
Abstract
We have previously demonstrated that fasting induced apoptosis and decreased cell proliferation in the rat intestinal mucosa. The aim was to investigate the effect of expanded polystyrene as indigestible material on apoptosis and cell proliferation in rat small intestinal mucosa during fasting. Male SD rats were divided into 3 groups. The first group was fed with chow and water ad libitum. The second group fasted for 72 hrs. The third group was fasted for 24 hrs and was fed expanded polystyrene. Intestinal apoptosis was evaluated by percent fragmented DNA assay, terminal deoxynucleotidyl transferase–mediated dUDP-biotin nick end-labeling (TUNEL) staining, and caspase-3 assay. Cell proliferation was analyzed by 5-bromo-2′-deoxyuridine (5-BrdU) uptake. Truncal vagotomy was performed to evaluate a role of the central nervous system. In the 72-hr fasted rat, mucosal height of the rat jejunum was decreased to 73% of that in rats fed ad libitum, and this decrease was partly restored to 90% in rats fed expanded polystyrene. The fragmented DNA was increased in fasted rats (28.0%) when compared with that in rats fed ad libitum (2.6%). The increase in fragmented DNA in fasted rats was recovered by feeding them expanded polystyrene (8.3%). TUNEL staining confirmed this result. The effect of polystyrene on apoptosis was decreased by truncal vagotomy. Expression of cleaved caspase-3 was increased in fasted rats, which was then decreased by feeding of expanded polystyrene. In contrast to apoptosis, feeding of expanded polystyrene had no reconstructive effect on 5-BrdU uptake in the intestinal epithelium, which was decreased by fasting to 60% of that in rats fed ad libitum. In conclusion, feeding of indigestible material partly restored the decrease in intestinal mucosal length in the fasted rats through the apoptotic pathway without any influence on BrdU uptake. Further exploration focused on the mechanism of this effect of indigestible material is required.
Morphologic homeostasis of the small intestinal epithelium is precisely regulated by cell proliferation and cell death, including apoptosis (1–4). We previously demonstrated that (i) apoptosis in the small intestinal mucosa is increased in the fasted rat (5), (ii) the increase in intestinal apoptosis in fasted rats is mainly regulated by the type I pathway, whereas the increase in apoptosis in damaged intestinal mucosa caused by ischemia-reperfusion is regulated by the type II pathway (6), and (iii) apoptosis in the small intestine is partly regulated by the central nervous system, which controls feeding behavior (4, 7–9).
Besides regulation by the central nervous system, cell proliferation and apoptosis are mainly regulated by peripheral factors, including nutrients (1–5). We previously demonstrated that lingual factors in addition to luminal nutrients are important for regulation of intestinal cell proliferation (10). However, it has not been reported whether indigestible material has any effect on intestinal cell proliferation. Intestinal mucosal atrophy has been observed during fasting (5, 6), and several reports indicated that indigestible material has beneficial effects on the atrophic intestinal mucosa (11, 12). This study aimed to examine the effect of indigestible material, expanded polystyrene that was not digested in the rat intestine (13), on cell proliferation and apoptosis in the rat jejunal (small intestinal) mucosa during fasting.
Materials and Methods
Animals and Surgery.
Male Sprague-Dawley rats were used in this study. Rats were housed individually in wire-bottomed cages to prevent coprophagia. They were placed in a room illuminated from 08:00
Collection of Intestinal Mucosa.
After fasting or eating expanded polystyrene, animals were anesthetized and then euthanized by exsanguination. The entire small intestine was carefully removed and placed on ice. The oral 10-cm part of the intestine was treated as the duodenum, and the rest of the intestine was divided into two equal segments representing proximal (jejunum) and distal (ileum) segments, and the jejunum was used in this study. Some pieces, approximately 2 cm in length, were resected from the middle portion of each segment and fixed in 10% neutral buffered formalin for measurement of mucosal height and for immunohistochemistry. Each segment was rinsed thoroughly with physiologic saline and opened longitudinally to expose the intestinal epithelium. The mucosal layer was harvested by gentle scraping of the epithelium with a glass slide as previously described (15). Half of the specimen was used for the DNA fragmentation assay, and the remaining half was used for apoptotic proteins assay with Western blot analysis.
DNA Fragmentation Assay.
Mucosal scrapings were processed immediately after collection to minimize nonspecific DNA fragmentation. Amounts of fragmented DNA were determined as previously described (15, 16) with some modifications. Briefly, mucosal scrapings from different intestinal segments were homogenized in 10 volumes of lysis buffer (5 mM Tris-HCl, 20 mM ethylenediaminetetra-acetic acid [EDTA], and 0.5 % [w/v] Triton X-100, pH 8.0). One-milliliter aliquots of each sample were centrifuged for 20 mins at 27,000 g to separate the intact chromatin (pellet) from the fragmented DNA (supernatant). The supernatant was decanted and saved, and the pellet was resuspended in 1 ml Tris buffer (pH 8.0) with 10 mM Tris-HCl and 1 mM EDTA. Pellet and supernatant fractions were assayed for DNA content by using a diphenylamine reaction as previously described (17). Results were expressed as the percentage of fragmented DNA.
Mucosal Height and Immunohistochemical Staining.
Tissue samples were removed from the jejunum and were immediately fixed in 10% neutral buffered formalin. Samples were then embedded in paraffin and sectioned. Specimens were stained with hematoxylin and eosin. Villus height, crypt height (depth), and mucosal height (villus height plus crypt depth) were measured with a light microscope using micrometer scale standards for fed ad libitum and fasted rats. Fragmented DNA was stained by using the terminal deoxynucleotidyl transferase–mediated dUDP-biotin nick end-labeling (TUNEL) method (18) with some modifications using an Apop Tag Kit (Oncor, Gaithersburg, MD). Specimens were dewaxed and immersed in phosphate-buffered saline containing 0.3% hydrogen peroxide for 10 mins at room temperature and then incubated with 20 μ g/ml proteinase K for 15 mins at room temperature. Seventy-five milliliters of equilibration buffer were applied directly onto the specimens for 10 mins at room temperature; subsequently 55 μ l of terminal deoxynucleotidyl transferase was added, and the mixture was incubated at 37° C for 1 hr. The reaction was terminated by transferring slides to prewarmed stop/wash buffer for 30 mins at 37° C. Specimens were covered with a few drops of rabbit serum, incubated for 20 mins at room temperature, covered with 55 μ l antidigoxigenin peroxidase, and incubated for 30 mins at room temperature. Specimens were then soaked in Tris buffer containing 0.02% diaminobenzidine and 0.02% hydrogen peroxide for 1 min for color development. Finally, specimens were counter-stained with hematoxylin. A minimum of 50 crypts were randomly selected for apoptotic index analysis, and the number of apoptotic cells was calculated. The apoptotic index was determined by dividing the number of cells by the total number of cells in the crypt column and multiplying by 100.
Mucosal cell proliferation was assessed by using 5-bromo-2′-deoxyuridine (5-BrdU) incorporation to identify cells in the S phase of the cell cycle (19). Rats were given 120 mg/kg 5-BrdU (40 g/l 5-BrdU and 4 g/l 5-fluorodeoxyuridine) at 90 mins before sacrifice. 5-BrdU was detected with a monoclonal antibody and a streptavidin–biotin staining system. The number of labeled cells in at least 10 well-oriented longitudinal crypts in each sample was determined under a light microscope by an examiner blinded to the study groups, and the number was reported as the number of 5-BrdU–labeled cells among the total crypt cells.
Isolation of Cytosolic Proteins.
Mucosal scrapings were immediately washed twice with ice cold phosphate-buffered saline (pH 7.4) and centrifuged at 1,000 g for 5 mins at 4° C. Pellets were then resuspended with two volumes of buffer A and lysed at 4° for 30 mins (20). Buffer A consisted of 250 mM sucrose (Sigma, St. Louis, MO), 20 mM N-[2-hydroxyethyl]piperazine-N,-[2-ethanesulfonic acid] (Sigma), 10 mM KCl, 1.5 mM MgCl2, 1 mM EDTA, 1 mM EGTA, 1 mM dithiothreitol, 1 mM phenylmethylsulfonyl fluoride, 10 μ g/ml aprotinin, 10 μ g/ml leuptin, and 1.8 mg/ml iodoacetamide). Homogenates were centrifuged at 1,000 g for 10 mins at 4° C to remove cell nuclei, and supernatants were again centrifuged at 10,000 g for 15 mins at 4° C to remove mitochondria. Supernatants of the 10,000 g spin were further centrifuged at 1,000,000 g for 1 hr at 4° C. The resulting supernatant represented the soluble cytosolic fraction, and pellets were the membranous fraction. Supernatant and pellet fractions of resuspended solutions in buffer were divided into multiple samples, which were frozen at − 80° C until immunoblotting.
Western Blot Analysis of Caspase-3.
Cleaved caspase-3 and procaspase-3 were analyzed in cytosolic fractions. Equal quantities (40 μ g) of lysates were resolved by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and electroblotted onto a nitrocellulose membrane (Bio-Rad Laboratories, Hercules, CA). After blocking with phosphate-buffered saline containing 0.1% polyoxyethylene sorbitan monolaurate (Tween-20, Sigma) and 5% skim milk at 4° C overnight, the membrane was incubated with rabbit polyclonal anti–caspase-3 antibody (1:1000; Cell Signaling Technology, Danvers, MA) for 1 hr. Antigen-antibody complexes were detected with horseradish peroxidase–conjugated anti–rabbit IgG (1:500, Santa Cruz Biotech, Santa Cruz, CA). Detection of chemiluminescence was performed by using ECL Western blotting detection reagents (Amersham Pharmacia Biotech, Buckinghamshire, UK). Densitometeric assessment of bands from the auto-radiogram was carried out by using Image Gauge VDS (Fujifilm, Tokyo, Japan). Band intensities were quantified by measuring the absolute integrated optical intensity, which estimates the band in the lane profile. Results were expressed as a ratio to β-actin densitometry units.
Statistical Analysis.
Results were expressed as the means ± SE. Data were evaluated by analysis of variance in which multiple comparisons were performed by the least significant-difference method. Differences were considered significant if the probability of the difference occurring by chance was less than 5 in 100 (P < 0.05).
Results
Jejunal Mucosal Height in the Rats Fasted for 72 hrs and Fed Expanded Polystyrene.
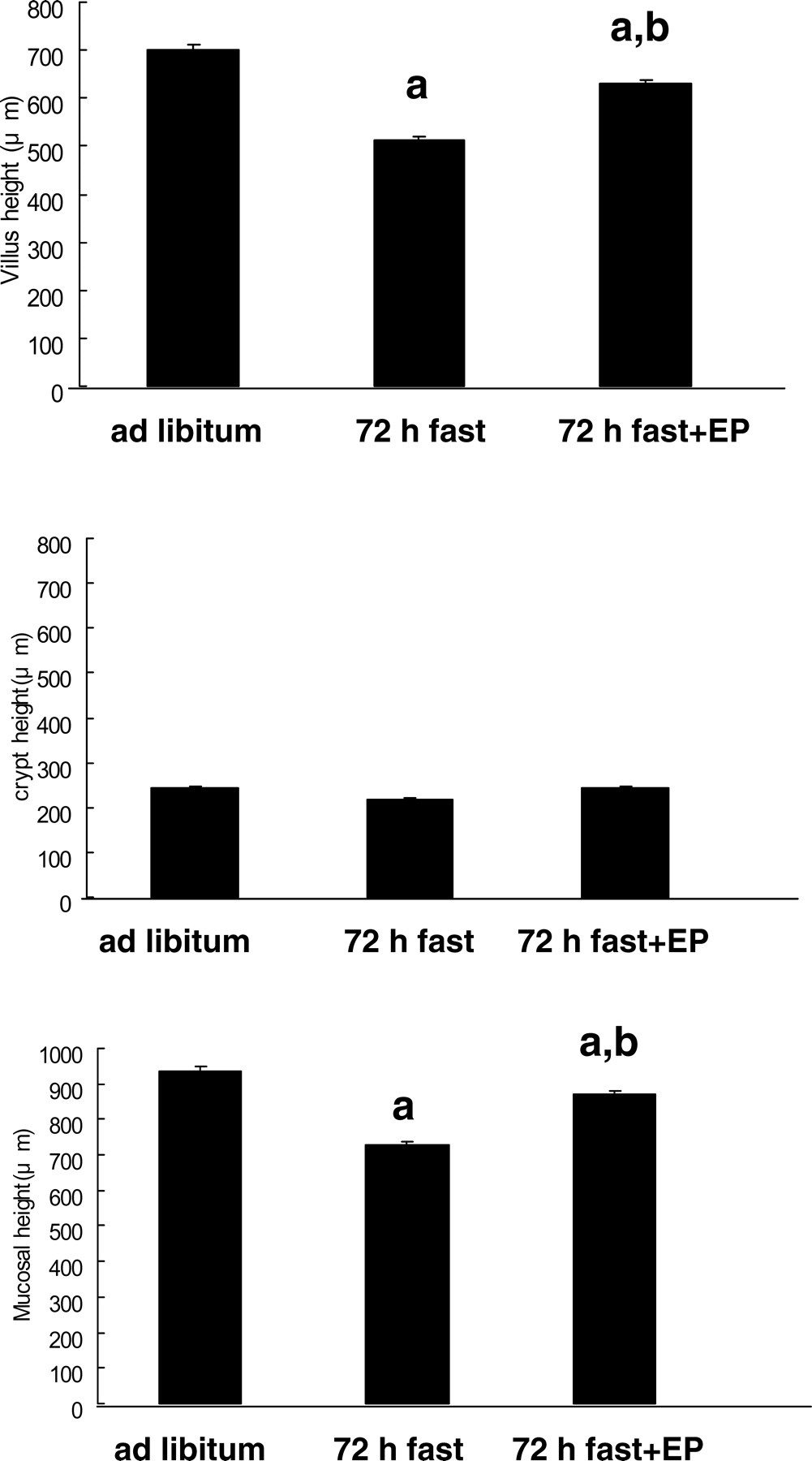
Jejunal mucosal height in the 72-hr fasted rats (729 ± 10 μ m) was significantly shorter than that of the rats fed ad libitum (936 ± 12 μ m) (Figs. 1 and 2, P < 0.05), and this was mainly due to a decrease in villus height (ad libitum, 698 ± 12 μ m; 72-hr fasting, 510 ± 18 μ m) but was not due to crypt height (ad libitum, 246 ± 23 μ m; 72-hr fasting, 216 ± 13 μ m). The decrease in mucosal height in 72-hr fasted rats was reduced as a result of their eating expanded polystyrene (P < 0.05), but the recovery was not complete as indicated in Figure 2 (872 ± 17 μ m). The average volume of expanded polystyrene eaten by rats was 28 cm3.
Cell Proliferation in the Jejunum.
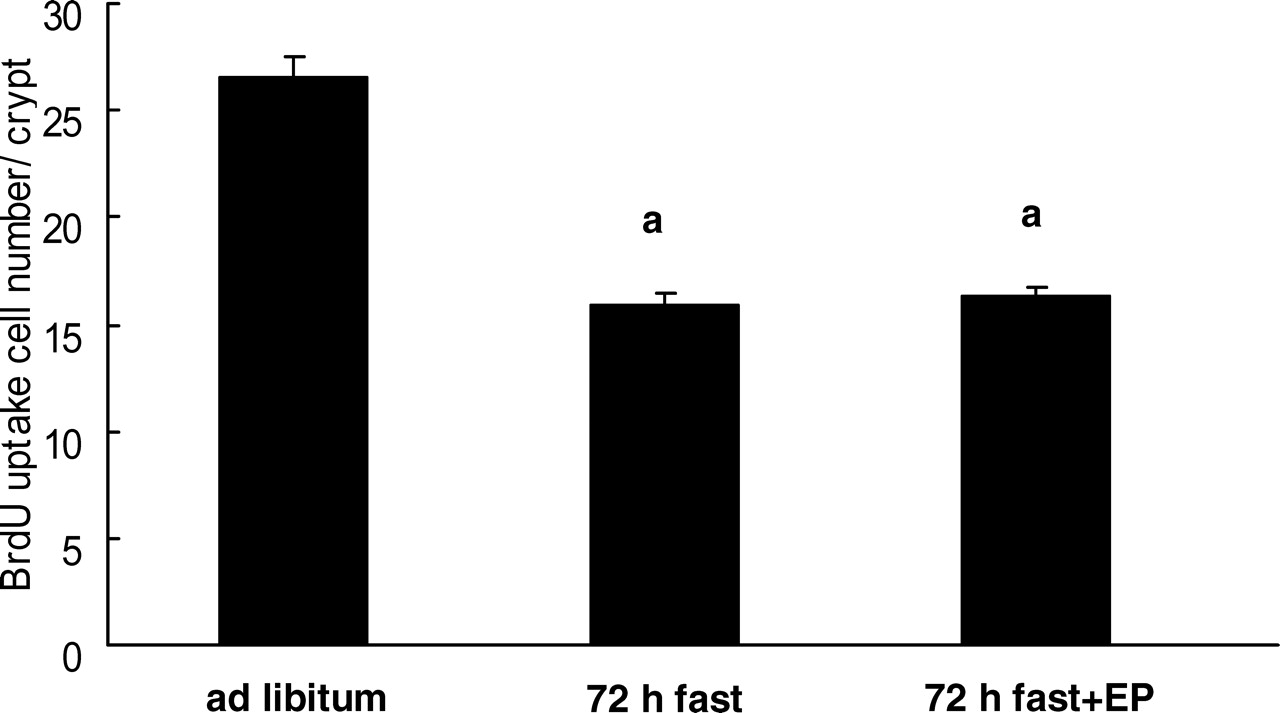
The number of 5-BrdU–positive cells in rats that had fasted for 72 hrs was significantly less than that in rats fed ad libitum (Fig. 3; ad libitum, 26.5 ± 1.6 5-BrdU–positive cells per crypt; 72-hr fasting, 16.0 ± 0.9 5-BrdU–positive cells per crypt). Feeding the rats expanded polystyrene had no effect on the decrease in 5-BrdU–positive cells in 72-hr fasted rats (16.3 ± 1.5 5-BrdU–positive cells per crypt).
Apoptosis in the Rat Jejunal Mucosa.
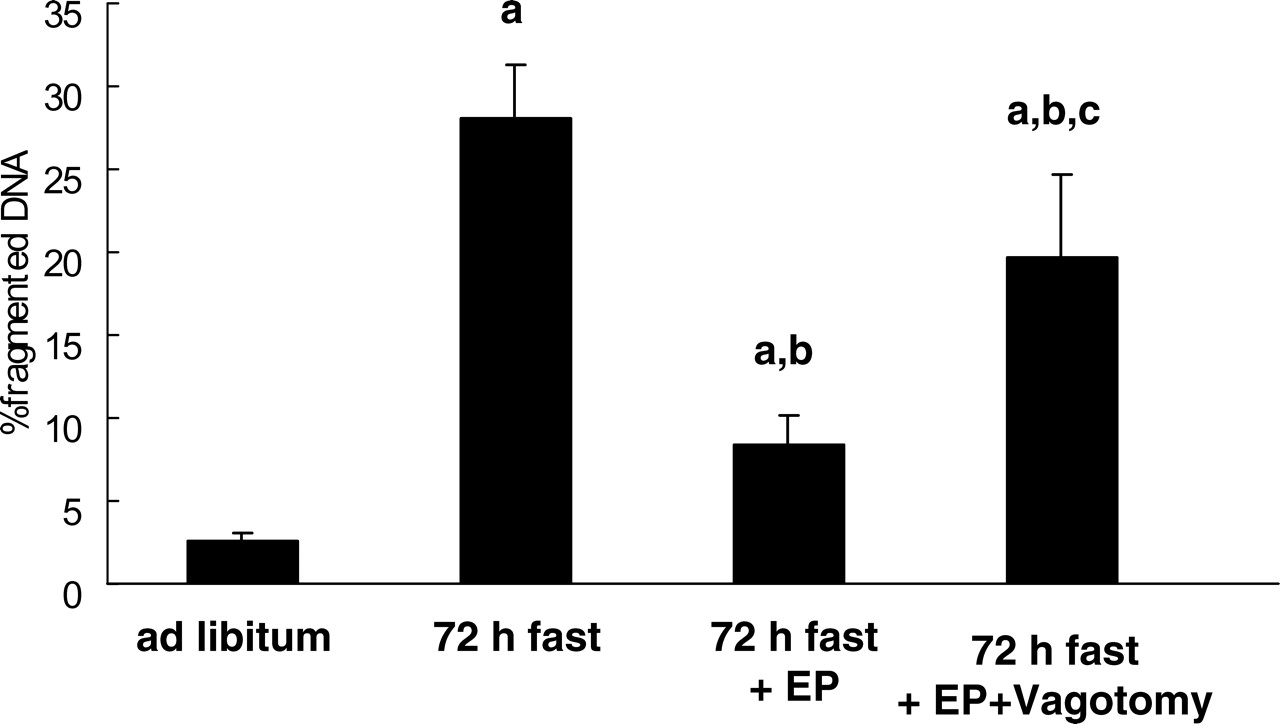
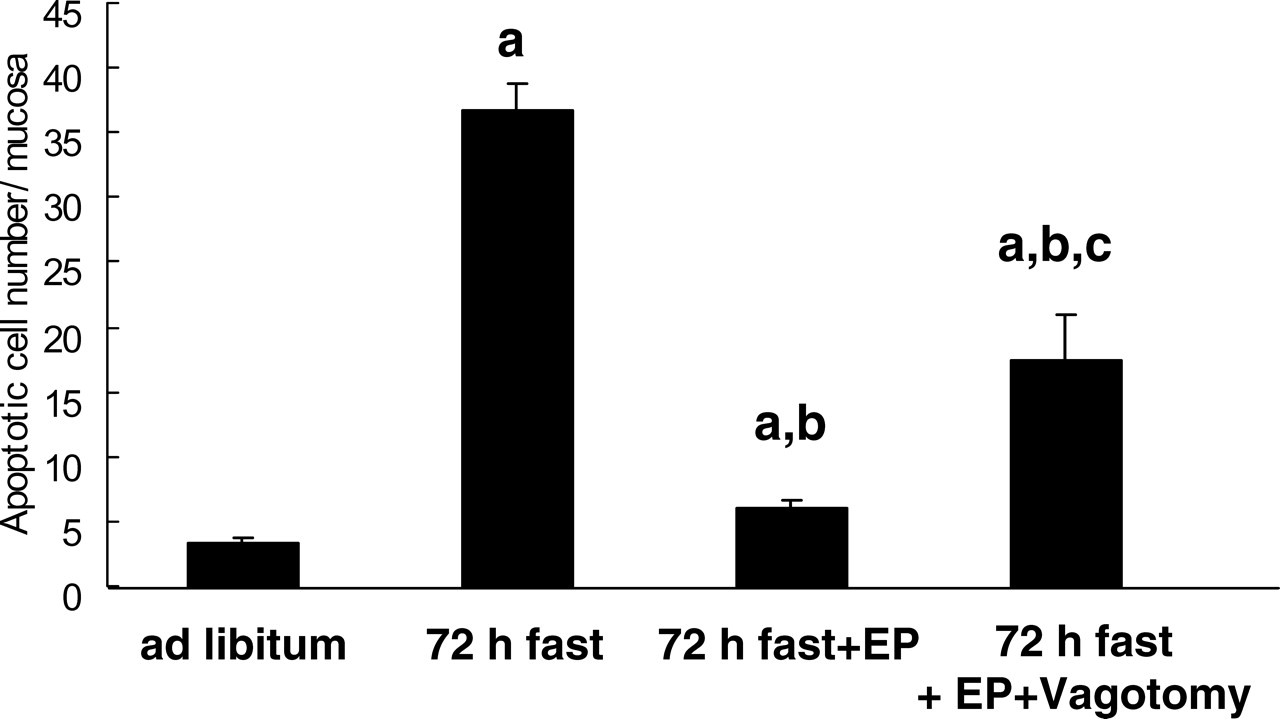
The percentage of fragmented DNA in the jejunal mucosa in 72-hr fasted rats (28.0% ± 3.3%) was significantly higher than that in rats fed ad libitum (2.6% ± 0.4%) (Fig. 4; P < 0.05). Agarose gel electrophoresis of total DNA revealed a distinct ladder of DNA fragments from the jejunum in 72-hr fasted rats (data not shown). These data indicated that enhanced jejunal mucosal apoptosis was induced by fasting. The increase in jejunal mucosal apoptosis was partly recovered by feeding the fasting rats expanded polystyrene (P < 0.05; 8.3% ± 1.9%). This attenuation effect of polystyrene on fasting-induced mucosal apoptosis was diminished by truncal vagotomy (P < 0.05), but it was not completely abolished (19.6% ± 5.0%).
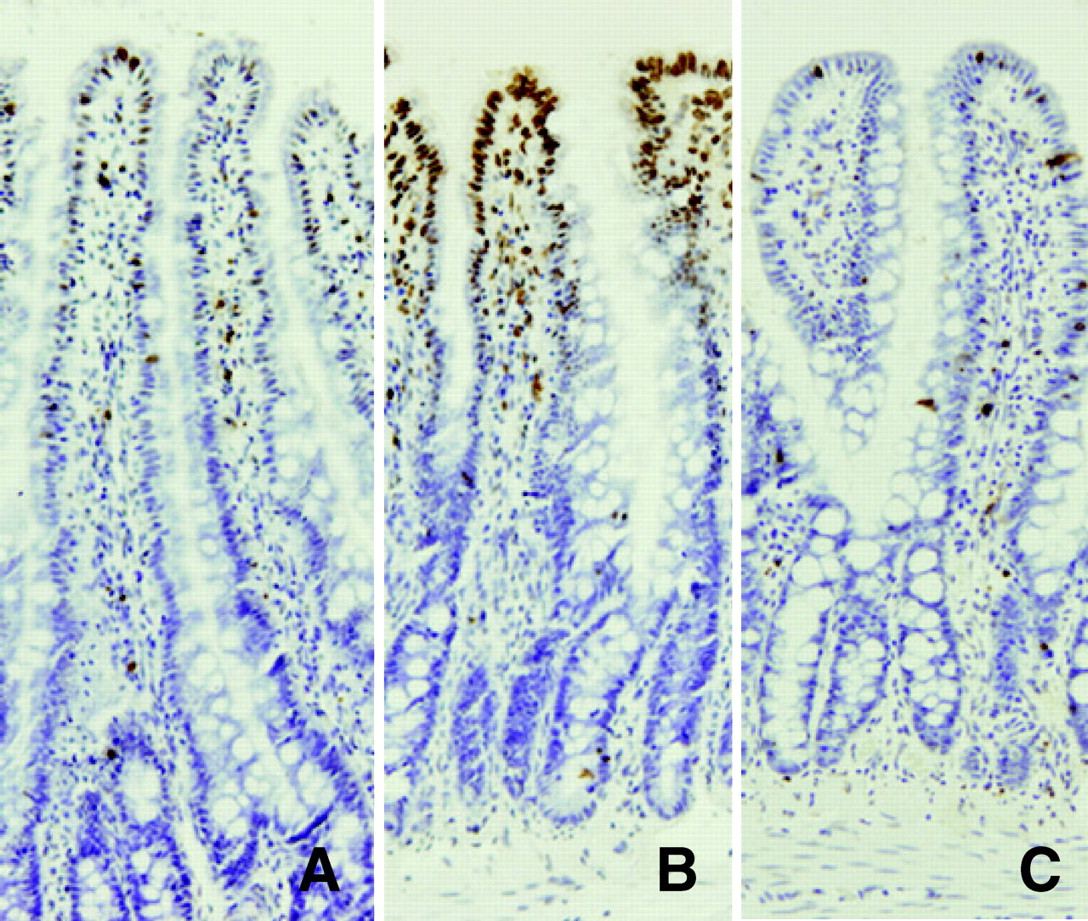
TUNEL staining of the jejunum showed that a few apoptotic cells were present in villus tips in rats fed ad libitum (Fig. 5A), and an increase in the number of apoptotic cells in 72-hr fasted rats was observed in the upper third of the intestinal villi (Fig. 5B). Feeding the rats expanded polystyrene decreased the number of apoptotic cells compared with that in 72-hr fasted rats (Fig. 5C). As indicated in Figure 6, the number of apoptotic cells was increased by 72 hrs of fasting (P < 0.05; ad libitum, 3.4 ± 0.3 apoptotic cells; fasting for 72 hrs, 36.7 ± 2.9 apoptotic cells), and this increase was attenuated by feeding the rats polystyrene (P < 0.05; 6.1 ± 0.4 apoptotic cells). Truncal vagotomy diminished the effect of polystyrene as indicated by the percentage of fragmented DNA (P < 0.05; 17.5% ± 3.4%).
Western Blot Analysis of Caspase-3.
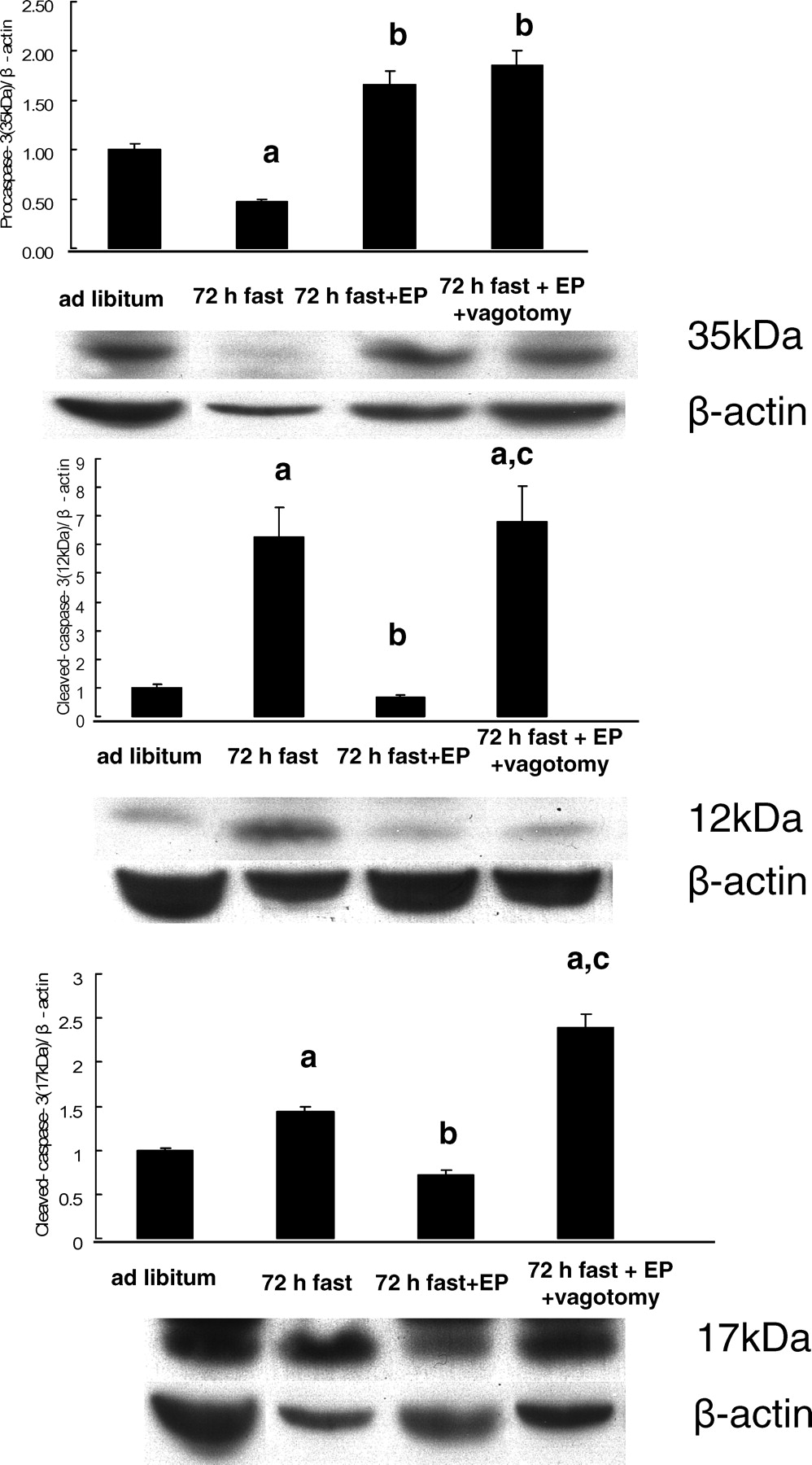
Cleavage of caspase-3 was evaluated in cytosolic fractions (Fig. 7). Expression of cleaved caspase-3 (17 kDa and 12 kDa) was significantly greater in 72-hr fasted rats than in rats fed ad libitum (P < 0.05), and the expression of procaspase-3 (35 kDa) was significantly less in 72-hr fasted rats than in rats fed ad libitum (P < 0.05). This effect of 72 hrs of fasting on procaspase-3 and cleaved caspase-3 was reversed by feeding with expanded polystyrene (P < 0.05). The influence of polyethylene on cleaved caspase-3 was abolished by truncal vagotomy, but vagotomy had no effect on the change in procaspase-3 induced by polystyrene.
Discussion
The results of the present study indicate that the decrease in jejunal mucosal height caused by 72 hrs of fasting was partly restored by feeding the rats indigestible material: expanded polystyrene. This attenuation effect was mainly due to suppression of mucosal apoptosis without any effect on cell proliferation as evaluated by 5-BrdU uptake. This result was supported by the finding that expanded polystyrene restored mucosal height by mainly affecting villus height without having any effect on crypt height, as fasting decreased villus height by inducing apoptosis. As indicated in previous studies (5, 6), apoptotic cells increased mainly in the villous tip; this increase might explain why the mucosal atrophy was limited in the villus, but the precise mechanism has not been demonstrated. Morphologic homeostasis of the small intestinal epithelium is precisely regulated by cell proliferation and cell death, including apoptosis (1–4). Cell proliferation and apoptosis are mainly regulated by luminal nutrients (1, 2, 5, 6) and humoral factors (1, 2, 5, 21–23). In addition to these factors, the present study suggested that mechanical stimuli restored the decrease in intestinal mucosal height with fasting by affecting mucosal apoptosis. The attenuation effect of indigestible material on intestinal mucosal atrophy during fasting might be protective against an increase in bacterial translocation as indicated in previous studies (11, 12), which warrant further exploration.
The present study indicated a trophic effect of polystyrene, which was not digested or fermented in rats (13), on the atrophic intestinal mucosa during fasting. This effect might be induced via several factors, including brain gut peptides, other humoral factors, a neuronal network, mechanical stimuli, and intestinal flora (4, 24, 25). Our previous studies have shown that the central nervous system, which is involved in feeding behavior, and lingual factors regulate intestinal cell proliferation and apoptosis (4, 7, 8–10, 14, 26). This study suggested the central nervous system might be involved in the effects of indigestible material because truncal vagotomy reduced the effect of indigestible material on intestinal apoptosis. Truncal vagotomy itself had no influence on feeding behavior in rats during a single week (14). Our previous studies indicated that the effect of the central nervous system on intestinal apoptosis was not influenced by truncal vagotomy (4, 7–9), although its effect on intestinal cell proliferation was changed by truncal vagotomy (4, 14, 27). The reason for the discrepancy between this study and the previous studies is unknown, but the effect of indigestible material involved afferent and efferent vagal nerves in this study, and only the efferent vagal nerve was involved in the previous studies. We did not evaluate the role of sympathetic nerves because they affect feeding behavior. The role of the autonomic nervous system warrants further exploration.
Although many apoptotic stimuli and signal transduction pathways have been demonstrated, two principal apoptotic pathways are well recognized: the type I pathway activated by extrinsic stimuli and the type II pathway activated by intrinsic stimuli. Our previous study showed that the type I pathway is the main pathway for rat intestinal apoptosis induced by fasting (6). In this pathway, the apoptosome may cleave procaspase-3, and cleaved caspase-3 induces small intestinal apoptosis (6, 28). The present study indicated that procaspase-3 was decreased and cleaved caspase-3 was increased after 72 hrs of fasting. This change in procaspase-3 and cleaved caspase-3 was restored to normal levels by feeding the rats indigestible material, and vagotomy abolished the effect of indigestible material on cleaved caspase-3. These results were consistent with the results of apoptosis evaluated by the DNA fragmentation assay and the TUNEL method. As shown in Figure 5, TUNEL-positive apoptotic cells were mainly located in villus tips of the rat small intestine during fasting as demonstrated previously (5, 6). Hyoh et al. (29) demonstrated that the activity of caspase-3 was increased in the top of the intestinal villus, which supports our results that apoptosis during fasting was induced in the top of the villus by activation of caspase-3 by a type I pathway.
In conclusion, indigestible material (expanded poly-styrene) attenuated a decrease in rat jejunal mucosal height after 72 hrs of fasting by decreasing enhanced apoptosis without having any effect on cell proliferation.
Light micrographs of the rat jejunum (stained with hematoxylin and eosin). (A) Rats fed ad libitum. (B) Rats that had fasted 72 hrs. (C) Rats that had fasted for 72 hrs and were fed expanded polystyrene. Magnification: ×100. A color figure is available in the online version of the journal. Measurement of mucosal height, villus height, and crypt height in the rat small intestine mucosa (jejunum). EP, expanded polystyrene. Values are the means ± SE. Six rats were tested in each group. a, P < 0.05 compared with rats fed ad libitum; b, P < 0.05 compared with 72-hr fasted rats. Effect of fasting and administration of the indigestible material expanded polystyrene (EP) on number of cells with 5-BrdU uptake in the rat jejunal crypt.Values are the means ± SE. Six rats were tested in each group. Thirty crypts were counted in each group. a, P < 0.05 compared with rats fed ad libitum. Effect of fasting and administration of the indigestible material expanded polystyrene (EP) on apoptosis evaluated by DNA fragmentation assay in the rat jejunal mucosa. Values are the means ± SE. Six rats were tested in each group. a, P < 0.05 compared with rats fed ad libitum; b, P < 0.05 compared with 72-hr fasted rats. c, P < 0.05 compared with 72-hr fasted rats fed EP. Light micrographs in the rat jejunal mucosa stained by theTUNEL method. (A) Rats fed ad libitum. (B) Rats that had fasted 72-hrs. (C) Rats that had fasted for 72-hrs and were fed expanded polystyrene. Magnification: × 100. A color figure is available in the online version of the journal. Effect of fasting and administration of the indigestible material expanded polystyrene (EP) on the number of TUNEL-stained cells in the rat jejunal mucosa. Values are the means ± SE. Six rats were tested in each group. a, P < 0.05 compared with rats fed ad libitum; b, P < 0.05 compared with 72-hr fasted rats. Effect of fasting and expanded polystyrene (EP) on the expression of procaspase-3 and cleaved caspase-3. Values are the means ± SE. Six rats were tested in each group. a, P < 0.05 compared with rats fed ad libitum; b, P < 0.05 compared with 72-hr fasted rats.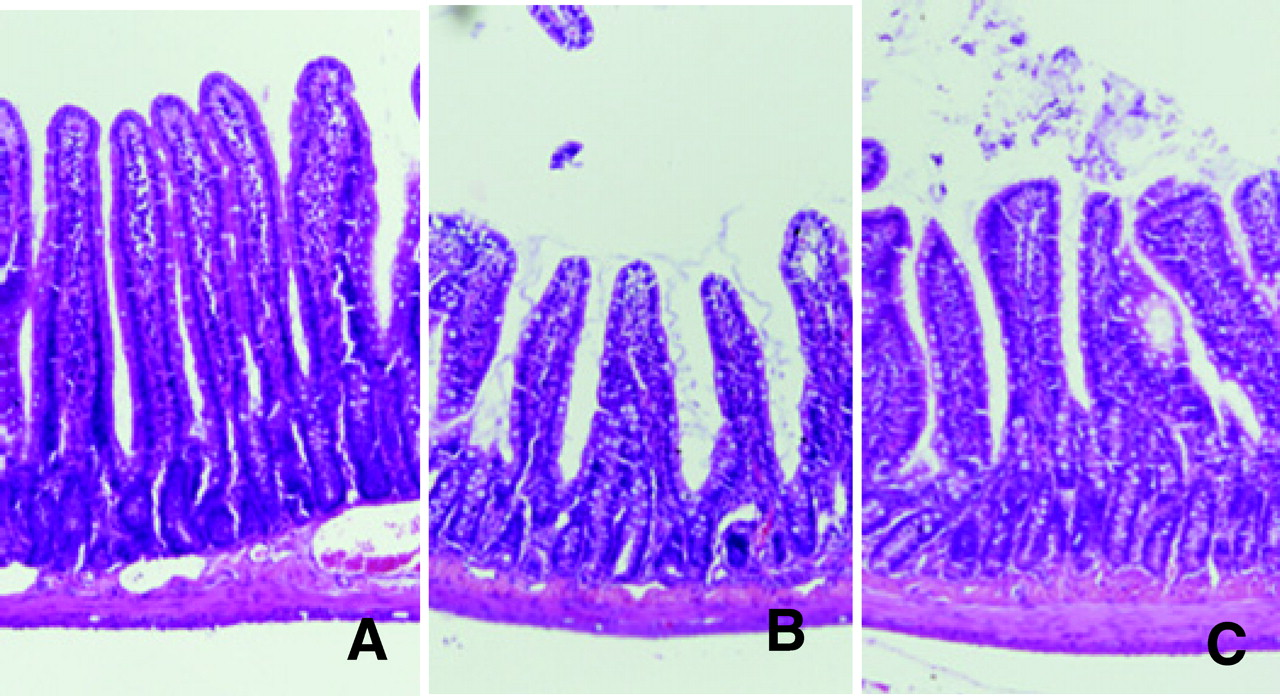
Footnotes
This work was supported in part by the president’s expenditure (research project expenditure) of Saga University and Grants-in-Aid from the Ministry of Education, Science and Culture in Japan for Scientific Research (15590658 and 18590690 to K.F. and 16590605 and 18590691 to R.I.).