
Abstract
Different but complementary foraging adaptations existed in the Payunia volcanic and the Andean highland (or mountain) subregions of southern Mendoza, Argentina during the late Holocene. We employ an evolutionary ecological perspective to study the zooarchaeology of foraging adaptations after 4000 BP. Seasonal exploitation of guanaco (Lama guanicoe Müller) in the mountains appears to have been sustainable, which relates to high productivity of mountain river valley bottoms during the late Holocene. The mountains were a productive, seasonally available resource patch. A greater diversity of fauna in the Payunia subregion resulted in a different foraging strategy with larger diet breadth. Despite mounting evidence that human population growth occurred during the late Holocene, there is little evidence of resource depression of high-ranked resources, such as the guanaco. Application of foraging theory models in this context clarifies that hunting decisions occurred in the context of two factors, strategies informed by traditional ecological knowledge and the context of landscape ecology.
Introduction
In western Argentina during the last 4000 years of the Holocene, hunter-gatherers sustainably utilized a productive environmental gradient along the escarpment of the Andes. In this environment, the guanaco (Lama guanicoe Müller) represents the largest prey animal and would have been the highest-ranked prey species in terms of net caloric returns of hunting. Foragers followed migrating guanaco herds, which were never domesticated in this region, during their seasonal migration from and to the highland river valleys of the Andes. Although there are no detailed studies of net return rates for species that would have been important prey, there were two additional animal taxa that would also have been important prey. The lesser and greater rheas (Rhea sp.) are large terrestrial birds, but they would have been difficult to capture compared to most other species (Giardina 2010). On the other hand, armadillos (family Dasypodidae) are comparatively small in size, but would have been exceptionally easy to capture. Thus, compared to other species available in this environment (other than guanaco), armadillos and rheas represent moderately high-ranked prey.
This study uses optimal foraging theory (OFT) as a conceptual framework for studying use of these animal resources during the late Holocene in southern Mendoza of western Argentina. Hunting and gathering is commonly studied in archaeology using OFT models from evolutionary ecology (e.g., Broughton et al. 2010; Butler and Campbell 2004; Cannon 2003; Gremillion et al. 2014; Munro 2004; Nagaoka 2002a, 2002b) because it provides a powerful explanatory framework for explaining types of prey taxa and resources patches that were exploited during prehistory (cf. Zeder 2012). OFT explores the consequences of the assumption that hunters make optimal decisions within the landscapes they occupy (references in Smith and Winterhalder 1992; Winterhalder and Smith 1981 1 ). Foragers hold fine-grained knowledge about the places they live, understanding the nuanced details of the species they hunt and gather. Such ecological knowledge must change over time as environments change (Winterhalder 1981:17). On archaeological time scales, it can be assumed that traditional ecological knowledge (TEK; sensu Berkes 1993) reflects many generations’ worth of information about environment, handed down through enculturation. Such information would have been used to make choices during foraging bouts, including when to select alternatives to the highest-ranked taxa. Thus, two contingent factors converge to affect short term decisions during foraging bouts (Winterhalder 1981), both of which contextualize this study of late Holocene animal exploitation in western Argentina: 1) TEK held by foragers, which is a source of information handed down through time; and 2) landscape ecology, which may comprise periods of environmental stability or change (Johnson and Hunn 2010).
In zooarchaeological studies that employ OFT, it is assumed that hunters made decisions upon prey encounter, choosing animals so as to maximize net returns while foraging (foraging efficiency) (Nagaoka 2001). Typically, zooarchaeologists analyze how these choices were made by prehistoric hunters using proxy measures of prey net returns; for example, prey species are commonly ranked according to body size because larger animals provide a greater amount of calories and are thus beneficial to exploit (Broughton 1994a, 1994b; Nagaoka 2002a, 2002b; Ugan 2005). Net returns have also been conceptualized in terms of prey escape strategies (e.g., speed or mobility), which relate to cost of exploitation. Species with similar body size may provide relatively equivalent gross returns but differ in terms of required effort for capture; thus, other factors (e.g., capture technology) being equal, tortoises are more desirable prey than hares because of a difference in cost (Stiner et al. 1999, 2000; Wolverton 2005).
The patch choice model framed within OFT (MacArthur and Pianka 1966; Winterhalder 1981) provides a useful framework for understanding differences between lowland and highland animal exploitation on a seasonal basis in western Argentina during the late Holocene. A high-return resource patch is one in which time spent in search of high-ranked prey species is minimized. A lower-return patch is one in which search time for high-ranked prey is greater. The lowland Payunia volcanic region and piedmont escarpment of the Andes represent more diverse lower-rank resource patches. In contrast, the highland patch would have offered very few animal species for exploitation, but the high seasonal net primary productivity of upland river valleys attracted and concentrated the migrating guanaco populations each year. On a seasonal basis, the Andean highland patch was high ranked during summers but had to be abandoned during winters by guanacos and humans. Foragers took advantage of these dynamics of landscape ecology during the late Holocene in southern Mendoza.
Related to prey and patch choice models, multiple zooarchaeological studies have shown that optimal decisions at the short-term scale of the foraging bout may not lead to sustainable resource use in the long term. Such studies demonstrate that over long periods, consistent exploitation of large prey animals may lead to population decline, which has been termed resource depression (Broughton 1997; Butler 2000; Nagaoka 2001; Stiner et al. 1999). Whether caused by over-exploitation or a change in prey behavior (Wolverton et al. 2012), resource depression represents a shift in what foragers would have encountered on the landscape. In contrast to these previous studies, prey and patch choice in southern Mendoza appear to have led to sustainable use of animal resources by human foragers during the late Holocene.
We employ the expectations of OFT to study long-term trends in exploitation of small, medium, and large game in two physiographic subregions of southern Mendoza in western Argentina during the late Holocene (Figure 1). Prior to 4000 BP during the mid-Holocene, site density in southern Mendoza was low and there was little exploitation of guanaco in the Andean highlands (mountains), particularly above 2200 masl. After 4000 BP, site density increased, and there was consistent exploitation of guanaco in the highlands; here we are concerned with how animal resource exploitation shifted during the late Holocene (after 4000 BP), a period in which human population appears to have increased (Gil 2006; Neme 2007; Neme and Gil 2008a; Neme and Gil 2008b; Neme et al. 2012).
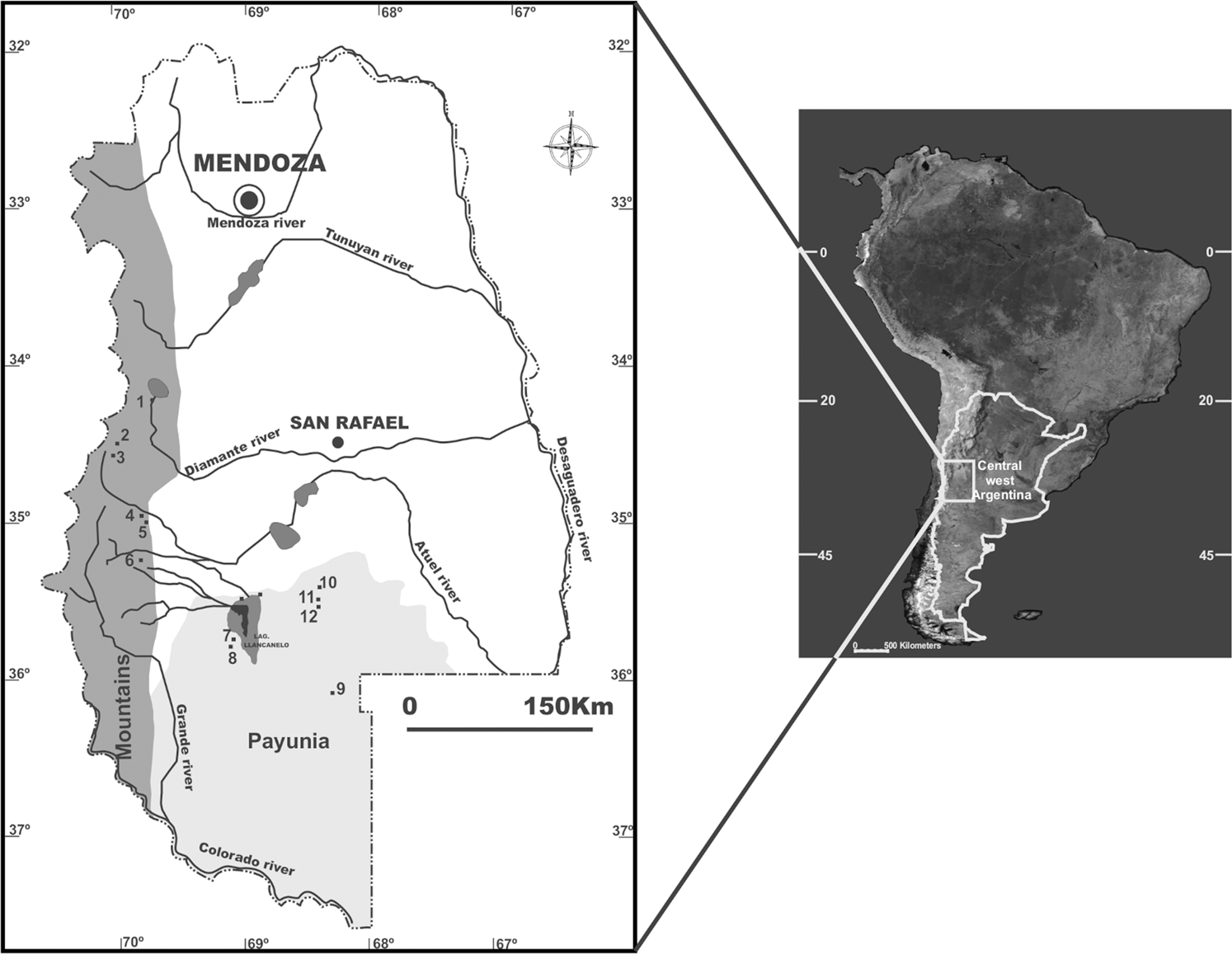
Map of southern Mendoza showing the sites studied in the Payunia and mountain subregions. 1: Laguna el Diamante 4; 2: Los Peuquenes; 3: El IndígenoI; 4: Palulo; 5: Panchino; 6: Cueva Colorado; 7: Llancanelo 29; 8: Llancanelo 50; 9: La Corredera; 10: Los Leones 5; 11: Agua de los Caballos; 12: Puesto Ortubia.
Methods and Materials
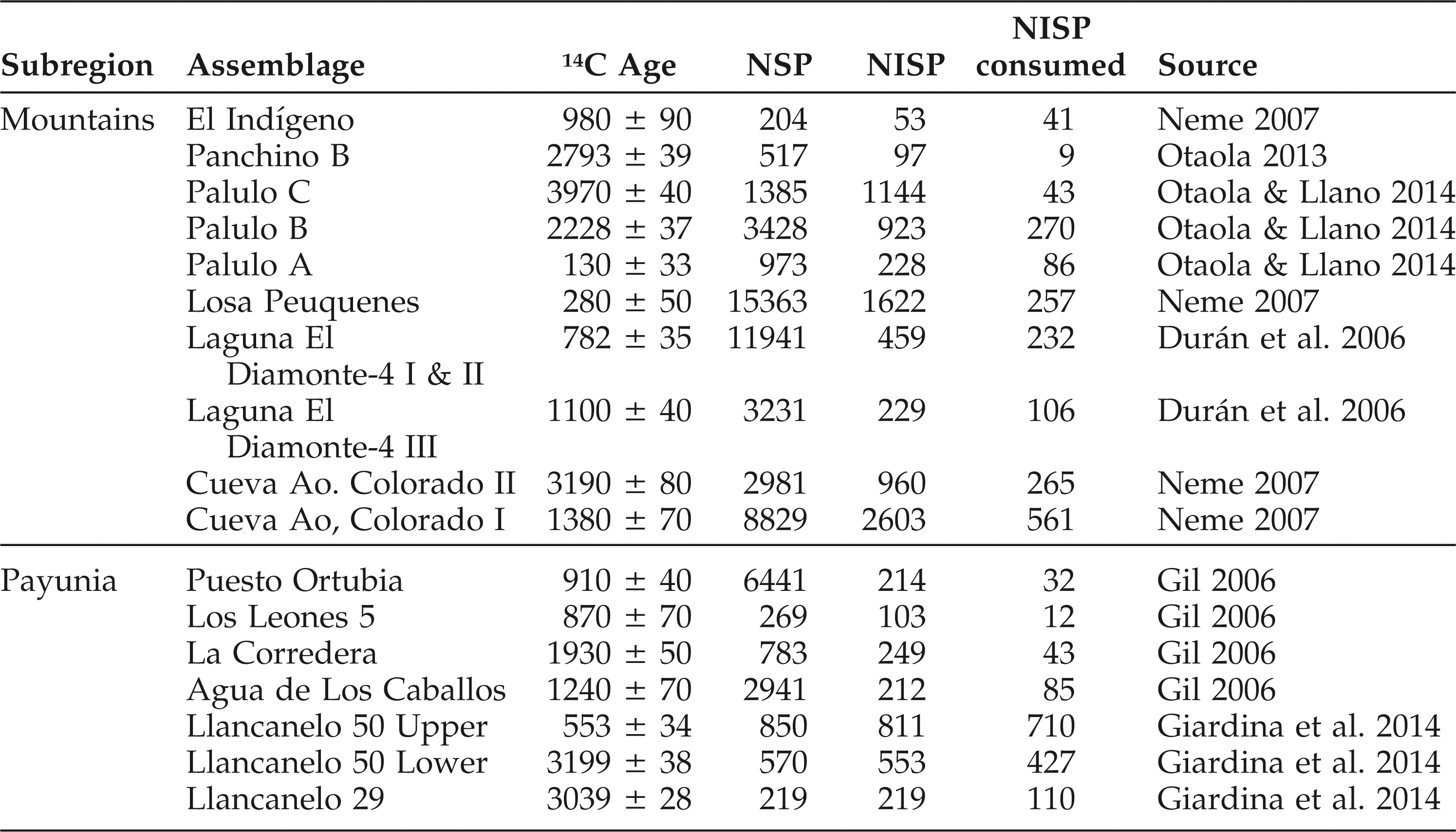
Zooarchaeological faunas analyzed in this study are from habitation sites in the Payunia and mountain subregions of southern Mendoza, Argentina (Figure 1; Table 1). Zooarchaeological remains in this study were analyzed in the Zooarchaeology Laboratory of the Museo de Historía Natural de San Rafael in Mendoza, Argentina following standards reported in Driver (1992, 2011; Wolverton 2013). Remains of taxa thought not to have been part of human diet (e.g., multiple species of small rodent) are not considered, and only faunal assemblages with NISP (number of identified specimens) greater than 50 are included in analyses with the exception of La Peligrosa (dated to 640 ± 70 14C BP; NISP = 47) in the Payunia, which comprises remains of one adult guanaco included in the mortality analysis (see below). Non-parametric descriptive and inferential statistics are employed in analyses following Wolverton et al. (2014).
Summary data for faunal assemblages analyzed in this study.
We used nestedness analysis and visual inspection of species area curves to assess sample representativeness for the early-late (4000 - 2000 BP; abbreviated as ELH) and late-late Holocene (post 2000 BP; abbreviated as LLH) following Lyman (2008; see also Otaola et al. 2015a; Peacock et al. 2012). Nestedness analysis determines whether or not assemblages of different taxonomic richness are nested subsets of one another. If faunas derive from the same animal community, then those with lower richness should nest within the composition of those with higher richness. Nestedness is measured as temperature between 0 and 100 degrees, with 0 representing perfectly nested subsets and 100 representing no nestedness (Ulrich 2008; Ulrich et al. 2009). Visual inspection of species area curves uses similar logic, but here sample size (NISP) is contrasted to taxonomic richness (NTAXA). As NISP increases cumulatively across a set of assemblages from the same animal community, common species should be rapidly encountered and progressively rarer taxa should be added with additional sampling. Eventually a curve plotting NISP against richness should asymptote when additional sampling fails to lead to the encounter of additional taxa. This is termed “sampling to redundancy” (Leonard 1987, 1997; Lepofsky and Lertzman 2005; Lyman 2008; Lyman and Ames 2004); thus, if a curve asymptotes and if faunas are nested, it is likely that faunal assemblages are representative of the prehistoric animal community.
Previous research using nestedness and species area curves indicates that the spatial scale of subregion is appropriate for analyzing late Holocene foraging efficiency in southern Mendoza (Otaola et al. 2015a). There are four physiographic subregions: the piedmont, the desert plain lowlands, the volcanic Payunia, and the Andean highlands (mountains). Zooarchaeological data are sparse in the piedmont and desert plain subregions; our study focuses on the Payunia (lowland) and mountain (highland) subregions. The Payunia of southern Mendoza extends from 1000 to 2000 masl with interspersed volcanic cones that reach up to 3800 masl along the escarpment of the Andes. The subregion has cold winters and dry climate with an average annual rainfall from 200 to 250 mm (Abraham 2000; González Díaz 1972). There are no permanent water sources and precipitation occurs mainly during the winter, with little rainfall between December and April.
Zoogeographically this area is located in Andean Patagonia (Ringuelet 1961). Following Roig's (1972) classification, in the Payunia there are two main faunal groups; mountain fauna and Patagonia steppe fauna. Together, these include guanaco (L. guanicoe), southern viscacha (Lagidium viscacia Molina), Andean fox (Pseudalopex culpaeus Molina), South American gray fox (Pseudalopex griseus Gray), puma (Puma concolor Linnaeus), Patagonian mara (Dolichotis patagonum Zimmermann), hog-nosed skunks (Conepatus sp.), tuco-tuco (Ctenomys sp.), mountain cavy (Microcavia sp.), viscacha rat (Octomys sp.), big hairy armadillo (Chaetophractus villosus Desmarest), dwarf armadillo (Zaedius pichiy Desmarest), turtles (Chelonids), and multiple species of bird, such as lesser rhea (Rhea pennata d'Orbigny), Andean goose (Chloephaga melanoptera Eyton), and many species of duck (Anas sp.). A detailed list can be found in Candia et al. (1993:80-84).
The mountain subregion, ranging from 1700 to 4400 masl in elevation, would have been inhabited seasonally because of subalpine and alpine cold winters. The mountain fauna (Roig 1972) shows a marked reduction in the abundance of small animal species due to seasonally restricted habitats. For example, low abundances of rheids and armadillos, which are common in the Payunia, occur in the lower reaches of the Andean highlands, declining in abundance with elevation up to approximately 2500 masl, becoming absent in the higher reaches of the Andes. Waterfowl, particularly the Andean goose (C. melanoptera) may be seasonally abundant in some parts of the mountains, but some large rodents (e.g., L. viscacia) are absent. Thus, the Payunia represents a region with a more diverse animal community with species that tend to be available for most of the year, and the mountains subregion represents a less diverse fauna with prey species that are available only during the summer months, dominated by seasonally migrant guanaco populations.
Test Implications
Many test implications are used to study the impacts of foraging, including shifts in prey choice, diet breadth, butchery and transport of carcasses, use of within-bone nutrients, and prey demography (references in Broughton and Cannon 2010). Here we use multiple proxy measures to address potential shifts in foraging efficiency over time in the Payunia and mountain subregions of southern Mendoza during the last 4000 years of the Holocene. As discussed earlier in the paper, foraging efficiency can decrease over time under conditions of sustained harvest of a prey taxon, leading to resource depression. Increases in foraging efficiency might relate to resource recovery from shifts in harvest over time or from increases in habitat productivity (environmental change). It is important to consider multiple lines of evidence to assess changes in foraging efficiency.
The number of taxa exploited (NTAXA) is a measure of taxonomic richness that relates to diet breadth (Grayson and Delpech 1998; Nagaoka 2001). If NTAXA increased over time in a subregion, it might indicate a condition of resource depression in high-ranked prey such that alternative, lower-ranked prey taxa were added to the diet. Alternatively, a shift in NTAXA could relate to variability in sample size because discovery probability of rare taxa increases with larger samples (Lyman 2008). Lower-ranked prey might also become higher ranked and added to diet with implementation of new exploitation technology. Diversity, a second proxy measure, assesses the distribution of abundance within the set of exploited taxa; we employ the Shannon Index of Diversity (H) to determine if there is change in the dominance of particular taxa over time. H is lower when richness is low and/or one or a few taxa dominate a distribution; it increases when richness is higher and/or when the distribution of abundance is more even.
The largest-bodied prey species during the late Holocene in southern Mendoza was the guanaco (L. guanicoe), which is considered to have been one of the highest-ranked animal resources (Neme and Gil 2008a). Similarly, armadillos would have been important prey, but for a different reason. It is one of the smallest prey animals that would have been available during the Holocene, but it is also easy to capture, making its net return moderately high (Otaola et al. 2015b). There are two species of armadillo that may be represented, which are difficult to distinguish in archaeofaunal assemblages, the pichi (Z. pichiy) and the big hairy armadillo (C. villosus). Both species would have been more abundant in the lowlands than the highlands.
As high-ranked prey animals, guanaco and armadillos should have been taken upon encounter. In mild contrast, there are two species of Rhea in southern Mendoza, which are also large-bodied compared to most other species. Rheids would have been more costly in terms of search and pursuit time during hunts relative to their size (Giardina 2010), making them lower ranked than armadillo or guanaco, but higher ranked than many other small animals, such as rodents, fish, and small game birds (e.g., Anatids). We construct four proportions to assess relative shifts in each type of prey, proportion of guanaco remains (pGUA), proportion of armadillo remains (pARM), proportion of rheid remains (pRHE), and proportion of all small animal remains (including armadillo) (pSA) in order to assess whether or not changes in foraging efficiency occurred during the late Holocene in the Payunia and mountains.
As a final line of evidence we explore population dynamics of guanaco during the late Holocene. This test implication of foraging efficiency requires the assumption that prey mortality data are representative of the life assemblage of animals on the landscape (Wolverton 2008). Under sustained exploitation pressure, the demographic structure of a population changes. This is termed “steepened survivorship,” and it means that there is a lower probability that a prey animal will survive to progressively older ontogenetic ages. That is, the probability of mortality is consistently high when harvest pressure is elevated, meaning the probability of survival to become older decreases and average life-span shortens, which shifts the population age structure towards higher proportional juvenile abundance. This effect is unlikely to occur without population reduction below carrying capacity due to density-independent mortality (i.e., predation) as the prey population must consistently be held under carrying capacity and is in a perpetual state of pseudo-population growth (Wolverton 2008)—the population is not truly growing, it is simply being held below carrying capacity. If such an effect occurred due to environmental change, carrying capacity would have to perpetually increase over time; instead, when carrying capacity increases, there is population growth for a generation or two in the prey population, which then resumes a stable demographic structure (Caughley 1977; Wolverton 2008).
It is assumed under the OFT framework that hunters would only take juveniles, which provide lower net returns, if larger, older adults are rare in the population. Thus, under conditions of resource depression, smaller, younger guanacos would become more desirable prey, further accentuating the dominance of juvenile remains in archaeological faunas. Zooarchaeologists study this effect with prey mortality data (Broughton 1997; Koike and Ohtaishi 1987; Lyman 1987; Munro 2004), and the signature of harvest that is commonly witnessed is a shift toward relatively juvenile-dominated mortality patterns.
The study of mortality or age structure data in zooarchaeology has been expressed in many ways (Lyman 1987; Stiner 1990), but here it can be quite simple: we assess whether or not the proportional abundance of juvenile guanaco remains changed over time (pJUV). For purposes of this analysis, pJUV is calculated as the proportion of NISP of long bones with unfused epiphyses. Assuming representativeness of the prey life assemblage, if the proportion of NISP of juveniles increased over time, survivorship steepened, which suggests that resource depression related to exploitation occurred. This test implication rests on the ability to generate ontogenetic age data from zooarchaeological samples. With increasingly fine-resolution age distributions in mortality profiles that distinguish multiple ontogenetic age classes, large samples are required (Lyman 1987; Stiner 1990). Our use of pJUV requires only two age classes (juveniles and adults).
Results
Late Holocene faunal assemblages are highly nested (Figure 2A) with low temperatures. Further, visual inspection of species area curves indicates sampling to redundancy for both temporal assemblages (Figure 2B). Related research using nestedness analysis and species area curves by Otaola et al. (2015a) indicates that evaluation of proxy measures of foraging efficiency are meaningful in southern Mendoza faunas at the subregional geographic scale; thus, we focus our attention on the Payunia and mountain subregions.
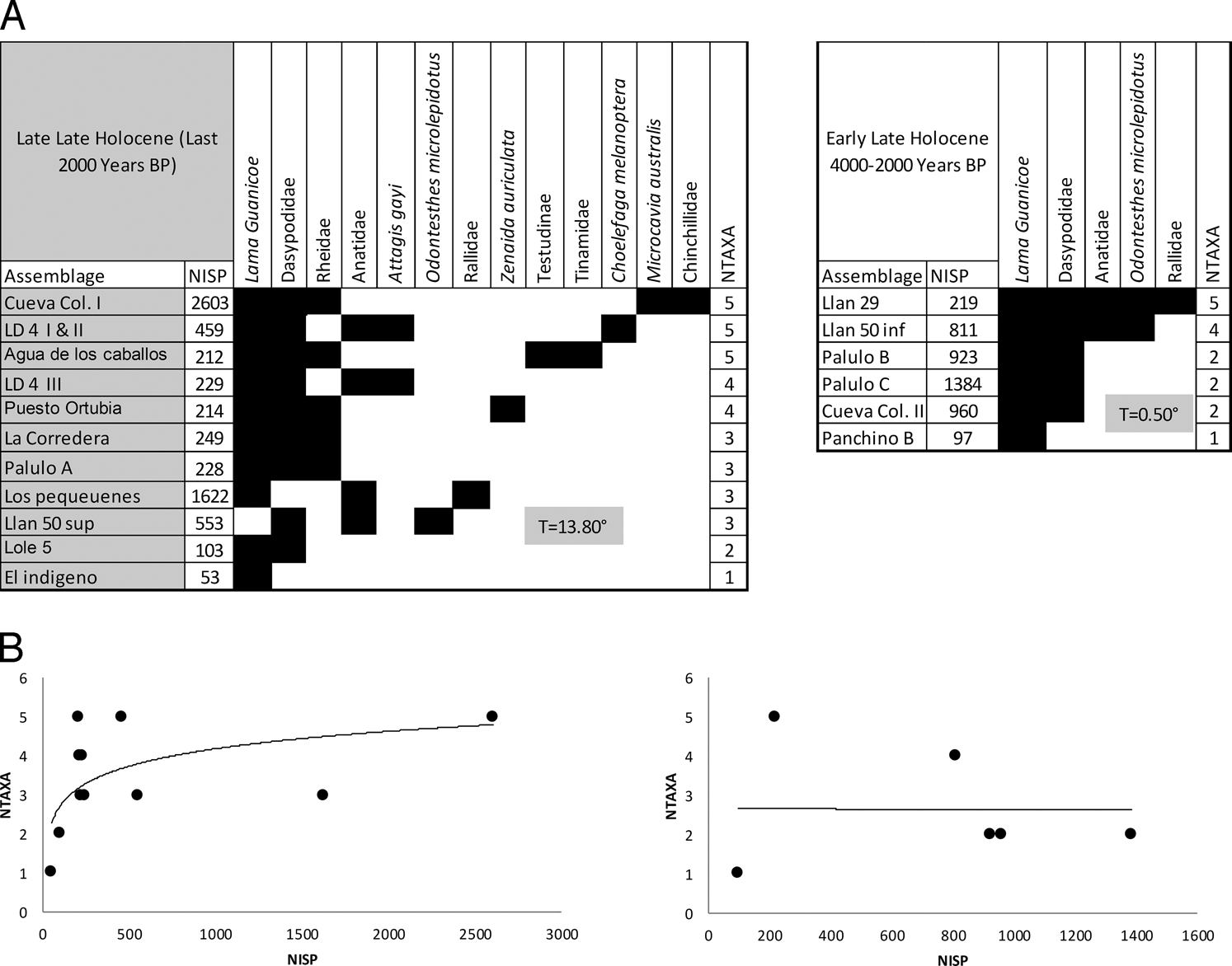
Nestedness (A) and species area curves (B) for faunal assemblages analyzed in this study. Post 2000 BP are the two graphs on the left side, and 4000 to 2000 BP are the graphs on the right side.
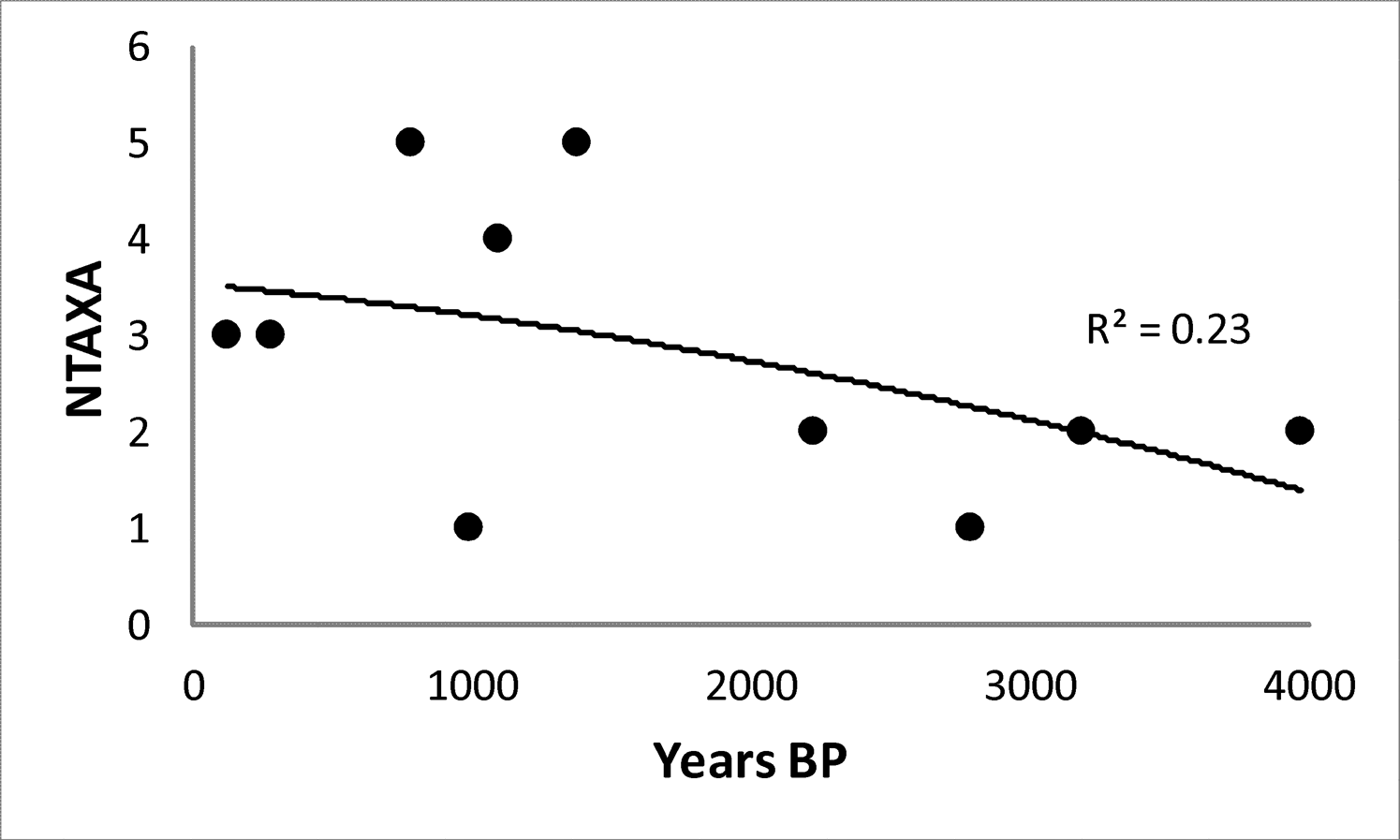
Guanaco remains are consistently dominant in the mountain faunas (Figure 3A; Table 2), and remains of rheids, armadillos, and small animals are uncommon and of low abundance (Figure 3B; Table 2). Median NTAXA over time is 2.5, and there is a slight increase in NTAXA over time (median NTAXA = 2 for the ELH and = 3.5 for the LLH). Although NTAXA correlates to sample size in the mountain region samples (r2 = 0.16; p >0.15), the correlation is slightly stronger between NTAXA and assemblage age (Figure 4; r2 = 0.23; p >0.15). The median of Shannon's Diversity Index (H) is low (0.18), mirroring that guanaco remains dominate the mountain faunas. Guanaco remains are also abundant in the Payunia faunas during the late Holocene, but not as dominant (Figure 5A). Taxonomic richness of the Payunia faunas is higher; median NTAXA over time is 4, and though NTAXA appears to decline from 4.5 to 3 between the ELH and LLH, there are only two sites represented for the ELH. That guanaco is not as dominant is also evident in the much higher median Shannon's H value for the Payunia assemblages at 0.69. Remains of rheids, armadillos, and other small animals (Figures 5A and 5B) represent larger proportions of these faunas. As with the mountain subregion, there is little evidence of change in taxonomic abundance or composition over time in the Payunia faunas during the late Holocene. In general, the mountain faunas appear to indicate specialization on exploitation of guanacos, and the Payunia faunas indicate a diverse diet that guanacos were an important part of. We return to this distinction in the discussion section.
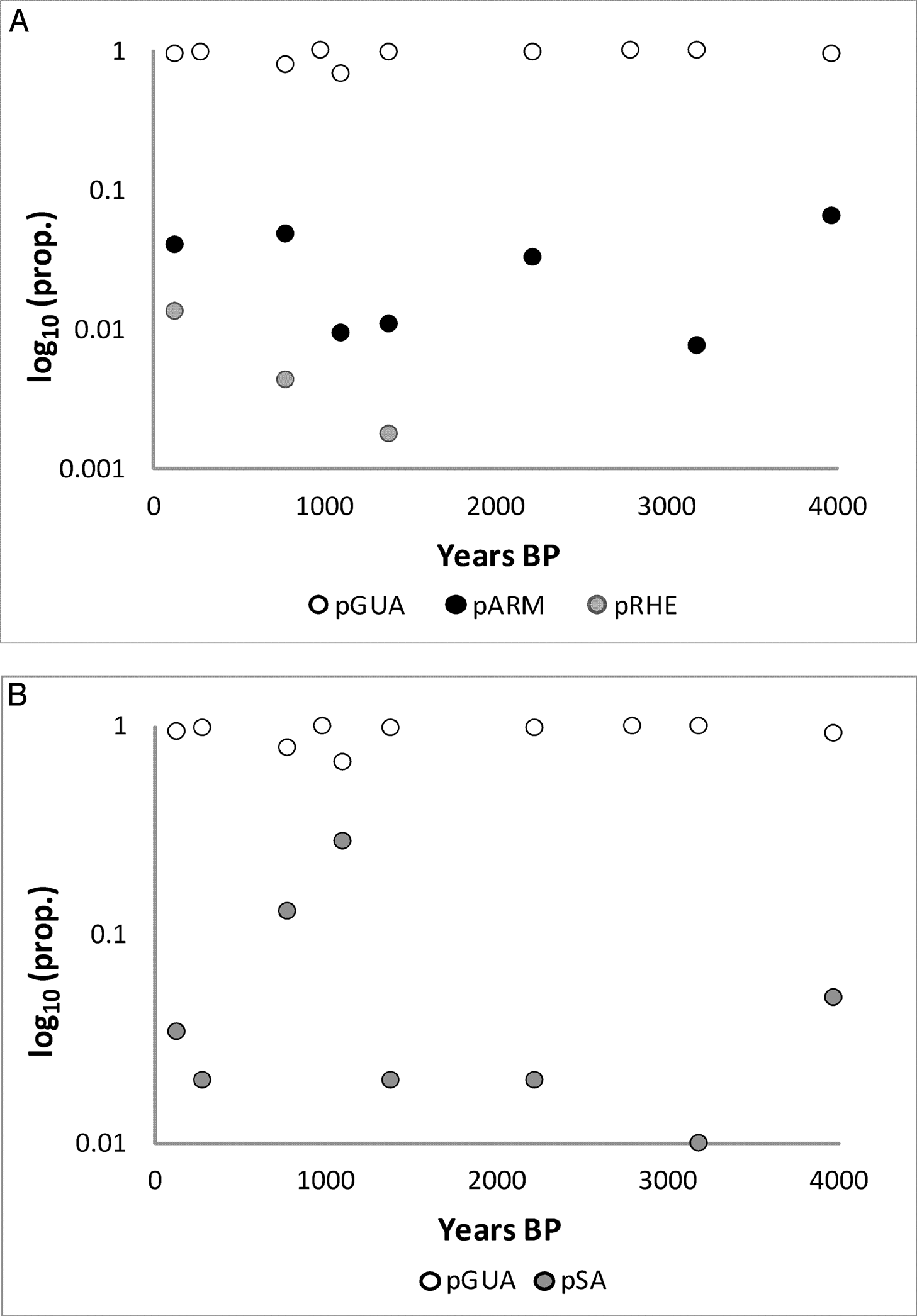
pGUA in comparison to pARM and pRHE (A) and in comparison to pSA (B) during the late Holocene for the mountain subregion. Note the Log10 y-axis; “prop.” stands for proportional abundance.
Relative abundance data from faunal assemblages.
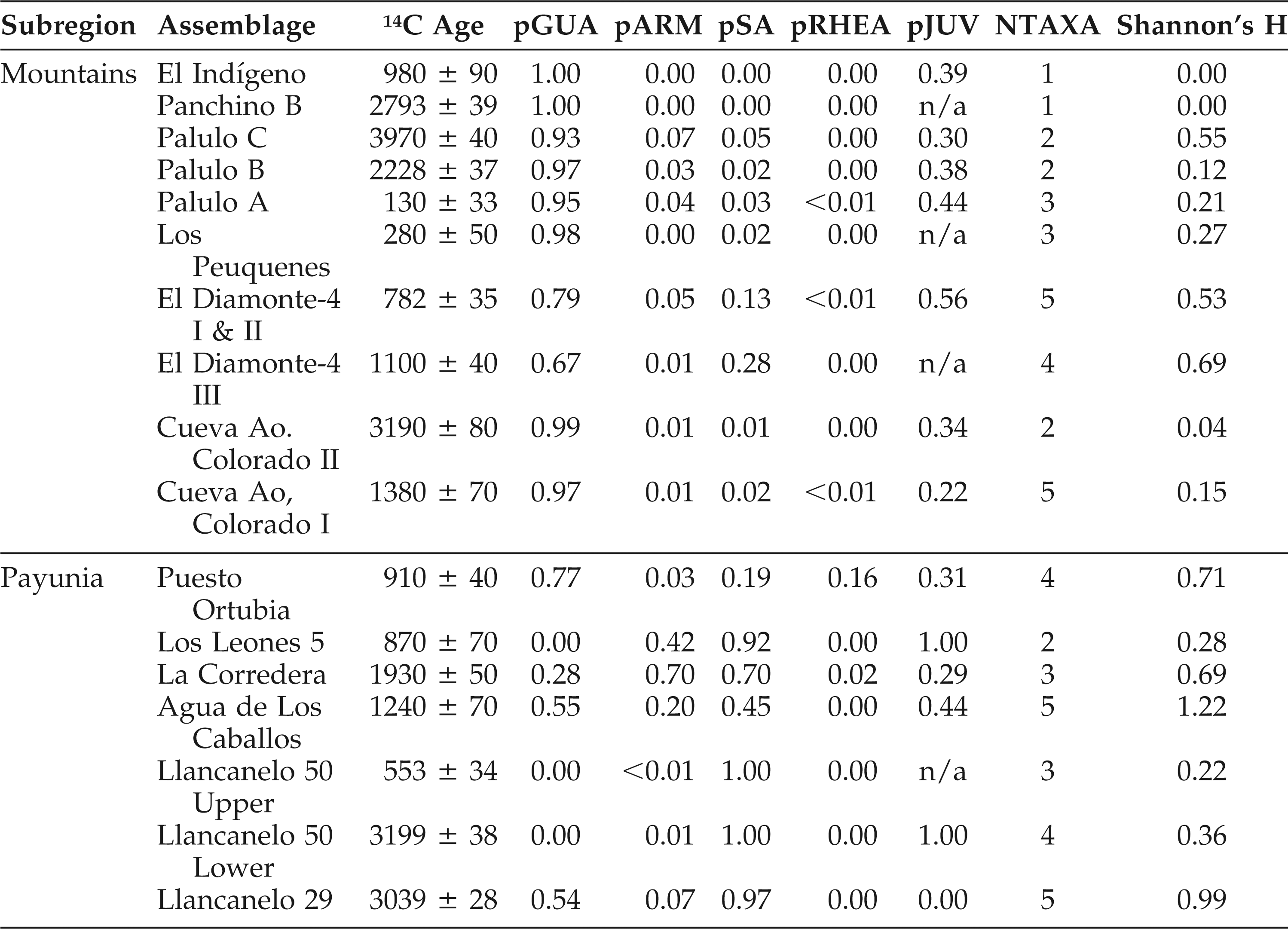
Taxonomic richness measured as NTAXA during the late Holocene in the mountain subregion.
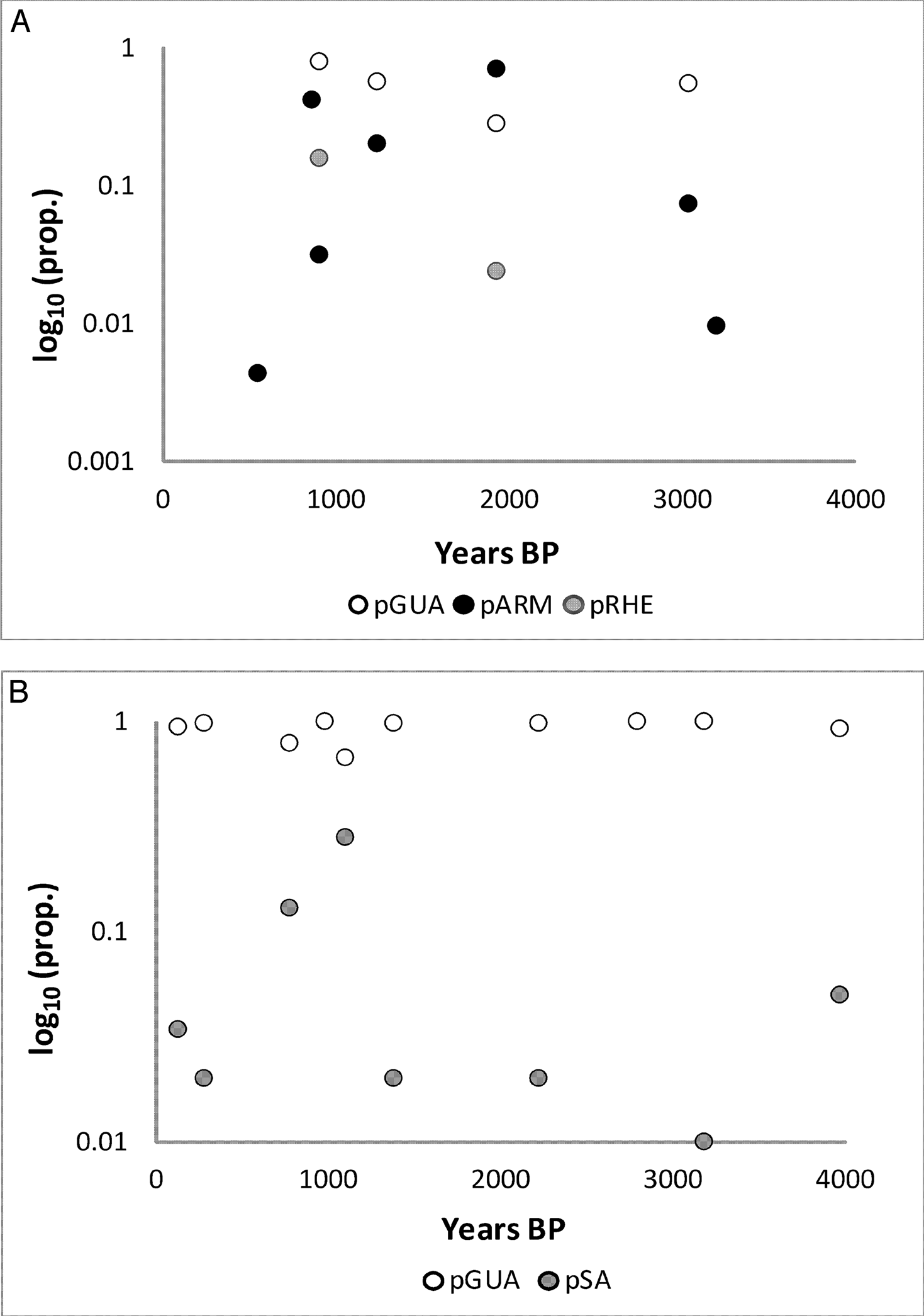
pGUA in comparison to pARM and pRHE (A) and in comparison to pSA (B) during the late Holocene for the Payunia subregion. Note the Log10 y-axis for A but not B; “prop.” stands for proportional abundance.
It appears that guanacos were sustainably hunted in both regions with no decline in abundance over time. However, it is unclear if the high proportional abundance in the mountains is due to sustainable harvest or simply to the fact that guanaco populations were more abundant than populations of other prey species in that subregion. Further, the slight increase in NTAXA over time (Figure 4) might indicate that diet breadth expanded, perhaps suggesting a reduction in foraging efficiency over time. Given the long-term focus on guanaco exploitation, an alternative test implication with which to examine the cumulative effects of harvest pressure is to determine whether or not guanaco survivorship steepened over time. There is no change over time in the proportion of juvenile guanaco remains in faunal assemblages in either subregion (Figure 6; Table 2). Median pJUV is similar between the two subregions, at 0.31 in the Payunia and 0.36 in the mountains. What differs between the subregions is the variability in pJUV over time. In the mountains the interquartile range is 0.12, but in the Payunia it is 0.44, indicating much higher variability in pJUV in the Payunia. Samples of guanaco remains are not large in either region, but they are very small in the Payunia, which may explain the high variability in pJUV for that subregion. An additional line of evidence with which to consider whether or not guanaco population size declined during the late Holocene in future studies, is ancient DNA analysis of haplotype diversity (following Broughton et al. 2013). Although this line of evidence is beyond the scope and capability of the current study, if exploitation depression occurred, a reduction in haplotype diversity is expected.
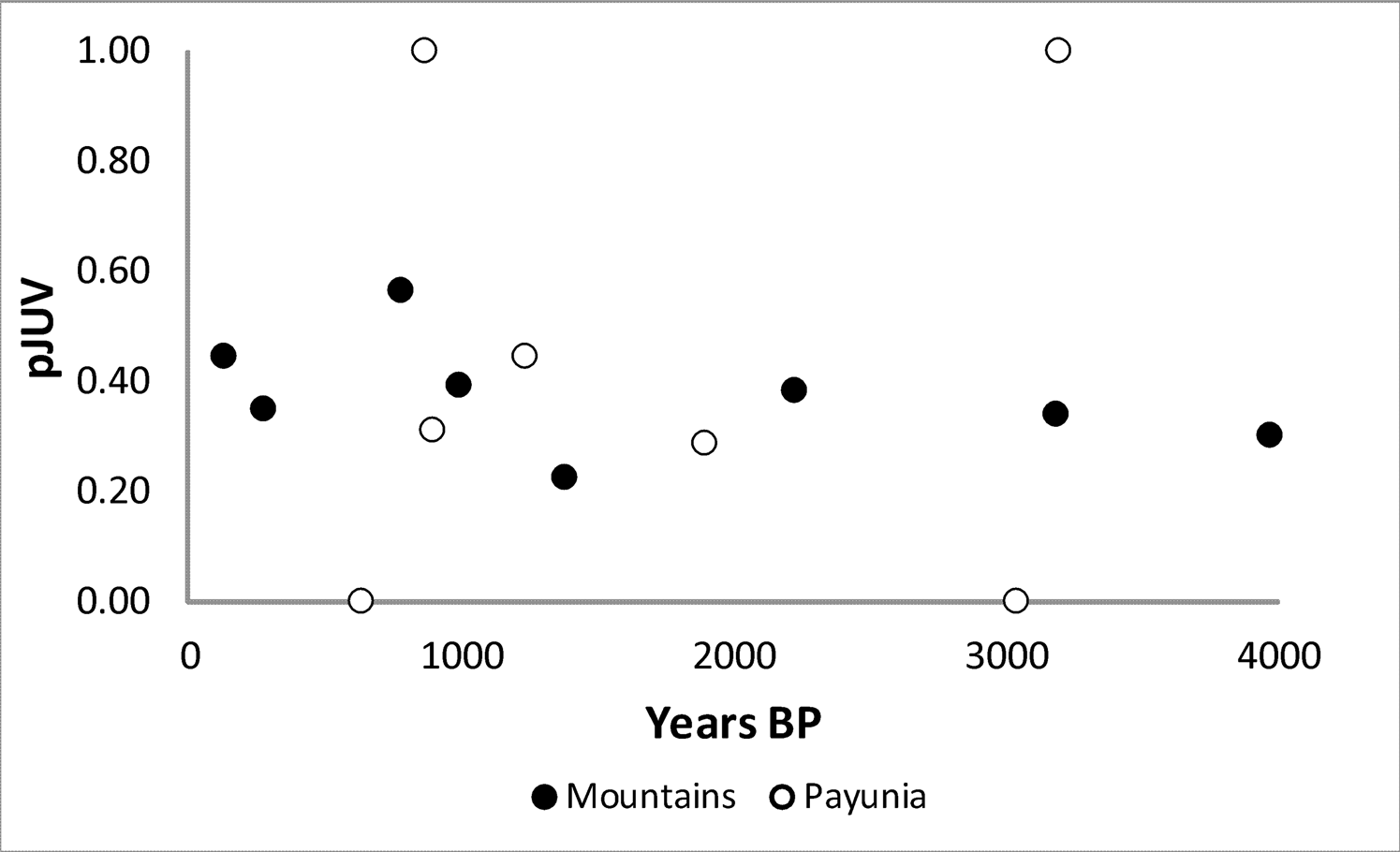
pJUV during the late Holocene for the mountain and Payunia subregions.
Despite subregional variability in climate records (Navarro et al. 2012; Páez et al. 2010, Zárate et al. 2010), generally modern climate was established during the late Holocene at roughly 4000 years ago. There appear to have been no major shifts in elevation-related distributions of habitat zones and environmental productivity. Although it is not the subject of this study, the majority of faunas analyzed here have received detailed taphonomic consideration in other publications (Fernández 2012; Fernández et al. 2009; Giardina 2010; Otaola 2013; Otaola et al. 2012, 2014); remains were commonly deposited by humans, owls, and carnivores and most faunas exhibit minor to moderate weathering and differential preservation of bone. Only remains of taxa typically considered to relate to human subsistence are included in the relative abundance measures employed in this study (Corbat et al. 2009; Giardina 2010; Otaola 2013).
In summary, each of the lines of evidence indicate a more diverse set of foraging strategies in the Payunia than in the mountains, which relates to a more diverse animal community there. In addition, we suspect that this has to do with the mountains being a productive, seasonal resource patch supporting guanaco populations (see discussion below). People did not live in the mountains (above 2000 masl) during the winter, and moved there to focus on guanaco exploitation during the summer. There is a slight increase in NTAXA suggesting an increase in diet breadth in the mountains over time, but prey choice and guanaco survivorship do not shift in either subregion, suggesting that sustained guanaco hunting did not lead to resource depression during the late Holocene.
Discussion
Archaeologists typically cannot directly observe foraging behaviors that relate to prey and patch choice; often foraging efficiency is studied through time-averaged aggregations of archaeological assemblages representing multiple foraging bouts (Grayson and Cannon 1999; Lyman 2003; Otaola et al. 2015a). In contemporary societies, TEK that relates to use of natural resources can be nuanced, detailed, and ephemeral, which Wyndham (2009) terms “subtle ecologies.” Such TEK is accumulated over time within cultures as “ecological understanding” through “incremental learning” (Turner and Berkes 2006:497), which can lead to sustainable resource use. Alternatively, in many cases despite nuanced TEK, foragers may not anticipate future shortages of prey and cause resource depression (Broughton 1994a, 1994b; Butler 2000; Cannon 2003; Nagaoka 2002a, 2002b). It is generations’ worth of TEK that informs choices about which prey to hunt and where to hunt animals (in what quantities) that are observed over time in many OFT studies (e.g., Munro 2004; Nagaoka 2002a, 2002b, 2005, 2006; Wolverton et al. 2012). Despite the power of TEK through enculturation, foraging represents an interplay between such knowledge and environmental contingencies of changes in landscape ecology (Berkes 1993; Johnson and Hunn 2010). We have observed such a dynamic during the late Holocene in western Argentina.
During the mid-Holocene (7000 to 4000 BP) people occupied hunting and gathering camps at the transition between the lowlands and highlands (at roughly 2000 masl) at low population densities (Neme and Gil 2009). In the lowlands (e.g., much of the Payunia) ranging below 2000 masl, water resources were scarce during the entire Holocene, but particularly during the middle Holocene (Gil 2006). During the late Holocene (after 4000 BP) regional population began to increase (Gil et al. 2014; Neme and Gil 2009), and areas above 2000 masl were occupied more frequently during summers.
Geographic expansion and increase of human populations appears to have continued throughout the late Holocene. In an isotopic archaeology study of human remains from southern Mendoza, Neme et al. (2013) found that δ13C in samples dated to 4000 to 2000 BP from the mountains, the piedmont, and the lowlands exhibit similar enrichment, suggesting high mobility and common patterns of resource use across these environments. Although there are no data on human bones from the mountains for the period after 2000 BP, Neme et al. (2013) found substantial separation in δ13C signals between samples from the piedmont and the lowlands, which may indicate that mobility declined. Lower δ13C values in the lowlands may relate to some consumption of maize, though additional research on if and when farming became important in the region needs to be conducted (Gil et al. 2009). Interestingly, during the terminal late Holocene, geographic extent of animal resource use expanded even farther, reaching above 3000 masl after 2000 BP (Neme 2007). In sum, the patterns of late Holocene animal resource exploitation in the Payunia and mountain subregions described in this paper played out in the context of regional population increase and expansion and shifting scales of resource use rooted in the mid-Holocene.
During the late Holocene the mountain subregion would have been a highly productive resource patch in terms of ”ecologically and evolutionarily relevant net primary productivity” (eNPP), which would have led to high seasonal carrying capacity for guanaco. Net primary productivity is the conversion of carbon dioxide and water into energy-rich plant tissues via photosynthesis after subtracting the costs of plant respiration. eNPP is seasonal primary productivity that relates to available nutrients in soils during the growing season (Huston 1993, 2012; Huston and Wolverton 2009, 2011; Wolverton et al. 2009). Subalpine and alpine environments are likely to have high eNPP in valley bottoms where soils form because mountain building, glaciation, and low levels of alpine vegetation lead to concentration of sediments high in soil micronutrients in such settings. This combination of factors leads to productive seasonal plant growth during periods of snowmelt. On an annual basis, productivity is limited by seasonal climate patterns, but during growing seasons, eNPP can be high. This seasonal productivity is available as nutrition for those animal species that can either survive the harsh high-elevation winters or that can travel via seasonal migration. Such a seasonal resource patch must be productive to balance the energy requirements of survival or migration in animals (Huston and Wolverton 2011; Mysterud et al. 2001). For humans, this eNPP becomes available as edible plants and in the form of animal resources as ”ecologically and evolutionarily relevant net secondary productivity” (eNSP), if they can exploit those animal populations.
Guanacos are large ungulates that are dietary generalists and are able to take advantage of the seasonal resource pulse of eNPP in the Andes during the southern hemisphere warm season (roughly November through mid-March) (Puig et al. 1996). A study by Puig et al. (2011) supports that the generalization about eNPP summarized above for high-elevation river valleys indeed applies today to guanaco populations in western, southern Mendoza. Guanaco dietary preferences vary between populations, but in general lowland habitats provide fewer preferred browse species (Puig et al. 2011, 2014). High elevation wetlands (in valley bottoms) and valley slopes boast a much richer variety of grasses, dense succulent monocots, and forbs than do lowland, winter habitats (see also Bahamonde et al. 1986). Given the lower guanaco food diversity in lowland winter range and the higher eNPP in highland summer habitat, it is less likely that guanacos migrate to high elevations; rather “guanacos would migrate toward low altitudes when the snow cover is too deep to survive, and would return toward high altitudes as soon as snow melting exposes the vegetation” (Puig et al. 2011:733).
Given the productivity of highland resources for guanaco populations today, and given the consistently high proportional abundance of guanaco remains in the mountain zooarchaeological assemblages, guanaco exploitation is best explained using the patch choice model from evolutionary ecology (Charnov 1976; see Nagaoka 2002a, 2002b). Though resources from the highland patch were only available for five to six months per year, late Holocene foragers consistently followed guanaco populations into the mountains to take advantage of the seasonal pulse in eNSP (Figure 3). At lower elevations, a more diverse array of lower ranked prey species was exploited (Figure 4). The gains of traveling to the mountains to exploit guanaco must have outweighed the benefits of foraging at lower elevations year round. It is not known how far people traveled to use the mountain resource patch, but this distance appears to have declined after 2000 BP given the isotopic evidence presented by Neme et al. (2013). Recall that after 2000 BP, the δ13C signal in human bone varies by subregion, which Neme et al. (2013) suggest relates to a reduction of mobility and an increase in human populations.
It is important to note that we have only discussed animal resources, primarily related to guanaco exploitation. The proposed distribution of eNPP between the mountains and other subregions provides a mechanism underlying guanaco migration, but as with human populations, it is unclear how far guanacos moved during their seasonal migrations. There may have been year-round, non-migratory populations in some lowland areas. On the escarpment of the Andes in the piedmont subregion, it is likely that guanacos migrated to the mountains during the entire late Holocene. Today the distribution and connectivity of guanaco populations is too fragmented to determine the extent of migration interactions, particularly from distant lowlands, such as the desert plains and large parts of Payunia. However, migration from the piedmont to the mountains still occurs in western, southern Mendoza (Puig et al. 2011, 2014).
Though highly productive for guanacos, the mountain patch would not have been as rich in terms of plant resources for humans during the late Holocene. Indeed, desert plains and volcanic Payunia lowlands boasted a higher diversity of plant resources that were used by humans (Llano 2012). If southern Mendoza witnessed human population growth during the late Holocene, it is thought that this high diversity of plant resources would have become progressively more important over time (Neme et al. 2013).
Conclusion
The assumption that environment provides constraints upon and opportunities within which human culture takes place over time is embedded in our use of OFT from evolutionary ecology. In the case of southern Mendoza, our interpretations of human foraging behaviors over time represent the impacts of many short term foraging bouts, annual, and seasonal migrations. We assume that nuanced use of such resources required detailed traditional ecological knowledge. Although it is difficult to precisely reconstruct individual foraging bouts, this does not mean that variation in culture is not captured in these time-averaged assemblages. How would humans have known how to survive annual and seasonal oscillations in resource availability? How would their knowledge about when and how to exploit guanaco populations have been handed down from generation to generation? As human populations grew in the region, what would have been the mechanism to ensure successful and sustainable resource use over time? We surmise that the answer is agency, knowledge, culture. If TEK informed foraging decisions during the late Holocene in southern Mendoza, it does not appear to have led to resource depression over time; rather, it appears to represent an example of sustainable use of a highly productive resource patch. Alternatively, human population growth was not high enough to lead to resource depression.
Footnotes
Acknowledgments
The authors gratefully acknowledge comments from colleagues at the 2014 meeting of ICAZ in San Rafael, Argentina, constructive criticism from three anonymous reviewers, as well as editorial comments from Kris Gremillion and Dana Lepofsky.
1
A thorough review of the optimal foraging theory literature is not provided in this paper. For details of application within ecological anthropology see references in Smith and Winterhalder (1992). For consideration of OFT in general see Stephens and Krebs (1986), and for details of application in archaeology see references in Broughton and Cannon (2010) and Nagaoka (2001).