
Abstract
Objective: Study the therapeutic effects and immunoregulatory mechanisms of anti-DR5 mAb on adjuvant arthritis (AA) rats.
Methods: AA rats induced by CFA, were treated with anti-DR5 mAb through mainline administration. Effect on the synovial membranes of the tissues was detected by H&E staining. Flow cytometry and MTT assay were used for detecting the induced apoptosis in an in vitro system and TUNEL assay was used for analysis in an in vivo system. The involvement of the apoptotic pathway was further proved by a caspase inhibition assay.
Results: Anti-DR5 mAb could induce synovial cell apoptosis in an in vitro system, which was related with the mRNA expression of DR5 on the cell surface. The mRNA expressions of c-myc and bcl-2 were decreased in synovial cells and those of p21, p53, and bax were increased. The protein expressions of caspase-8/3/9, RANKL, JNK2, and c-Jun were raised and that of bcl-2 was decreased. When the caspase inhibitor was added to the synovial cells treated with anti-DR5 mAb, it showed a dose-dependence inhibition effect, indicating that anti-DR5 mAb inducing apoptosis might be through the caspase pathway.
Conclusion: This study shows that anti-DR5 mAb can ameliorate arthritic symptoms. The mechanisms of the treatment are related to the increase in synovial cell apoptosis by regulating the mRNA expression of DR5 and apoptosis-related genes, prolonging the duration of the cell cycle by modulation of the mRNA expression of cell cycle-related genes, and the protein expression of the molecules in the caspase pathway and RANKL, JNK2, and c-Jun.
Introduction
Rheumatoid arthritis (RA) is an autoimmune disease whose etiology is unknown (1). The resistance of synovial lymphocytes, macrophages, and fibroblasts to apoptosis may be a contributing factor in the persistence of the disease. Electron microscopic examination revealed few cells that displayed morphological features of apoptosis (2, 3). Consistent with the electron microscopy data, recent investigations with terminal uridine deoxynucleotidyl transferase (dUTP) nick end labeling (TUNEL) have demonstrated low frequencies of apoptosis (4, 5). Additionally, rats subjected to experimental arthritis displayed low levels of apoptosis (6), while joint inflammation and destruction in a variety of animal models was ameliorated by enhanced apoptosis (7). Collectively, these data suggest that a viable therapy for RA may be achieved by enhancing the level of apoptosis in the joints.
Another member of the tumor necrosis factor superfamily, namely, the Apo2 ligand (APO2L)/tumor necrosis related apoptosis-inducing ligand (TRAIL), was described to induce apoptosis in a Fas/CD95-independent manner (8, 9). TRAIL interacts with 5 different receptors, of which only DR4 (TRAIL-R1) (10) and DR5 (TRAIL-R2) (11) are capable of transmitting death signal in tumor cells (12). RA fibroblast-like synoviocytes (FLSs) were found to constitutively express DR4 and DR5 on the cell surface. DR5 appears to be the main mediator of TRAIL-induced stimulation, as the RA FLS proliferation induced by an agonistic anti-DR5 antibody was comparable with that induced by TRAIL (13).
Yao et al. (14) examined the ability of human TRAIL, administered by adenoviral (Ad)-mediated gene transfer to eliminate the hyperplastic synovium. Infection with Ad TRAIL of the synovial cells derived from RA patients resulted in significant apoptosis in 3 out of 5 lines. Moreover, primary rabbit synovial fibroblasts were also sensitive to Ad TRAIL. A recent experiment has demonstrated increased DR5 expression on most malignantly transformed cells that can be eliminated by a novel agonistic anti-human DR5 mAb, TRA-8. TRA-8 induces a strong apoptosis response without hepatocellular toxicity (15).
In an in vitro system, TRA-8 induced apoptosis of RA synovial cells and inhibited the production of matrix metalloproteinases induced by pro-inflammatory cytokines. In an in vivo system, TRA-8 effectively inhibited the hypercellularity of an SV40-transformed RA synovial cell line and completely prevented the bone erosion and cartilage destruction induced by these cells (16). These results indicate that increased DR5 expression and susceptibility to DR5-mediated apoptosis are characteristic of the proliferating synovial cells in RA. Since highly proliferating “transformed-appearing” RA synovial cells play a crucial role in bone erosion and cartilage destruction in RA, the specific targeting with an agonistic anti-DR5 mAb of DR5 on RA synovial cells may be a potential therapy for RA.
Materials and Methods
Animals.
Pathogen-free male Sprague-Dawley (SD) rats (200 ± 2 g; 2–3 months of age at the start of the experiments) were obtained from Shanghai Slac Laboratory Animal Co., Ltd. and were maintained in our central animal facility. The Animal Care and Use Committee of the University of XiaMen approved all experiments.
Generation of Anti-DR5 mAb.
The DR5 gene was constructed by over-lapping PCR, then was cloned into pET-22b(+), which contain a six histidine tag. The DR5 used in this study were purified through a nickel ion column (Novagen, USA). BALB/c mice were immunized with sDR5. Mice were boosted three times and the splenocytes were fused with SP2/0-Ag 14 in the presence of 50% polyethylene glycol in complete serum-free DMEM, followed by culturing in DMEM/HAT and DMEM/HT (Sigma, USA) selective media. Supernatants from positive wells were tested for the presence of antibodies that bind sDR5 using ELISA. Hybridomas that produced Abs that bind to sDR5 underwent three rounds of limiting dilution analysis. mAb isotypes were determined by ELISA using a mouse immunoglobulin isotyping ELISA kit (Pharmingen, USA). The mAb used in this study was purified by protein G affinity chromatography (Pierce, USA).
Establishing and Grouping of Rat AA Models.
All rats were immunized at the right paw with a single 0.05 ml intradermal injection of CFA (paraffin oil 8.5 ml and Arlacel A 1.5 ml, containing 6 mg/ml of Mycobacterium bovis, French 1173P2). Clinical signs of arthritis (that is, paw swelling) were usually observed by day 14 and monitored during the course of disease by water displacement plethysmometry every 3 days. Paw swelling was expressed as delta paw volume (that is, with paw volume before onset of arthritis subtracted). The animals divided into five groups (n = 12/group): normal group, model group (AA rats without treatment), AA rats with MTX (Sigma, USA) treatment, AA rats with anti-DR5 mAb (produced by our lab, purity of 95%, IgG 1 (17)) treatment, and AA rats with mouse IgG (Sigma, USA) treatment. For animal treatment, the tail vein was injected at day 15 after immunization in animals for 15 days. The tail skin was prepared with 75% ethanol, and MTX (0.36 mg/ml, 1 ml per rat per week), anti-DR5 mAb (0.2 mg/ml, 1 ml per rat per day), or mouse IgG (0.2 mg/ml, 1 ml per rat per day) were injected mainline in the indicated concentrations in a total volume of 50 μl saline. The course of arthritis was monitored regularly until rats were sacrificed. We stopped treatment at day 30 and one-third of the rats of each group were sacrificed at days 30, 37, and 44 by CO2 inhalation.
Separating and Culturing of Synovial Cells.
AA rats were sacrificed and synovial membrane tissues of each group were harvested with asepsis. The tissues were washed in 200 mg/L of ice-cold D-Hanks added with 200 kU/L penicillin and streptomycin (without Ca2+, Mg2+) for three times, and collected by centrifugation for 15 min at 400 g. The resulting pellets were re-suspended in 2 ml DMEM without serum and 2 ml 0.2% II type of collagenase in culture plate and incubated for 4 h at 37°C, 5% CO2. Then 4 ml 2.5 g/L trypsin was added to the same plate and incubated for another 30 min at 37°C, 5% CO2. The intermixture was filtrated by a 200 item nylon web to wipe off connective tissues, and the synovial cells were collected by centrifugation for 15 min at 400 g. Finally, fresh DMEM with 15% fetal bovine serum was added to the cells and incubated for 24 h at 37°C, 5% CO2. Then the culture liquid was discarded and the cells were washed, fresh DMEM with 15% fetal bovine serum was added, and the cells continued incubating at 37°C, 5% CO2. The cells were digested by 0.5 g/L trypsin-0.2 g/L EDTA and subcultured.
Histopathology.
The ankle sections (5 μm) fixed in 4% paraformaldehyde were processed following the routine and stained with hematoxilin and eosin (H&E) for histological analysis of the inflammatory reaction, using the optical microscope (Olympus B-50, Japan, Tokyo).
TUNEL Analysis.
For TUNEL analysis, ankle sections (5 μm) fixed in 4% paraformaldehyde were deparaffinized and rehydrated. The tissue was permeabilized with 20 mg/ml of proteinase K for 30 min. Terminal deoxynucleotidyl transferase enzyme and dUTP conjugated to a fluorescein cocktail were added to the tissue sections according to the manufacturer’s specifications (in situ death detection kit; Roche Biochemical, Indianapolis, IN). Nuclei were counterstained with propidium iodide. Slides were mounted with glass coverslips using mounting medium for fluorescence (Kirkegaard and Perry Laboratories, Gaithersburg, MD). Specimens were examined and photographed on a Nikon Eclipse E400 microscope (Nikon, Garden City, NY) equipped for phase contrast and epifluorescence visualization using the digital SPOT camera. The percentage of apoptotic nuclei, as determined by the number of propidium iodide-stained nuclei that were positive on TUNEL staining (n = 3–4 ankles per time point), was calculated by a pathologist who was blinded to the purpose of the study.
Cell Apoptosis Analysis.
The effect of anti-DR5 mAb on cell proliferation was measured using an MTT based assay. Briefly, the cells (5,000/well) were incubated in a 96-well plate (Costar, Cambridge, USA ) in the presence of various concentrations of Anti-DR5 mAb (1.5625, 3.125, 6.25, 12.5, 25, 50 μg/ml) in a final volume of 0.2 ml for 12 h. Thereafter, 0.02 ml of MTT solution (5 g/L) was added to each well and then incubated for 4 h. After centrifugation, the supernatant was removed from each well. The colored formazan crystal produced from MTT was dissolved in 0.15 ml of DMSO and then the optical density (OD) value was measured at 490 nm by a multiscanner autoreader (Dynatech MR 5000, Chantilly, VA). The following formula was used: cell proliferation inhibited (%) = [1-(OD of the experimental samples/OD of the control) × 100%].
DNA fragmentation, serving as a late apoptosis marker, was detected by agarose gel electrophoresis. Cells (1 × 106 cells/ml) at the exponentially growing phase were treated with or without anti-DR5 mAb (50 μg/ml) for 12 h. The cells were washed twice with cold PBS, lysed in 20 ml lysis buffer (50 mM Tris-HCl, pH 7.4; 10 mM EDTA and 0.5% sodium N-lauroylsarcosinate). Lysate was incubated sequentially with 500 mg/ml ribonuclease A and 500 mg/ml proteinase K at 50°C for 60 min in a shaking water bath. Equivalent amounts of DNA (2–3 mg) were then analyzed by 2% agarose gel electrophoresis at 50 V in TBE buffer (2 mM EDTA, pH 8; 89 mM Tris-HCl and 89 mM boric acid). The DNA fragmentation pattern was visualized and photographed under transmission UV light after ethidium bromide staining.
To determine the apoptosis, anti-DR5 mAb-treated cells (25 μg/ml, 12 h) were washed in PBS and resuspended in binding buffer at a concentration of 1 × 106 cells/L. After incubation, 195 μl of the solution was transferred to a 5 ml culture tube with 5 μl annexin V-FITC (BD, USA) added. The tube was then incubated for 30 min at room temperature in the dark. The cells were washed with binding buffer and resuspended in 190 μl binding buffer, with 10 μl PI added. Finally, the tube was gently vortexed and incubated for another 30 min in the dark. Cells were analyzed by the FACSCalibur flow cytometer (Becton Dickinson, USA) with CellQuest software.
RT-PCR Analysis.
Total RNA was isolated using TRIzol reagent. Complementary DNA was synthesized by RT with SuperScriptTM II RNase H-RT (Invitrogen, USA). cDNA was obtained by RT of 1–4 μg of total mRNA, with oligo (dT)18 at 0.5 μg/μl as the primer, and the reaction was done according to the manufacture’s protocol (Invitrogen, USA).
The conditions used were 1 cycle at 95°C for 5 min followed by 30 cycles at 95°C for 50 s, 53°C for 50 s, and 72°C for 1 min. The products were resolved on a 1% agarose gel. Each sample was normalized to GAPDH level.
Western Blot Analysis.
Aliquots (30 μg of protein) of synovial cell lysates were separated on 12% SDS-PAGE, blotted onto a nylon membrane and probed with primary antibodies (Sigma, USA) for 2 h at 4°C. Rabbit anti-caspase-3 (1:1,000 dilution), anti-caspase-8 (1:1,000 dilution), anti-caspase-9 (1:1,000 dilution) antibodies, and mouse anti-bcl-2 (1:1,000 dilution), anti-RANKL (1:1,000 dilution), anti-FHIT (1:1,000 dilution), anti-JNK2 (1:1,000 dilution), anti-c-Jun (1:1,000 dilution), anti-PTEN (1:1,000 dilution) antibodies were from Sigma-Aldrich. Rabbit anti-actin (1:1,000 dilution) antibody was from Santa Cruz Biotechnology. Membranes were washed with 0.05% (vol/vol) Tween 20 in PBS (pH 7.6) and incubated with 1:5,000 dilution of horseradish peroxidase-conjugated secondary Abs (Promega, USA) for 60 min at 4°C. Protein bands were visualized by DAB reaction. Each sample was normalized to β-actin level.
Caspase Inhibition Assay.
Whether anti-DR5 mAb induces synovial cells apoptosis through caspase pathway was further measured using an MTT based assay. Briefly, the cells (5,000/well) were incubated in a 96-well plate (Costar, Cambridge, MA, USA), then added anti-DR5 mAb (IC50 = 12.5 μg/ml) and various concentrations (5, 10, 15, 20, 25, 30, 35 μM) of caspase inhibitor (C0605, Sigma, USA) at the same time in a final volume of 0.2 ml for the indicated times. Subsequent steps were the same as the above.
Statistical Analysis.
Data are presented as the mean ± SD per group. Statistical analysis was made for multiple comparisons using analysis of variance and Student’s t test. A P value < 0.05 was considered to be statistically significant.
Results
Anti-DR5 mAb Can Ameliorate Hind Paw Inflammation and Pathological Changes in AA Rats.
Following an injection of Mycobacterium bovis, the Sprague-Dawley (SD) rats used in this study developed adjuvant arthritis (AA), and it followed the expected clinical course. After 3 days, the paw swelling of AA rats treated with anti-DR5 mAb was less than that of AA rats with no treatment or mouse IgG treatment (P < 0.01) (Fig. 1).
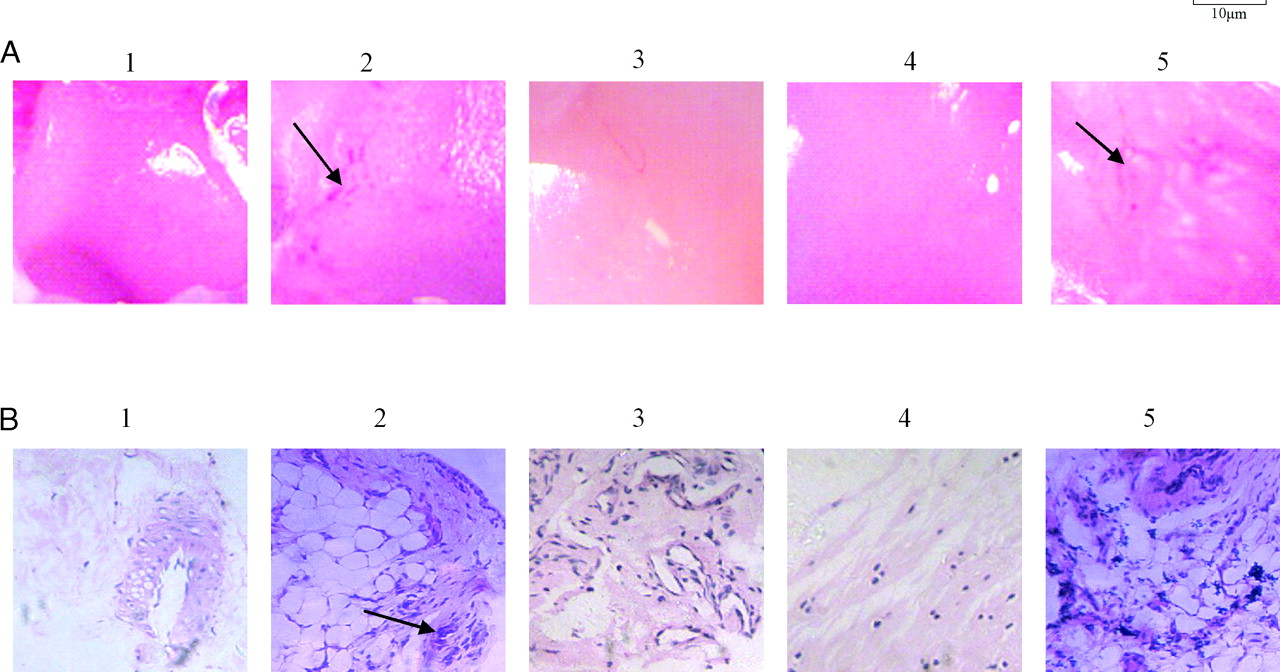
Pathological examination revealed that compared to the normal group (Fig. 2A-1), all AA rats developed severe arthritis characterized by marked hyperplasia of the lining layer cells, neovascularization, erosions of the articular cartilage and bone, and degradation of the internal matrix of the hyaline cartilage due to neovascularization. Compared to the model group (Fig. 2A-2), the AA rats treated with anti-DR5 mAb (Fig. 2A-4) and methotrexate (MTX) (Fig. 2A-3) showed improved joint tissue pathology to some extent. However, the AA rats treated with mouse IgG showed the same pathological changes compared with the model group (Fig. 2A-5).
Figure 2B illustrates the representative staining of synovial tissues from different groups. Hematoxylin and eosin (H&E) staining revealed that unlike the synovial tissues of the normal group, those of the model group and AA rats treated with mouse IgG exhibited typical pathological features of RA, including hyperplasia of the synovium, infiltration of inflammatory cells, and increased vascularity. After treatment with anti-DR5 mAb, hyper-plasia of the synovium decreased and the extent of infiltration of inflammatory cells was reduced, indicating that anti-DR5 mAb can ameliorate the pathological injury due to AA in rats.
Anti-DR5 mAb Can Induce Synovial Cells Apoptosis In Vivo and In Vitro.
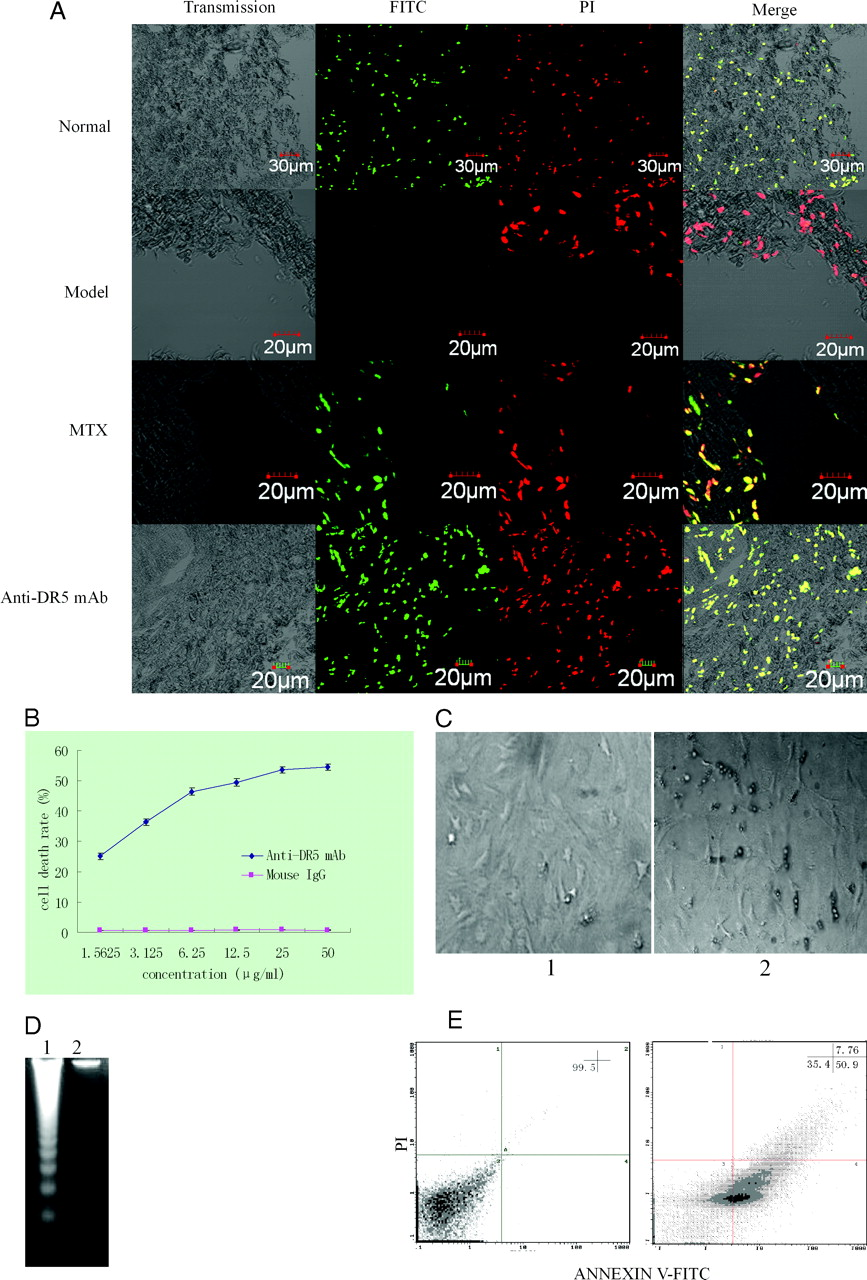
Having observed the pathological changes in AA rats treated with anti-DR5 mAb, we wanted to know whether anti-DR5 mAb induces synovial cell apoptosis in an in vivo system (Fig. 3A). Compared to the normal group, the number of TUNEL+ (yellow, apoptotic) cells was minimal in the model group. However, the number of TUNEL+ (yellow, apoptotic) cells was higher in the AA rats treated with anti-DR5 mAb than in the model group, suggesting anti-DR5 mAb can induce synovial cell apoptosis in an in vivo system.
Subsequently, we studied the in vitro effect of the anti-DR5 mAb. We first used a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT) assay to analyze whether the anti-DR5 mAb could inhibit the in vitro proliferation of the synovial cells of the model group. When the cells were treated with anti-DR5 mAb at 1.5625 μg/ml for 12 h, the cell death rate was 25.1 ± 1.11%, but when the concentration of anti-DR5 mAb was increased to 50 μg/ml, the cell death rate was 54.41 ± 1.15%. This difference in cell death rate indicates dose dependence, suggesting that anti-DR5 mAb can inhibit synovial-cell proliferation (Fig. 3B).
The synovial cells of the model group treated with a high dose anti-DR5 mAb for 12 h were observed under an optical microscope. The cells shrunk and developed bubble-like blebs on their surface. The apoptotic cells broke down into smaller pieces called apoptotic bodies that other body cells recognized and digested (Fig. 3C).
Genomic DNA fragmentation as a hallmark of apoptotic cell death was confirmed using an agarose gel electrophoresis. Unlike the results of agarose gel electrophoresis of the control group, the electrophoresis of the treated cells (treatment with anti-DR5 mAb at 50 μg/ml for 12 h) revealed a typical ladder pattern of internucleosomal DNA fragmentation (Fig. 3D).
We used flow cytometry to study the mode of cell death induced by anti-DR5 mAb. As shown in Figure 3E, in the synovial cells of the model group, the death mode mediated by anti-DR5 mAb was mostly apoptosis and in accord with the results observed with MTT treatment.
Anti-DR5 mAb Can Prolong the Synovial Cell Cycle.
From MTT assay, we concluded that anti-DR5 mAb could inhibit synovial cells growth. So, we next analyzed whether the mechanism was related to cell cycle-related genes (Fig. 4). For this, we first detected the mRNA expression of c-myc that activates the genes regulating the cell cycle machinery (18). Compared with the normal group, c-myc mRNA expression in the model group and mouse IgG treatment group was higher. And the c-myc mRNA expression in the synovial cells of AA rats decreased after treating with anti-DR5 mAb and MTX.
We further analyzed the mRNA expression of p21WAF1/CIP1, which has an important function in negative regulation of the G1 phase check-point. Compared with the normal group, p21WAF1/CIP1 mRNA expression in the model group and mouse IgG treatment group decreased, while compared with the expression in the model group, that in the AA rats treated with anti-DR5 mAb and MTX increased.
p53 is a positive regulator of p21WAF1/CIP1; hence, we also detected the mRNA expression of p53. Compared with the normal group, p53 mRNA expression in the model group and mouse IgG treatment group decreased, while compared with the expression in the model group, that in the AA rats treated with anti-DR5 mAb and MTX increased.
Finally, we studied the mRNA expression of the apoptosis-related genes bcl-2 and bax in different groups. Compared with the normal group, bcl-2 mRNA expression in the model group and mouse IgG treatment group increased, while compared with the expression in the model group, that in the AA rats treated with anti-DR5 mAb and MTX decreased. Contrasting results were obtained for bax.
Induction Apoptosis by Anti-DR5 mAb May Be Through Caspase Pathway.
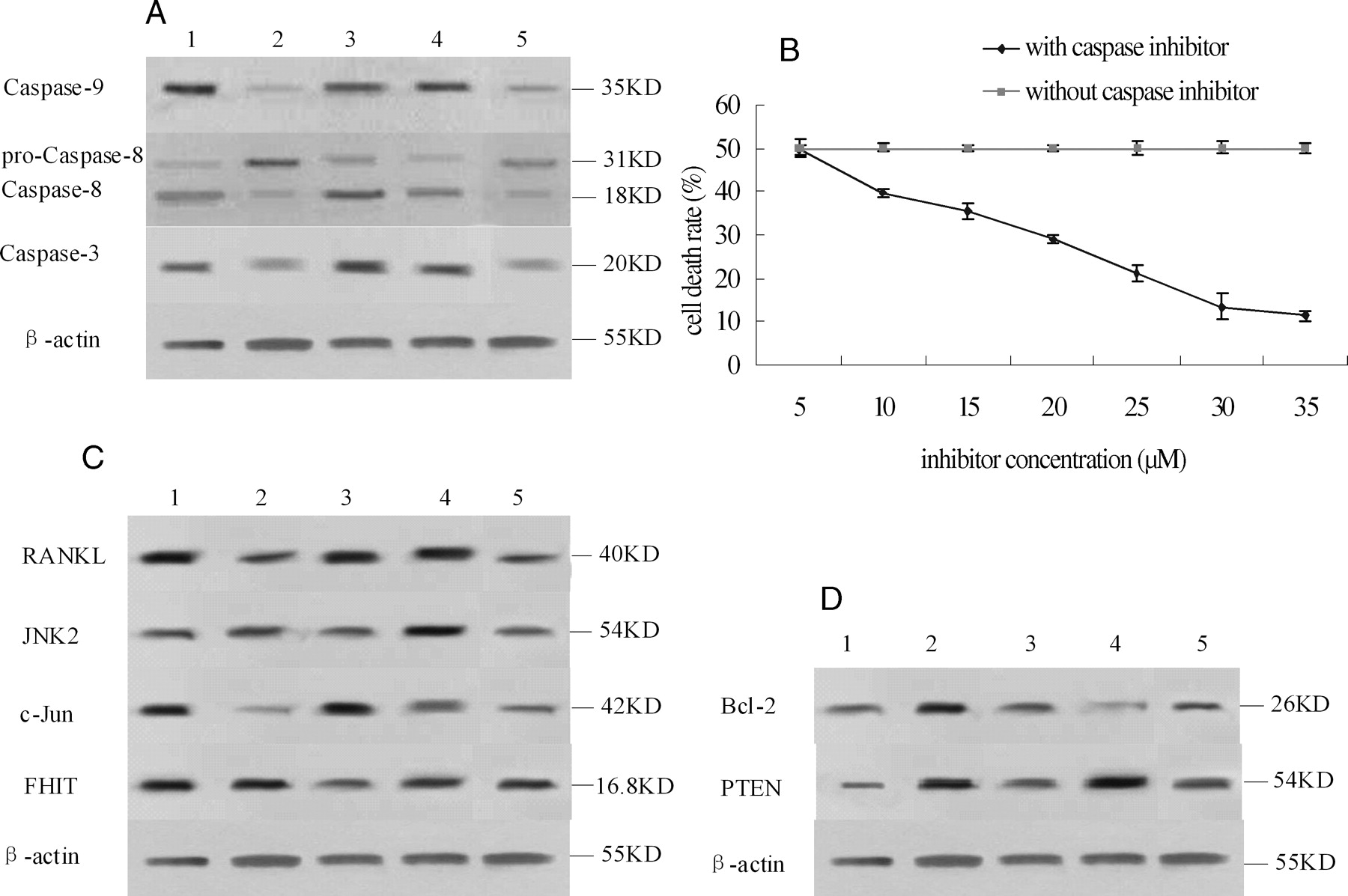
Analysis of the expression of the proteins involved in the apoptosis pathway revealed that the level of caspase-8/3/9 was different in each group (Fig. 5A). Compared with the normal group, the expression of caspase-9/3 in the model group and mouse IgG treatment group were lower, while compared with the expression in the model group, that in the AA rats treated with anti-DR5 mAb was higher. Compared with the normal group, the expression of pro-caspase-8 in the model group and mouse IgG treatment group were higher, but after clipping, the expression of activated caspase-8 was lower. The expression of caspase-8/3/9 in the AA rats treated with MTX was the same as that in the AA rats treated with anti-DR5 mAb.
In order to prove that anti-DR5 mAb induces apoptosis via the caspase pathway, we added various concentrations (5, 10, 15, 20, 25, 30, 35 μM) of a caspase inhibitor to synovial cells treated with anti-DR5 mAb. The results showed that the caspase inhibitor could inhibit the apoptosis induced by anti-DR5 mAb. The inhibition effect is dose dependent and was maximum when the concentration of inhibitor increased, the induction of synovial cell apoptosis by anti-DR5 mAb might be through the caspase pathway (Fig. 5B).
Other Factors Involved in Anti-DR5 mAb Treated Synovial Cells.
In order to study other factors involved in this treatment, we further analyzed the expression of c-Jun, receptor activator of nuclear factor (NF)-κ B ligand (RANKL), and JNK2.
Compared with the normal group, the expression of c-Jun, and RANKL in the model group and mouse IgG treatment group was lower, but the expression of JNK2 and fragile histidine triad (FHIT) was not significantly different. Compared with the model group, the expression of c-Jun, RANKL, and JNK2 in the AA rats treated with anti-DR5 mAb was higher; the expression of FHIT was not significantly different; in AA rats treated with MTX, the expressions of c-Jun and RANKL were higher, while the expressions of JNK2 and FHIT were lower (Fig. 5C).
We also detected the protein expression of bcl-2, phosphate and tensin homolog (PTEN) on synovial membrane cells simultaneously. Compared with the normal group, the expression of bcl-2 in the model group and mouse IgG treatment group was higher; while compared to the expression in the model group, that in the AA rats treated with anti-DR5 mAb was lower, the protein expression of PTEN was not significantly different, and the expressions of bcl-2 and PTEN were lower in the AA rats treated with MTX (Fig. 5D).
Discussion
After inflammation for 30 days, the AA rats showed synovial membrane incrassation and fibrosis, hyperplasia of the lining layer cells, pannus formation, erosions of articular cartilage and bone, and degradation of the internal matrix of the hyaline cartilage due to neovascularization. Our results revealed that after inflammation on the 15th day, a secondary inflammation develops and reaches a maximum at 24–27 days. These pathological changes were consistent with former studies (19). An intravenous injection (1 ml·rat−1·d−1) of anti-DR5 mAb on the 3rd day has a great effect on AA rats even if the treatment is stopped. After inflammation for 30 days, when detecting the swelling paw there is a prominent statistical difference compared with the model group, suggesting that anti-DR5 mAb treatment could ameliorate arthritis symptoms. One-third of the rats from each group were sacrificed at days 30, 37, and 44, and we did not observe a considerable difference among the results (data not shown), indicating that the treatment with anti-DR5 mAb for 15 days did not cause other effects.
Having observed that pathological changes occur in AA rats treated with anti-DR5 mAb, we wanted to know whether anti-DR5 mAb induces synovial cell apoptosis in an in vivo system. Comparing with the normal group, the number of TUNEL+ (yellow, apoptotic) cells was minimal in the model group. However, the number of TUNEL+ (yellow, apoptotic) cells was higher in the AA rats treated with anti-DR5 mAb than in the model group, suggesting that anti-DR5 mAb can induce synovial cell apoptosis in an in vivo system.
Although the hyperproliferating synovial cells are nonmalignant, they have been described as “transformed-appearing synoviocytes” because they share some common features with transformed cells, including a dense rough endoplasmic reticulum, numerous irregular nuclei, and changes in the spindle-shaped normal cell skeleton (20). Therefore, we examined the antiproliferative activity of anti-DR5 mAb against synovial cells. Evidently, the antiproliferative effects were gradually augmented with the increased concentration. The results suggested that this activity may be due to the direct toxicity of the antibody against synovial cells.
Next, whether the apoptotic pathway is involved in the cell death caused by anti-DR5 mAb was investigated. Several lines of evidence presented in our study indicate that apoptosis is the mode of cell death caused by this antibody complex. Morphological changes were clearly observed after 12 h. Consistent with this observation, the nuclear fragments, visualized as a ladder on electrophoresis, were also evident. The result of cell apoptosis analysis assessed by flow cytometry indicated that the apoptosis is the main mode of death of synovial membrane cells treated with anti-DR5 mAb.
We studied the molecular mechanism involved in the apoptosis induced by anti-DR5 mAb. Three mechanisms are known to be involved in the apoptotic process: a death receptor pathway, a mitochondria pathway, and a mechanism in which the endoplasmic reticulum plays a central role (21). Our observation revealed that anti-DR5 mAb-treated synovial cells activate caspase-9 and increase the expression of bax and decrease the expression of bcl-2, and then induce mitochondria to release cytochrome c. On the other hand, these cells activate caspase-8 by anti-DR5 mAb binding with DR5, resulting in the formation of a trimer and by recruiting Fas-associated protein with death domain (FADD). Ultimately, caspase-8 and caspase-9 activate caspase-3, suggesting that anti-DR5 mAb induced synovial membrane cell apoptosis through the death receptor and the mitochondria and finally via activation of the caspase pathway. The caspase inhibition assay further supports this viewpoint.
Further, we studied the factors that regulate the main apoptosis pathway. As we know, abnormal or ectopic overexpression of c-myc in primary cells activates a protective pathway through the induction of p19/p14ARF and a p53-dependent cell death pathway (22). First, we analyzed the mRNA expression of cell cycle-related genes such as c-myc, p53, and p21. Our results indicate that anti-DR5 mAb treatment of synovial cells appears to decrease the probability of the entry of these cells into the cell cycle through the decreased mRNA expression of c-myc and increased mRNA expression of p53 and p21.
In this study, we also explored the other regulators involved. Secchiero P et al. (23) have reported that TRAIL could regulate normal erythroid maturation through an ERK-dependent pathway but not through the p38/mitogen-activated protein kinase (MAPK) or JNK pathways. However, our results showed that anti-DR5 mAb treatment increased the protein expression of RANKL, c-Jun, and JNK2 in synovial cells, indicating that the cells could activate the JNK pathway. Activation of this pathway was probably to activate the expression of death receptors, such as DR5, that would further enhance apoptosis. That is why even after termination of the anti-DR5 mAb treatment, the therapeutic effects persisted.
In many human cancers, the FHIT is also impaired and its induction of apoptosis seems to be FADD dependent, caspase-8 mediated, and independent of mitochondrial amplification (24). The tumor suppressor PTEN functions as a phosphoinositide 3-phosphatase; its activity is antagonistic to that of phosphatidylinositol 3-kinase, and it negatively regulates cell proliferation and survival signals (25). In our studies, the level of expression of the FHIT and PTEN proteins was not significantly different in AA rats regardless of therapy. This indicated that the effects of anti-DR5 mAb treatment in AA rats were not mediated by FHIT or PTEN pathway.
In conclusion, our results demonstrate that functionally, the activity of anti-DR5 mAb on synovial cells is almost as effective as a cure. Our study is the first to suggest that anti-DR5 mAb could probably be considered as a novel therapeutical candidate for the treatment of RA, based on evidence from experiments in in vitro and in vivo systems. Further studies on T cells, B cells, and monocytes/macrophages involved in immune-mediated inflammation in RA are currently being pursued to determine the selective cytotoxicity of anti-DR5 mAb. Thus, a DR5-specific strategy may be effective not only as an anti-cancer therapy, but also as an anti-inflammatory therapy for RA.
Effect of anti-DR5 mAb on the swelling ankle of rat adjuvant arthritis (x ± s, n = 12). Normal: Rats injected with 0.1 ml of 0.85 NaCl; Model: Rats received a single injection of 0.1 ml of Freund’s complete adjuvant; MTX treatment: Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with MTX 1.0 ml·rat−1·week−1 for 15 days; Anti-DR5 mAb treatment: Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with anti-DR5 mAb 1.0 ml·rat−1·d−1 for 15 days; Mouse IgG treatment: Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with mouse IgG 1.0 ml·rat−1·d−1 for 15 days. Effect on pathological changes in ankle of AA rats by anti-DR5 mAb. A, Pathological changes of AA rats were observed by a pathologist who was blinded to the purpose of the study (×20). 1 (normal): Rats injected with 0.1 ml of 0.85 NaCl; 2 (model): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant; 3 (MTX treatment): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with MTX 1.0 ml·rat−1·week−1 for 15 days; 4 (anti-DR5 mAb treatment): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with anti-DR5 mAb 1.0 ml·rat−1·d−1 for 15 days; 5 (mouse IgG treatment): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with mouse IgG 1.0 ml·rat−1·d−1 for 15 days. B, H&E (synovial inflammation, original magnification, ×40)-stained representative joint sections from different animal groups (photos were taken before the rats were sacrificed on the 30th day). Arrows point to inflammatory cells. Anti-DR5 mAb induce synovial cells apoptosis in vivo and in vitro. A, TUNEL assay. Ankle sections (5 μm) fixed in 4% paraformaldehyde were deparaffinized and rehydrated, then following the manufacturers’ instructions. At last, cells were mounted in PI and visualized. Green, FITC labeled dUTP showing apoptosis; Red, PI stained nucleolus; Yellow, merged staining showing TUNEL+ cells. B, Cytotoxic effects of anti-DR5 mAb (concentration from 1.5625 μg/ml–50 μg/ml) on synovial cells of model group. C, Synovial cell of model group was observed under optical microscope. 1: synovial cell of model group with mouse IgG treatment; 2: synovial cell of model group treated with 50 μg/ml anti-DR5 mAb for 12 h. D, Cells were cultured with 50 μg/ml anti-DR5 mAb for 12 h, and the DNA fragmentation was analyzed by agarose gel electrophoresis. 1: synovial cells of model group treated with anti-DR5 mAb; 2: synovial cells of model group with mouse IgG treatment. E, Flow cytometric analysis of death mode of synovial cells of model group treated by anti-DR5 mAb (25 μg/ml +12 h) after staining by ANNEXIN V FITC/PI. The results showed that the cell death mode mediated by anti-DR5 mAb is mostly apoptosis. Left: control, right: synovial cells of Model group treated with anti-DR5 mAb. mRNA expression of bax, bcl-2, c-myc, p53, and p21 in synovial cells. Anti-DR5 mAb could increase the expression of p21, p53, and bax and decrease the expression of c-myc and bcl-2 comparing with model group. 1 (normal): Rats injected with 0.1 ml of 0.85 NaCl; 2 (model): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant; 3 (MTX treatment): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with MTX 1.0 ml·rat−1·week−1 for 15 days; 4 (anti-DR5 mAb treatment): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with Anti-DR5 mAb 1.0 ml·rat−1·d−1 for 15 days; 5 (mouse IgG treatment): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with mouse IgG 1.0 ml·rat−1·d−1 for 15 days. Mechanism on the treatment by anti-DR5 mAb. A, Western blot analysis of Caspase-8/3/9. Analysis of the expression of the proteins involved in the apoptosis pathway revealed that anti-DR5 mAb could activate the level of Caspase-8/3/9 compared with model group. B, Caspase inhibition assay. The cells treated with anti-DR5 mAb (IC50) were added with various concentrations of caspase inhibitor from 5 μM to 35 μM for 12 h. The inhibition effects of anti-DR5 mAb on synovial cells were taken in a dose-dependent manner. C, Western blot analysis of c-Jun, RANKL, JNK2, and FHIT. Anti-DR5 mAb could increase the expression of c-Jun, RANKL, and JNK2 compared with model group, but the expression of FHIT has no significant difference. D, Western blot analysis of Bcl-2, PTEN. Anti-DR5 mAb could decrease the expression of Bcl-2 compared with model group; while the protein expression of PTEN has no significant difference. 1 (normal): Rats injected with 0.1 ml of 0.85 NaCl; 2 (model): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant; 3 (MTX treatment): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with MTX 1.0 ml·rat−1·week−1 for 15 days; 4 (anti-DR5 mAb treatment): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with Anti-DR5 mAb 1.0 ml·rat−1·d−1 for 15 days; 5 (mouse IgG treatment): Rats received a single injection of 0.1 ml of Freund’s complete adjuvant, followed by injection with mouse IgG 1.0 ml·rat−1·d−1 for 15 days.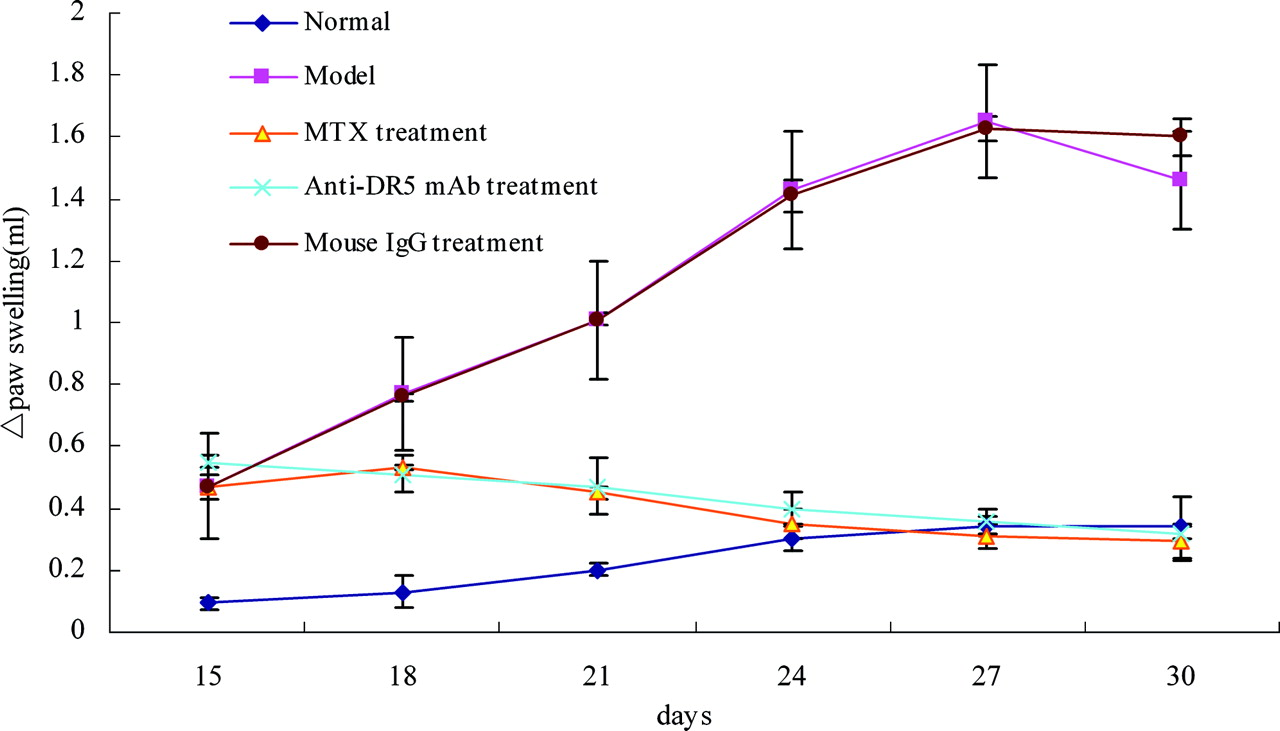

Footnotes
1
These authors contributed equally to this work.
Supported by the Scientific Research Foundation of Fujian Province (No. 2007J0116).