
Abstract
Nutrition during pregnancy and lactation can program an offspring’s metabolism with regard to glucose and lipid homeostasis. A suboptimal environment during fetal, neonatal and infant development is associated with impaired glucose tolerance, type 2 diabetes and insulin resistance in later adult life. However, studies on the effects of a low protein diet imposed from the beginning of gestation until adulthood are scarce. This study’s objective was to investigate the effects of a low protein diet imposed from the gestational period until 4 months of age on the parameters of glucose tolerance and insulin responsiveness in Wistar rats. The rats were divided into a low protein diet group and a control group and received a diet with either 7% or 25% protein, respectively. After birth, the rats received the same diet as their mothers, until 4 months of age. In the low protein diet group it was observed that: (i) the hepatic glycogen concentration and hepatic glycogen synthesis from glycerol were significantly greater than in the control group; (ii) the disposal of 2-deoxyglucose in soleum skeletal muscle slices was 29.8% higher than in the control group; (iii) there was both a higher glucose tolerance in the glucose tolerance test; and (iv) a higher insulin responsiveness in than in the control group. The results suggest that the low protein diet animals show higher glucose tolerance and insulin responsiveness relative to normally nourished rats. These findings were supported by the higher hepatic glycogen synthesis and the higher disposal of 2-deoxyglucose in soleum skeletal muscle found in the low protein diet rats.
Introduction
Nutrition during pregnancy and lactation can program an offspring’s metabolism, including their glucose and lipid homeostasis (1). Protein-energy malnutrition is the most prevalent form of nutritional disorder in developing countries. Epidemiological (2) and laboratory studies (3) indicate that a suboptimal environment during fetal, neonatal and infant development is associated with the development of impaired glucose tolerance, type 2 diabetes mellitus (T2DM) and insulin resistance in later adult life. These studies have revealed a strong inverse relationship between birth weight and the risk of developing T2DM and metabolic syndrome (3, 4). The “Thrifty Phenotype Hypothesis” proposes that low birth weight individuals are characterized by early adaptations to a potentially adverse intra-uterine environment and that these adaptations optimize the use of a restricted nutrient supply to ensure survival (5, 6).
The term ‘programming’ is employed to denote the molecular mechanisms underlying the insult when it occurs during a critical or sensitive period of development, and it results in long-term or permanent changes in the structure or function of an organ or metabolic action that causes the individuals to become more susceptible to subsequent metabolic diseases (6, 7). The mechanistic basis of this relationship remains the subject of research and debate. Energy conservation directed at accelerating body fat recovery (or catch-up fat) contributes to obesity relapse after slimming and to excess fat gain during catch-up growth after malnutrition (8). A thrifty metabolism promotes the preferential accumulation of catch-up fat, which is associated with insulin resistance, during catch-up growth after earlier periods of faltered growth (9). Additionally, a thrifty metabolism has been implicated in the link between earlier growth retardation and catch-up growth and increased risks of type 2 diabetes and cardiovascular disease later in life (10).
The maternal protein restriction rat model (i.e., low-protein or LP) has been used with undernutrition imposed during various developmental phases in early life. Studies were carried out on some tissues in an effort to clarify the molecules and events are involved in this process. It was determined that the impact of maternal protein restriction on metabolic programming is most pronounced in insulin-sensitive tissues. Snoeck et al. (11) showed that fetal malnutrition in rat pups results in a reduced number of β cells, in a diminution in the proliferation of islet cells, in a reduced islet size and in a marked decrease in islet vascularization. Latorraca et al. (12) and Martins and Sawaya (13) have suggested that glucose homeostasis is maintained in LP rats and nutritionally stunted children. Increased responsiveness to glucose was observed in rats as a result of alterations in the initial steps of the insulin signal transduction pathways. Adult offspring, whose mothers were submitted to protein restriction during lactation, support the hypothesis that hypermetabolic status programming occurs (14). In a rat model of protein deficiency during pregnancy and lactation, an altered insulin response in the liver was demonstrated in offspring (15). During adult life (weeks 6–12 of life), LP offspring have better glucose tolerance than controls (16–18). However, there are age-dependent changes, such that by the 44th week of life the glucose tolerance of LP offspring is similar to (19), and by 15 months of age significantly worse than, that of controls (20). Female rats that received an LP diet had a more delayed insulin resistance than male rats (21), demonstrating a different profile for age and sex in the development of T2DM.
At 3 months of age, adipocytes from LP offspring present increased basal and insulin-stimulated glucose uptake relative to controls (22). This may be related to a three-fold increase in the number of insulin receptors in LP adipocytes. Indeed, adipocytes from LP animals had significantly higher basal and insulin-stimulated insulin receptor substrate (IRS)-1-associated phosphatidylinositol 3-kinase (PI 3-kinase) activities. There was also an increase in p85-associated PI 3-kinase activity in these adipocytes, as well as a reduction in the number of p110-beta catalytic subunits of the PI 3-kinase in adipocytes from LP animals (22). In freshly isolated muscle strips from 3-month-old rats fed with a control diet (20% casein) and rats that received a diet containing 8% protein during the gestational and lactational period, it was observed that the muscle strips from LP animals (preincubated with insulin at 300 pM) took up more [3H]methylglucose than the controls, thus showing an augmented insulin sensitivity (23). With respect to studies of an LP diet’s effects on parameters of insulin resistance and/or metabolic syndrome, most studies have been conducted using an LP diet during gestation, lactation, or gestation and lactation, but after the lactation period the animals are returned to a normal and low fat diet.
The metabolic programming that results from the LP diet during pregnancy and lactation certainly persists after the lactation period. The objective of this study was to examine the effects of a LP diet during pregnancy, lactation and on into adulthood so we cannot separate the effects from each stage (gestational, lactational and post-lactational). However, it is known that the main effects of metabolic programming occur during pregnancy and lactation and that the stage of pregnancy influences the metabolic programming’s effects (24).
Sugden and Holness (25) showed that rats that were submitted to a protein-restrict diet during pregnancy, lactation and continued with low protein diet until the age of 20 weeks have an increased insulin sensitivity compared to control groups (20% protein) and to the group subjected to low protein diet during pregnancy. This study showed that exposure to protein restriction during the early life alone, leads to relative insulin resistance and hyperinsulinaemia in adulthood, but this relationship is gender specific, observed only in males. It is appropriate to investigate the effects of an LP diet on rats from gestation until 4 months after birth, with respect to glucose tolerance and insulin responsiveness, muscle glucose uptake, as well as hepatic glycogen synthesis and concentration.
Methods and Materials
Chemicals.
2-[U-14C]-Deoxy-D-Glucose (2-DG) (specific activity 308 mCi/mmol), D-[U-14C]-Glycerol (specific activity 157 mCi/mmol) and D-[U-14C]-Glucose (specific activity 297 mCi/mmol) were obtained from Amersham International (Buckinghamshire, UK). Glycogen was obtained from Sigma Chemical Co. (St. Louis, MO, USA). All Other chemicals used were obtained from Merck S.A., Porto Alegre, Brazil. Diagnostic kits were obtained from Labtest Diagnóstica (Lagoa Santa, MG, Brazil).
Animals and Undernutrition Model.
Female Wistar rats from our breeding colony were used. They were maintained at 22°C, on a 12 h light/12 h dark cycle until experimental age. The females were allowed to mate with males, and day 0 of gestation was taken as the day in which the vaginal plugs were expelled. The litter size was adjusted to eight pups per mother on the first postpartum day. The protocols conformed to the guidelines of the Committee on Care and Use of Experimental Animal Resources, School of Veterinary Medicine and Animal Science of the University of São Paulo, Brazil.
Gestational and lactational undernutrition was induced in pups by restricting the mother’s diet to a 7% protein content (control group: 25% protein). All animals were fed ad libitum from the pregnancy’s beginning and during the entire gestational and lactational period, during which the undernutrition was produced by decreasing the global amount of nutrients accessible to the pups. At 21 days postnatal, the pups were weaned, and from that point until the experimental age (16 weeks), additional undernutrition was induced by maintaining the same 7% protein diet. Both groups were sacrificed by decapitation to collect blood and adipose tissue. Additionally, the liver and soleum muscle were rapidly removed and weighed.
Diet.
An isocaloric diet containing 25% or 7% protein, salts and vitamins as recommended by the Association of Official Analytical Chemists (26), and as previously described by our group (27, 28), were used (Table 1). The animals’ weights were measured at birth, 7, 30, 60, 90 and 120 days.
Sample Collection.
After 16 weeks of treatment, all rats were sacrificed by decapitation, and blood samples were collected in test tubes and immediately centrifuged at 3000 × g for 10 min to obtain serum samples.
Insulin Responsiveness Test (IRT).
An intra-peritoneal IRT was performed at 16 weeks of age in low protein diet and control female rats. Before the test, the rats were fasted overnight (8 h). Subsequently, they were injected intraperitoneally with an insulin solution (Humulin; Lilly France S.A., Fergesheim, France) at a dose of 0.75 U/kg body weight. Blood samples were obtained from the cut tip of the tail immediately before and 30, 60 and 120 min after the injection. The glycemia was determined using a glucosimeter (Accu-Chek® Advantage; Roche Diagnostics, Mannheim, Germany).
Glucose Tolerance Test (GTT).
An intraperitoneal GTT was performed at 16 weeks of age in low protein diet and control female rats. The rats were fasted overnight (8 h) before the test. They were then injected intraperitoneally with a 40% (wt/vol) glucose solution at a dose of 2 g glucose/kg body weight. Blood samples were obtained from the cut tip of the tail immediately before and 30, 60 and 120 min after the injection. The glycemia was determined using a glucosimeter (Accu-Chek® Advantage; Roche Diagnostics, Mannheim, Germany).
In all other experiments, the female rats were submitted to a 6-fast, and the animals were killed between 14 and 15 h after the start of the experiment.
Glucose Incorporation from L-[U-14C]-Glycerol to Liver Glycogen.
After decapitation, the liver was removed, and between 80–100 mg of hepatic tissue was immediately cut into 0.3-mm slices using a McIlwain tissue chopper. The tissue slices were incubated in 1.0 mL Krebs-Ringer bicarbonate buffer (KRb) at pH 7.4, containing 5 mM glucose + 1 mM glycerol + 0.2 μCi L-[U-14C]-Glycerol. Incubations were carried out in flasks after the contents were sparged with a 95% O2:5% CO2 mixture for 1.0 min. The slices of liver were incubated for 1 h at 35°C in a Dubnoff metabolic shaker (60 cycles/min) according to the method of Dunlop et al. (29). The reaction was stopped by putting the flasks on ice (4°C) and adding 1.0 mL KOH (60%). The flasks’ contents were transferred to tubes, and the flasks were washed with 0.5 mL KOH (30%). Afterwards, the tubes were heated to 100°C for 20 min or until the tissue was completely homogenized. After cooling, 3.8 mL ethanol 99°G.L. was added, and the tubes were warmed at 70°C for 10 min and then put on ice for 15 min (until glycogen precipitation). After centrifugation (2000 × g for 10 min), the supernatant was rejected, and the pellet was suspended in 0.3 mL of Milli Q water and transferred to vials. The vials were charged with 1.7 mL of ethanol 95°G.L., and iced for 20 min. The material was centrifuged at 2000 × g for 10 min, and the supernatant was again discarded (30). Then 0.2 mL of water and scintillation liquid were added to the pellet. The radioactivity incorporation was measured and represents the glucose incorporation from the glycogen precursors studied. All results are expressed relative to the initial specific activity of the incubation system.
Hepatic Glycogen Concentration.
The hepatic glycogen concentration was measured using the Krisman method (30).
Uptake of 2-Deoxy-Glucose in Soleum Skeletal Muscle.
Strips of soleum muscle weighing about 45 mg (40–50 mg) were incubated at 37°C with gentle agitation for 30 min in KRb at pH 7.4 that was previously oxygenated (95% O2:5% CO2 mixture for 1.0 min) and contained 5 mM glucose and 0.1 μCi 2-[U-14C]-Deoxy-D-Glucose. The medium was sparged with 95% O2:5% CO2 throughout the incubation to prevent tissue hypoxia. After incubation, the reaction was stopped by putting the flasks on ice (4°C). The incubation system was removed, and the slices were washed 3 times with 1.0 mL KRb. The slices and the washed solution were transferred to tubes and centrifuged 2000 × g for 10 min. The supernatant was removed, and 0.2 mL of a 2 N NaOH solution was added to the pellet. After the tissue homogenization, the scintillation liquid was added, and the radioactivity incorporation was measured in a Wallac scintillation counter (Turku, Finland). The radioactivity represents the uptake of 2-deoxy-glucose in the studied tissue.
Statistical Analyses.
All calculations were analyzed using a Student’s t test. The data in the text are presented as the mean ± SEM (range). Significance levels were set to P < 0.05 and P < 0.001. All analyses were done with SPSS 10.0 (SPSS, Chicago, USA).
Results
Exposure to the protein-restricted diet from the beginning of fetal life through suckling and until 16 weeks of age resulted in an important reduction in the offsprings’ body weight, with a ~30% corporal weight deficiency in the LP diet fed rats group (compared to the control group) at the seventh postnatal day and 70% and 66% deficiencies at 30 and 120 days postnatal, respectively (Table 2). Similarly, the undernutrition resulted in a reduction in feeding from weaning until 90 days of age, with a 72% increase in feeding between the 60th and 90th day in the low protein diet rats.
The ratio of liver weight/100 g corporal weight for the LP diet fed rats was significantly greater than that of the control group, while the ratio of the retroperitoneal and visceral adipose tissue weight/100 g corporal weight for the low protein diet rats was significantly lower than that observed in the control group (Table 3).
To observe the effects of LP diet on insulin responsiveness parameters, we performed the Insulin Responsiveness Test (IRT) and the Glucose Tolerance Test (GTT). In the GTT (Fig. 1), the blood glucose concentration of the LP diet fed rats was significantly lower than that of the control group at 30, 60 and 120 min after glucose i.p. injection (2.0 g/kg rats). The LP diet fed rats demonstrated a decreasing glycemia deficiency as time went by (31%, 21% and 19% for the 30, 60 and 120 min time points, respectively).
The IRT showed that the glycemia of the LP diet fed rats was significantly lower than that of the control group at 30 (33%) and 60 (22%) min after i.p. insulin injection. This is related to better insulin responsiveness in the low protein diet rats as young adults (Fig. 2). These results demonstrate better glucose tolerance in low protein fed rats in young adult life. In addition, at 16 weeks the serum glucose levels of both groups of rats were similar (110.5 ± 3.0 mg % and 108.5 ± 3.8 mg % for the control and low protein diet rats, respectively). The rats in the other experiments were also used for the TTG and IRT tests. The IRT and TTG tests were performed 4 and 2 days, respectively, before the tissue experiments.
Considering the increased responsiveness to insulin and the glucose tolerance in the LP diet adult female rats, we endeavored to observe undernutrition’s effects on the liver, skeletal muscle and adipose tissue parameters. The LP diet fed group had hepatic glycogen concentrations and levels of hepatic glycogen synthesis from glycerol that were significantly greater than in the control group (165% and 70%, respectively; Table 4). Strips of soleum skeletal muscle from low protein diet rats showed higher glucose uptake (29.8%), as compared to the control group (Table 5).
Discussion
Impaired fetal growth has been associated with an increased risk of developing metabolic syndrome (31). The “Thrifty Phenotype Hypothesis” proposes a metabolic adaptation more beneficial if the poor conditions continue, but if the postnatal environment provides plentiful nutrition, these individuals will be at increased risk of developing conditions like obesity and T2DM as a result of this programming conflict (5).
Since there were few reports in the literature studying undernutrition from gestation until adulthood, our discussion is based on gestational, lactational and post-lactational undernutrition. However, the “Thrifty Phenotype Hypothesis” does not exclude the possibility that the malnutrition extends until adulthood, which is what typically happens with malnutrition in humans.
In this study, we investigated some effects of protein undernutrition imposed during the gestational, lactational and post-lactational (until 4 months after birth) periods on some periphery insulin resistance indicator parameters. On the first postnatal day, the low protein diet female rats (protein: 7%) showed a diminution of corporal weight greater than 20% when compared to the control group (protein: 25%), which indicates a conspicuous effect of gestational undernutrition. At 4 months of age, the weights of the LP diet fed rats were on average 39.5% of that observed in the control rats. The permanent reduction in body growth (Table 2) was accompanied by a reduction in food intake (Table 2) until 90 days of age. However, when the consumption of food is expressed on the basis of body weight, the 90-day-old low protein diet rats consumed double the food of the control rats. However, at 4 months of age, the corporal weight of rats fed with LP diet is only 39.5% of the control group. This data indicates that the effects of the LP diet persist despite the increased food consumption.
Table 3 shows that the LP female rats display a higher ratio of hepatic weight/100 g corporal weight, suggesting a more pronounced effect of the malnourishment on the corporal weight than on the hepatic weight. The control rats presented a ratio of retroperitoneal and visceral fat tissue/100 g corporal weight that was 84% higher than in low protein diet rats. Although in absolute terms most organs are reduced in weight in the LP offspring, when expressed relative to body weight, it is apparent that growth restriction is not uniform throughout all tissues (36). Clearly, the diminished retroperitoneal and visceral fat mass is related to the lower body weight of the adult LP offspring.
At 3 months of age, adipocytes from LP offspring have an elevated basal and insulin-stimulated glucose uptake and express more insulin receptors than the controls (37). Conversely, these animals have increased basal and insulin-stimulated insulin receptor substrate (IRS)-1-associated phosphatidylinositol 3-kinase activity. PI 3-kinase is essential for the mediation of several metabolic insulin actions, including the stimulation of glucose uptake and the inhibition of lipolysis (22). In addition, the LP offspring are resistant to the antilipolytic action of insulin (38, 39). At 3 months, the LP adipocytes are also more sensitive to the action of catecholamines to stimulate lipolysis (39), which is related to an increased expression of β-adrenergic receptors (40). The decreased quantity of adipose tissue in the low protein diet rats probably contributes to the higher insulin sensitivity in these animals (41, 42).
Figures 1 and 2 show that LP diet fed rats have a greater insulin responsiveness, as evidenced by the glucose tolerance test and insulin responsiveness test, which corroborate the “Thrifty Phenotype Hypothesis.” With an experimental model of maternal protein restriction imposed during lactation, Fagundes et al. (14) demonstrated that LP animals present lower serum insulin levels and slightly lower glycemia than control groups. Indeed, a lower HOMA index was observed, which may indicate that despite hypoinsulinemia, there was normal or even greater than normal insulin sensitivity, since a higher HOMA index indicates insulin resistance (43).
Insulin-sensitive tissues undergo extensive, programmed changes in response to maternal protein restriction. The hepatic glycogen concentration was 2.6 times higher, and hepatic glycogen synthesis from glycerol was 70% higher in low protein diet rats as compared to control rats (Table 4). An elevated hepatic glycogen concentration is an indicator of high insulin sensitivity, and considering that there was no difference in glycemia between the studied groups at time zero in the glucose tolerance test (Fig. 1), the increase in the hepatic glycogen concentration in the low protein diet group cannot be attributed to a low mobilization of hepatic glycogen. Ex vivo perfusion studies have shown that livers from adult male LP offspring (3 months old) are resistant to the stimulatory action of glucagon on hepatic glucose output (44). This change is accompanied by an 80% reduction in the expression of hepatic glucagon receptors. Membrane receptors appear to be a common target of such metabolic programming, with hepatic insulin receptors being up-regulated in response to maternal protein restriction (44).
After feeding, the glycogen synthesis occurs mainly indirectly. In the low protein diet group, the increases in the hepatic glycogen concentration and glycogen synthesis (Table 4), as well as the higher 2-deoxy-D-glucose uptake by soleum skeletal muscle in 16-week-old female rats (Table 5), corroborate the “Thrifty Phenotype Hypothesis” (45). Muscle is considered to be the major site of glucose uptake postprandially (46) and has therefore been an important research focus. Freshly isolated tibialis anterior muscle strips from low-protein animals that were preincu-bated with 300 pM insulin took up more [3H]methylglucose than similarly prepared samples from control animals (23). Thus, the increase in insulin sensitivity can contribute to glucose uptake in the basal state in low protein diet rats. However, the possibility that other factors could also contribute to the difference in basal glucose transport cannot be excluded. The increase in GLUT4 concentration in muscle plasma membranes from LP animals, as compared to control animals, is an important factor because there are twice the number of insulin receptors in skeletal muscle plasma membranes from LP rats/mg of protein, and the receptors in the LP rats have the same affinity as the receptors from the skeletal muscle plasma membranes of the control group (23). These results reinforce our data from the glucose uptake and insulin sensitivity tests, since at an insulin concentration of 16 nM, we observe the same response in both animal groups.
Conclusion
This study shows that protein undernutrition imposed from the gestational period until 4 months after birth induces a reduced adipose mass in the low protein diet animals and higher insulin responsiveness and glucose tolerance, which may be related to increased hepatic glycogen concentration and hepatic glycogen synthesis, as well as the higher disposal of glucose in skeletal muscle slices. The pathological consequences of chronic under-nutrition are at least in part due to endocrine abnormalities related to altered biological insulin action.
Nutritional Composition of the Diets (g/100 g) a
Corporal Weight and Food Intake of Rats Submitted to Different Diets a
Effects of Low Protein Diet on Liver Weight/100 g Corporal Weight and Total Adipose Tissue Weight/100 g Corporal Weight Ratio in 16-Week-Old Female Rats a
Effects of Low Protein Diet on Hepatic Glycogen Concentration and Hepatic Glycogen Synthesis from L-[U-14C]-Glycerol in 16-Week-Old Female Rats a
Effects of Low Protein Diet on Uptake of 2-[U-14C]-Deoxy-D-Glucose (2-DG) by Soleum Skeletal Muscle in 16-Week-Old Female Rats a
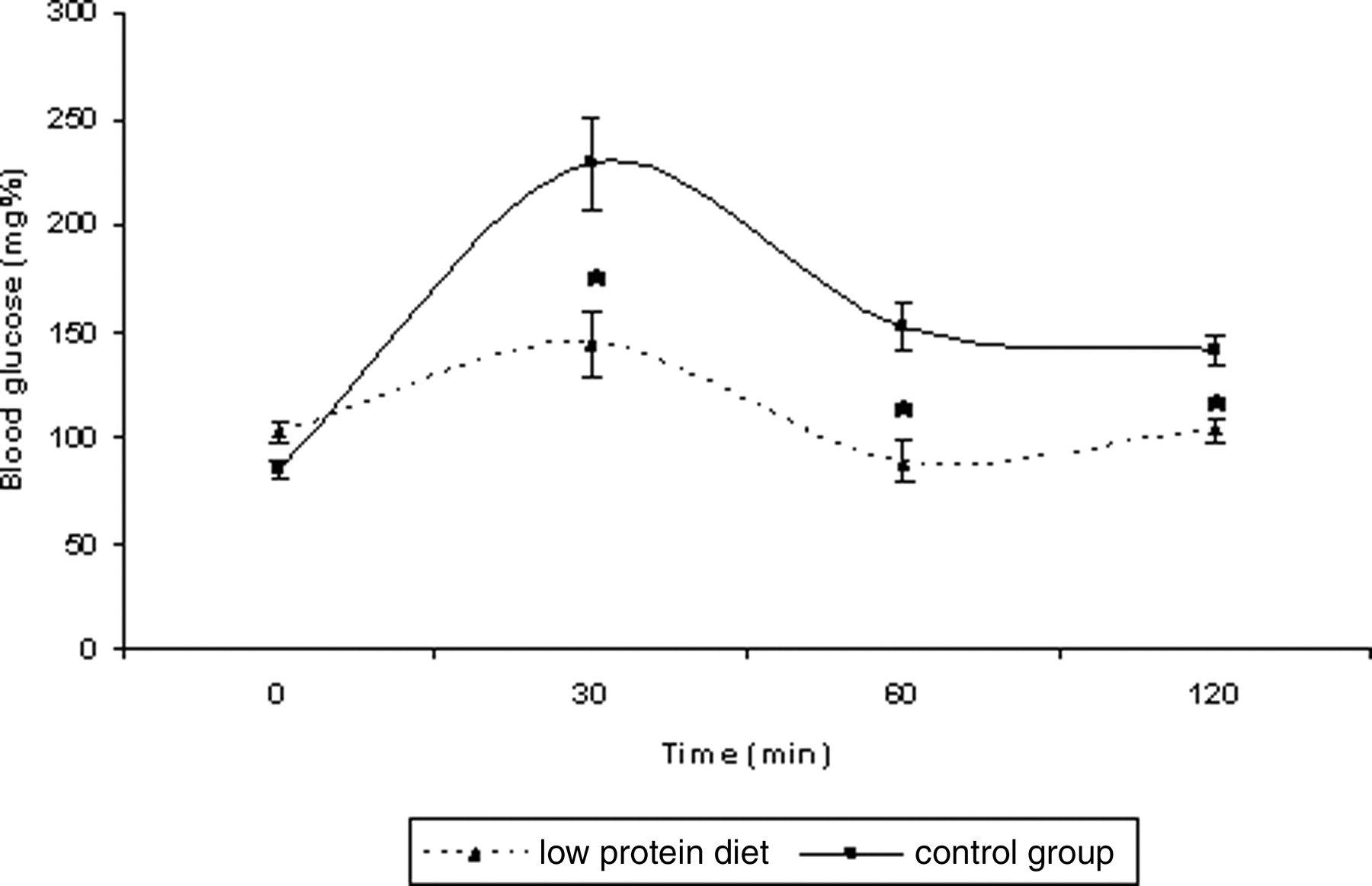
Effects of low protein diet on an intraperitoneal glucose tolerance test involving the administration i.p. of glucose (2.0 g/kg rats). The results are expressed as the mean ± SEM. The data were analyzed using the Student’s t test to compare the diets (n = 7–10). * (P < 0.05).
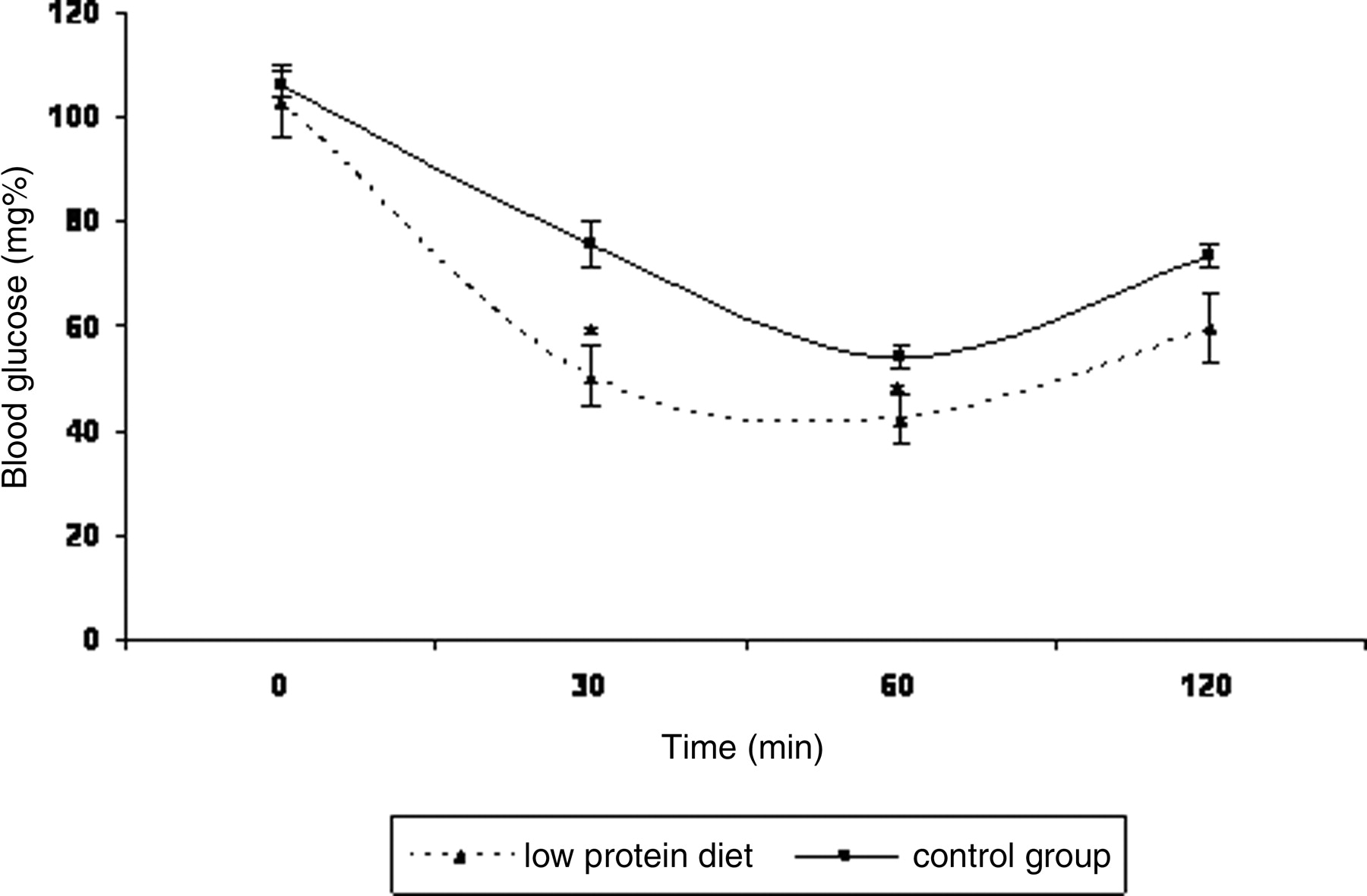
Effects of low protein diet on an Insulin Responsiveness Test involving the injection i.p. of insulin (0.75 mU/kg rats). The results are expressed as the mean ± SEM. The data were analyzed using the Student’s t test to compare the diets (n = 7–10). * (P < 0.05).
Footnotes
This research was supported by CNPq, PROSPEQ/UFRFGS, FAPERGS and by a FINEP research grant to the “Rede Instituto Brasileiro de Neurociência (IBN-Net)” #01.06.0842.
Acknowledgements
We are grateful to Blanver-Farmoquimica for providing the fiber used in the animals’ diet.