga suggests a quite different possibility, albeit one similarly linked with the binary form of many Nage zoological names and the fact that this identical form allows for varying local interpretations. Specifically, the evidently novel interpretation of mapa bonga described above reveals a possible source of new nomenclatural distinctions and taxonomic contrasts, a resource that can be exploited whenever folk zoologists find a need—intellectual, practical, or otherwise—to express distinctions within categories. In other words, it is not impossible that the minority interpretation of what is by all other indications a monotypic term for skinks could “catch on“ and spread among the folk zoological community.
I conclude with an observation concerning the terms “lumpers“ and “splitters.“ I have used these expressions simply to distinguish people who regard particular named taxa as monotypic and terminal and those who do not. However, this differs from the use of the terms in international biology, where “splitters“ denotes taxonomists who for example interpret as separate species what lumpers consider to be at best subspecies. In contrast, the ethnozoological cases discussed here have more to do with disagreement over what names denote. In addition, this disagreement often reflects, at least in part, differing degrees of familiarity with animals denoted by the names (cf. Berlin et al. 1981:106, who “equate [folk zoological] splitting with expertise“), whereas the varying interpretations of lumpers and splitters in international science are presumably equally informed by the same biological evidence. Yet these differences raise a question. To take the example of the frog taxon tu gea: just because the majority of Nage view this as a terminal taxon without further divisions does not mean that none is aware of physical differences among creatures so designated, nor indeed of the contrary interpretation of tu and gea as referring to different kinds of frogs. As suggested by the fact that lumpers in this instance included many older men generally knowledgeable of local fauna, some may well be aware of such differences, but may deem them insufficient for representing the names as labels for different subsumed kinds. In other words, they may treat observable difference within tu gea as no more significant than the variation in size, color, and so on which occurs within all terminal taxa and thus decide, like the lumpers of international biology, “not to split.“ Variation of this sort therefore may not always reflect lack of familiarity with the creatures denoted. On the other hand, whatever the reason, the possibility of varying interpretations of Nage biological names, provided by syntactical features of their language, facilitates the contrasting interpretations of both splitters and lumpers.
Footnotes
1.
For assistance in matching Nage names with international scientific taxa, I am indebted to John Acorn, an entomologist and award-winning science broadcaster with the Department of Renewable Resources at the University of Alberta.
2.
As regards the precise herpetological reference, it is important to note that Arndt's biological glosses are often inexact, apparently reflecting a lack of knowledge of or interest in natural history. In fact, since Flying lizards seem to be called by quite different names in Ngadha dialects, Arndt's mapa radza could with equal probability refer to a skink.
3.
Paralleling poi cico are Nage expressions like metu mule, incorporating the names of two kinds of ants, and emu hale, ‘mosquitoes [and] flies’. Having mostly metaphorical uses, both are non-taxonomic or special-purpose categories and therefore more comparable to bhada ja (‘buffalo [and] horses’, denoting the most valuable large livestock) than pegi ngozo. Nevertheless, both can be seen to possess taxonomic potential in the same way as does poi cico.