
Abstract
Background:
High-density cultures require operating below the critical threshold of shear stress, in order to avoid reducing the specific growth rate of the cells. When determining this threshold, direct inspection of the cells in flow provides insight into the conditions of shearing.
Objective:
Aim of this study was using a novel rheo-optical setup for the observation of cells in laminar shear flow and the determination of the critical shear stress required to damage them in their natural environment.
Methods:
Dunaliella salina cells were sheared and observed in flow for shear stresses of up to 90 Pa, at ambient temperature, without adding thickeners. The critical shear stress was determined by fitting a hydrodynamics-based criterion to the experimental data on the percentage of deformed cells after shearing.
Results:
Single cells, clusters and strings of cells were visible in shear flow. The strings formed at maximum shear stresses of 10 Pa or higher. Cells lost motility for maximum shear stresses higher than 15 Pa, and more than 80% of the cells were deformed at maximum shear stresses higher than 60 Pa. The estimated critical shear stress was 18 Pa.
Conclusions:
Shear stresses higher than 18 Pa should be avoided when cultivating D. salina.
Introduction
Mixing, on one hand, ensures light and substrate availability in microalgae cultivation systems, on the other hand, it introduces shear stress to the cell suspension, which might be detrimental to cell growth, or even lead to cell death [1]. To achieve optimal cell growth, shear stress needs to be kept below some critical threshold. As the continuous advances in biotechnology improve cell densities and productivities of bioreactors, the need for improving the mixing conditions is emphasized [2], thus underlining the need to operate in conditions near critical shear stress. Therefore, accurate determination of critical shear stress for a cell species is becoming increasingly important for the design and operation of biotechnological processes.
Mixing in a bioreactor is achieved by agitation, pumping or gas sparging, which introduce shear stress into the cell suspension. This is considered a key issue for the cultivation process of many microalgae [3], and has led to several investigations on the effect of shear stress on microalgae cells. Low agitation rates induced oxidative stress to P. reticulatum cells [4], and high agitation rates generated bubbles that ruptured at the surface of the culture, damaging nearby cells, where the damage threshold for stirring speed was different for P. tricornutum and P. cruentum cells [5]. Pumping led to loss of motility for T. suecica [6], chain breaking for S. costatum and damaging of H. ostrearia cells [7], depending mainly on the pump type and frequency of loops. Gas sparging increased the death rate of microalgae D. salina [8], P. tricornutum [9] and P. reticulatum [10], primarily through an increase in superficial gas velocity. Some researchers did not just investigate the effects of shear stress but also attempted to determine a critical, strain-dependent threshold. Contreras et al. [11] found that for shear rates higher than 7 · 103 s−1, there would be a decrease in the maximum specific growth rate of P. tricornutum. Camacho et al. [10] have shown that a critical shear stress as low as 0.16 mPa led to growth inhibition of P. reticulatum, and a further increase of shear stress led to cell death. Michels et al. [12] determined a critical shear stress of about 1.3 Pa for C. muelleri. However, the cells in these experiments lost viability without any visible signs of deformation. The existing literature makes it apparent that the determination of the critical shear stress is not only useful but also remains a relevant issue in biotechnological research and its applications.
Microalgae Dunaliella salina, one of the world’s richest natural sources of β-carotene, is enclosed in a plasma membrane and has no cell wall [13]. The lack of cell wall is important to mention here, because microalgae without cell wall are considered to be more susceptible to shear stress [14]. D. salina is mostly grown in large open-ponds, in areas such as Australia and Israel [15], where shear stress is avoided, or raceway ponds where shear stress is minimal [16]. As the effort on optimizing growth and carotenoid production of D. salina is still ongoing [17,18], determining the critical shear stress would allow ensuring good mixing rates, while avoiding any lethal effects on the cells. In addition, investigation of the shear stress required to damage most cells of a culture, would be useful for the design of downstream processes based on mechanical cell disruption by liquid shear stress, such as high speed or high pressure homogenization [19].
Experimental determination of the effect of shear stress on a population of cells requires a well-defined flow environment, such as a rheometer [20–22], or a capillary tube [21,23,24]. When the intensities of shear stress obtainable by the rheological equipment are not sufficient to damage the cells, thickening agents are used to increase the dynamic viscosity of the medium and thus the resulting shear stress [12,22,25]. In such cases, it must be shown that the thickener has no effect on the cells. This is performed e.g. by shearing the cells with the thickener at shear stresses below critical, and showing that no loss of cell viability occurred [12]. Nevertheless, it is obviously preferable to avoid introducing any additional effects to the system. Here, a narrow-gap rheometer with parallel plate geometry, was used [26], capable of reaching a maximum shear stress of 100 Pa without any additives. This was adequate for the investigation of cell damage for D. salina.
The goal of this work was the application of a novel rheo-optical setup for the observation of cells in laminar shear flow and the determination of the critical shear stress that leads to their deformation, without the addition of thickeners in the culture medium. D. salina cells were sheared in a narrow-gap rheometer, where the selected transparent parallel-plate geometry allowed for high-speed camera observations of cells under flow conditions. A critical shear stress criterion, based on the hydrodynamics of the parallel-plate geometry, was fitted to the experimental data in order to determine the critical shear stress for D. salina. Finally, the intensities of shear stress used for the experiments were put into perspective regarding other microalgae and industrial processes.
Materials and methods
Experimental procedure
Cell count, motility and deformation were evaluated through off-line microscope images, prior to shearing in the narrow-gap rheometer. Next, a sample of the cell culture was placed in the rheometer and shearing was initiated. During shearing, a high-speed camera video of the cells in shear flow was captured, with the camera positioned near the edge of the plates, where the cells experience maximum shear stress. Subsequently, the upper plate was raised, causing a droplet of the entire cell suspension to form in the center of the plate, making it impossible to take samples from specific radial positions. Thus, a homogenized sample was taken and used to obtain off-line microscope images of the cells after shearing.
Micro-organism and culture medium
Dunaliella salina, strain no. 184.80, was provided by the Culture Collection of Algae at Goettingen University (SAG). It was cultivated in a modified Johnson’s medium [27], with the following composition (all quantities in g/l): Na 2 EDTA, 189 · 10−6; FeCl 3 · 6H 2 O, 244 · 10−6; H 3 BO 3, 61 · 10−6; (NH 4)6 Mo 7 O 24 · 4H 2 O, 38 · 10−6; CuSO 4 · 5H 2 O, 6 · 10−6; CoCl 2 · 6H 2 O, 5.1 · 10−6; ZnCl 2, 4.1 · 10−6; MnCl 2 · 4H 2 O, 4.1 · 10−6; MgCl 2 · 6H 2 O, 1.5; MgSO 4 · 7H 2 O, 0.5; KCl, 0.2; CaCl 2 · 2H 2 O, 0.2; KNO 3, 1; NaCl, 65; NaHCO 3, 0.04; KH 2 PO 4, 0.04. Cells were grown in 1 l flasks placed 30 cm away from a warm light source (Osram T8 L 18W/827 LUMILUX INTERNA G13, color temperature 2700°K), with an alternating day/night cycle (12 h, 12 h). Subcultures were created every 2 weeks, placing 500 ml of the culture and 500 ml fresh medium in a 1 l flask.
Cell count and deformation
D. salina are highly motile biflagellate cells that use their flagellas for movement. The cells move constantly and relatively fast when observed in the microscope under normal conditions, so that their motility cannot be mixed up with random fluid movement, and can therefore be considered as a measure of cell viability. However, this constant cell movement results in the cells continuously shifting in and out of focus, an issue that could affect the accuracy of the cell count. The answer to this issue is the addition of 5% Lugol Potassium-Iodine solution to samples containing moving cells before taking the off-line microscope images. This results in immediate loss of cell motility and sedimentation, which allows obtaining well-focused images and therefore accurate cell counts. For the cell counts, a sample from the cell suspension was placed in a Thoma cell-counting chamber with a depth of 0.1 mm. The cell concentration of each measurement was in the order of 106 cells/ml. The volumetric proportion of the cells in the suspension was up to about 1%, which suggests that no significant cell–cell interaction should be expected during the measurements.
Regarding cell deformation, cells were differentiated between intact, i.e. the cells that retained the original ellipsoidal shape of D. salina, and deformed, i.e. the cells that visibly deviated from this shape. The distinction is presented in Fig. 1. The percentage of intact cells was calculated as follows:
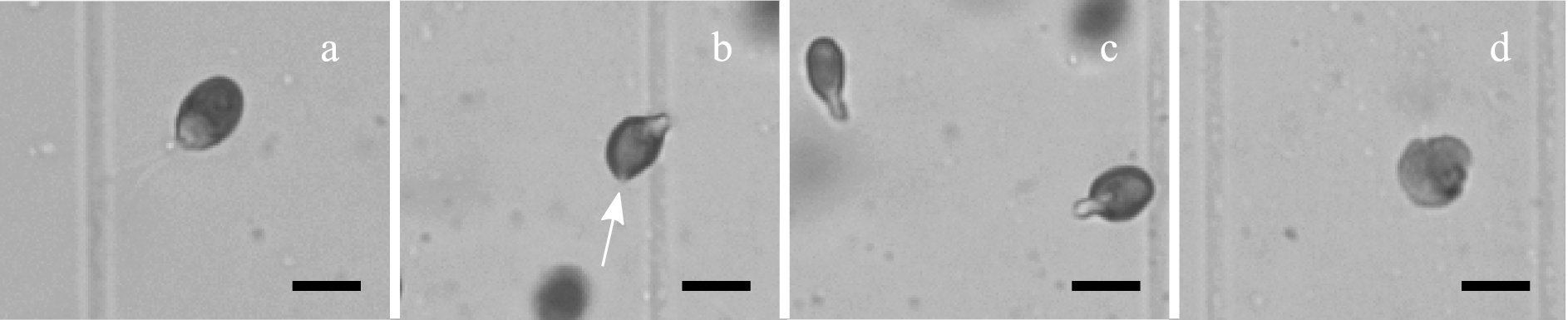
Four types of cells visible in the off-line microscope images after shearing. (a) intact; (b) deformed with a visible rupture of the cell membrane; (c) deformed and elongated where the flagellas are usually attached; (d) deformed with a collapsed membrane structure. Scales represent 10 µm.
The experiments were performed with a narrow-gap rheometer working in the parallel-plate configuration, where the upper plate has a 50 mm diameter and the lower plate 75 mm. The narrow-gap rheometer is based on a rotational rheometer (UDS 200 from Physica) that was modified to gain a gap-width precision of ±0.7 µm. To obtain this precision, we used optical windows from Melles Griot as plates and aligned them parallel to each other and perpendicular to the rotational axis employing a confocal interferometric sensor, which allows determination of the gap width independently from the rheometer reading. The rheometer was recalibrated daily, following a procedure explained in detail in a previous publication [26].
The cells were sheared after two weeks of growth, at which point they were in the stationary phase. 200 µl samples were placed between the parallel plates and sheared in constant maximum shear stresses between 0 and 90 Pa for a duration of 1000 s. A gap of 30 µm was selected, which allowed free cell flow, since D. salina cells have an average major axis length of 10 µm. After each experiment the upper plate was raised and a homogenized sample was taken for the microscope measurements. The experiments were performed at ambient temperature, which fluctuated between 24–26°C. Viscous heating was considered negligible for these experiments, as previously shown [26].
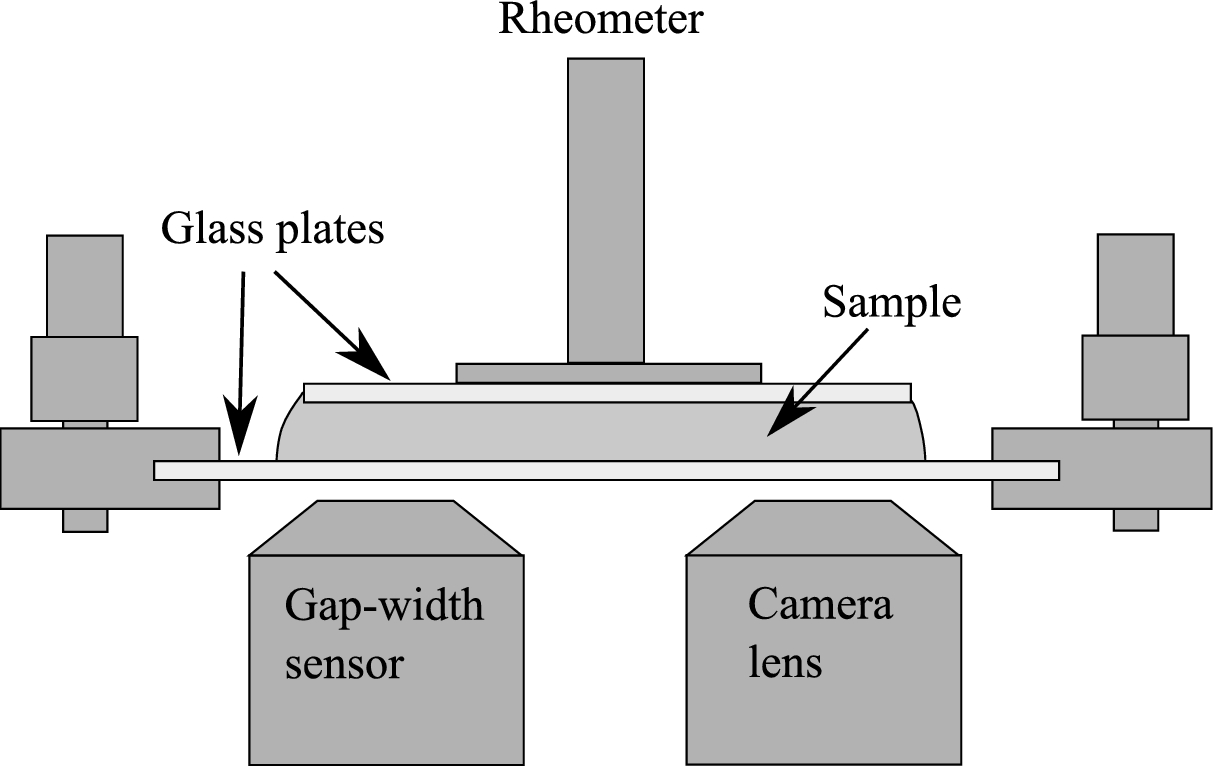
Sketch of the rheo-optical setup. The top plate rotates and is connected to the rheometer. The bottom plate is stationary, and the position of the camera lens can be adjusted along the radius of the bottom plate. For details on the use of the gap-width sensor we refer to [26].
The transparent glass plates of the parallel-plate geometry allowed for witnessing cells in shear flow with the use of a high-speed camera (Mikrotron Phantom Miro M310) connected to a 20×-lens, as shown in Fig. 2. The camera was placed under the bottom plate of the geometry and the lens was pointed to the outer rim of the plate, where the cells experience the maximum shear stress for a given shear rate. Additionally, a white LED-lamp (VLP IntraLED 2020/W (Volpi)) connected to a light guide (Lumatec 3716) was used to shed light directly from above at an angle of 45° with respect to the camera axis. The lamp was placed about 3 cm away from that area, and was turned off immediately after each experiment, to avoid continuous heating of the area and heat-shock-induced cell death. A sample frame rate of 10,000 fps with an exposure time of 10 µs were adequate to observe cells in flow for a maximum shear stress up to 70 Pa. For the cases of 80 and 90 Pa, frame rates of 16,000 fps and 18,000 fps, and exposure times of 10 µs and 20 µs were used, respectively. The exposure time was increased in the case of 90 Pa to provide adequate lighting conditions for the video.
Rheology
The narrow-gap rheometer measurements were performed, using a parallel-plate geometry, for which the shear rate
The distribution of shear stresses is not homogeneous in the gap between the two plates, since it depends on the position r on the plate. The radial distribution is characterized by:
Shear stress is not easily obtainable for complex flows, as they arise e.g. in a stirred tank or a pump, and thus another parameter is necessary to compare between different processes. Here we use the energy dissipation rate ε [2]:
When the energy dissipation rate is multiplied by the duration of the process, it gives the total energy dissipation E. This is another measure considered relevant for process design and comparison [25], as different processes could reach the same levels of total energy dissipation either by adjusting the energy dissipation rate or the shearing duration. Therefore, both the energy dissipation rate and the total energy dissipation are relevant when designing a cultivation system or a downstream process for D. salina cells.
Critical shear stress criterion
To determine the safe operational levels of shear stress, we assume there is a critical value of shear stress
Fitting Eq. (6) to the experimental data for each maximum shear stress where cell deformation occurred, i.e. in cases where at least 20% of the cells were deformed, determines the critical shear stress. The least-square fitting of the equation was performed using the curve-fitting toolbox in MATLAB (version R2015a), and provided the best fit along with 95% confidence intervals for the coefficient.
Results and discussion
Rheological characterization
The measured maximum shear stress increased linearly with an increase in maximum shear rate, while the apparent viscosity remained relatively constant for maximum shear rates in the range of 2 · 103–105 s−1 (less than 5% difference from average viscosity), which indicates the cell suspension acted as a Newtonian fluid. The average dynamic viscosity measured in that range was 1.14 ± 0.01 mPa · s (with 95% confidence). At lower shear rates than the chosen range, the torque resolution of the narrow gap rheometer was too low to accurately measure the viscosity, and at higher shear rates inertia forces were too strong, pushing the suspension out of the gap between the parallel plates.
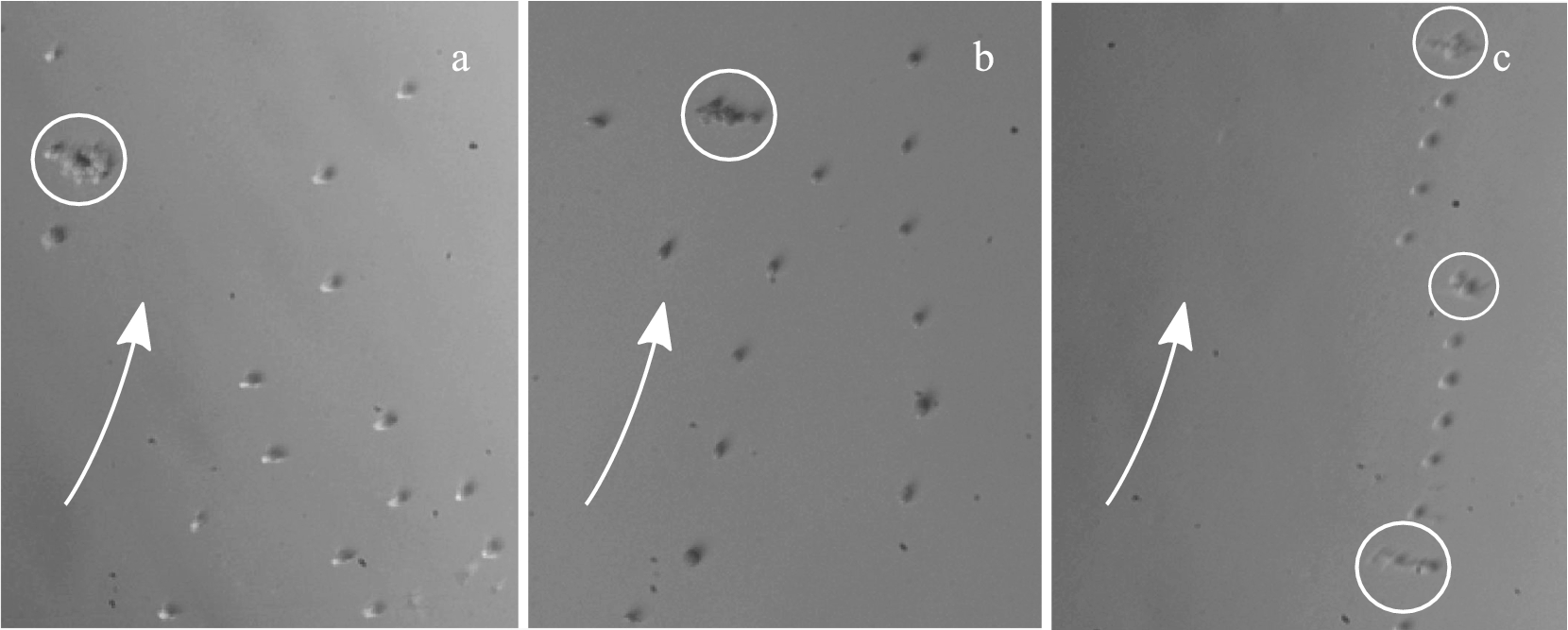
Formations of cells in laminar flow depending on maximum shear stress: (a) 5 Pa; (b) 15 Pa; (c) 70 Pa. Arrows indicate direction of flow, circles indicate clusters of cells. The in situ high-speed camera frames show cells experiencing maximum shear stress, since the frame location was positioned near the edge of the plates.
At the beginning of each experiment, before shearing was initiated, the cells moved unobstructed in the suspension within the gap, without any signs of immobilization or attachment to the plates. Single cells could be observed in flow and for a maximum shear stress of 10 Pa, cells started forming strings, while for maximum shear stresses higher than 15 Pa, most cells arranged as strings, as shown in the in situ high-speed camera frames presented in Fig. 3. This phenomenon has already been observed for suspensions of polystyrene spheres close to the wall of a parallel-plate setup [28]. Also, clusters of cells appeared for all shear stresses investigated, forward-tumbling in the flow direction without visibly breaking up into smaller clusters. The clusters consisted of about 5 to 10 cells and were generally stretched in the vortex direction.
Effect of shearing intensity on D. salina
As presented in Fig. 4, increasing the maximum shear stress resulted in a decrease in the percentage of visible intact cells. Up to a maximum shear stress of 15 Pa, more than 90% of cells remained intact and motile. When the cells were immobilized with Lugol Potassium-Iodine solution, the cell flagellas could be observed in the off-line microscope images. For maximum shear stresses higher than 15 Pa, all cells lost motility and no flagellas were visible in the images. For a maximum shear stress higher than 60 Pa more than 80% of the cells were deformed after shearing, some with visible points of rupture in their membrane, as shown in Fig. 1(b). It is clear that the loss of cell motility is observed only where a significant reduction in the percentage of intact cells occurs (at least 20% reduction), i.e. for maximum shear stresses higher than 15 Pa. This suggests that the critical shear stress should be in the same order of magnitude.
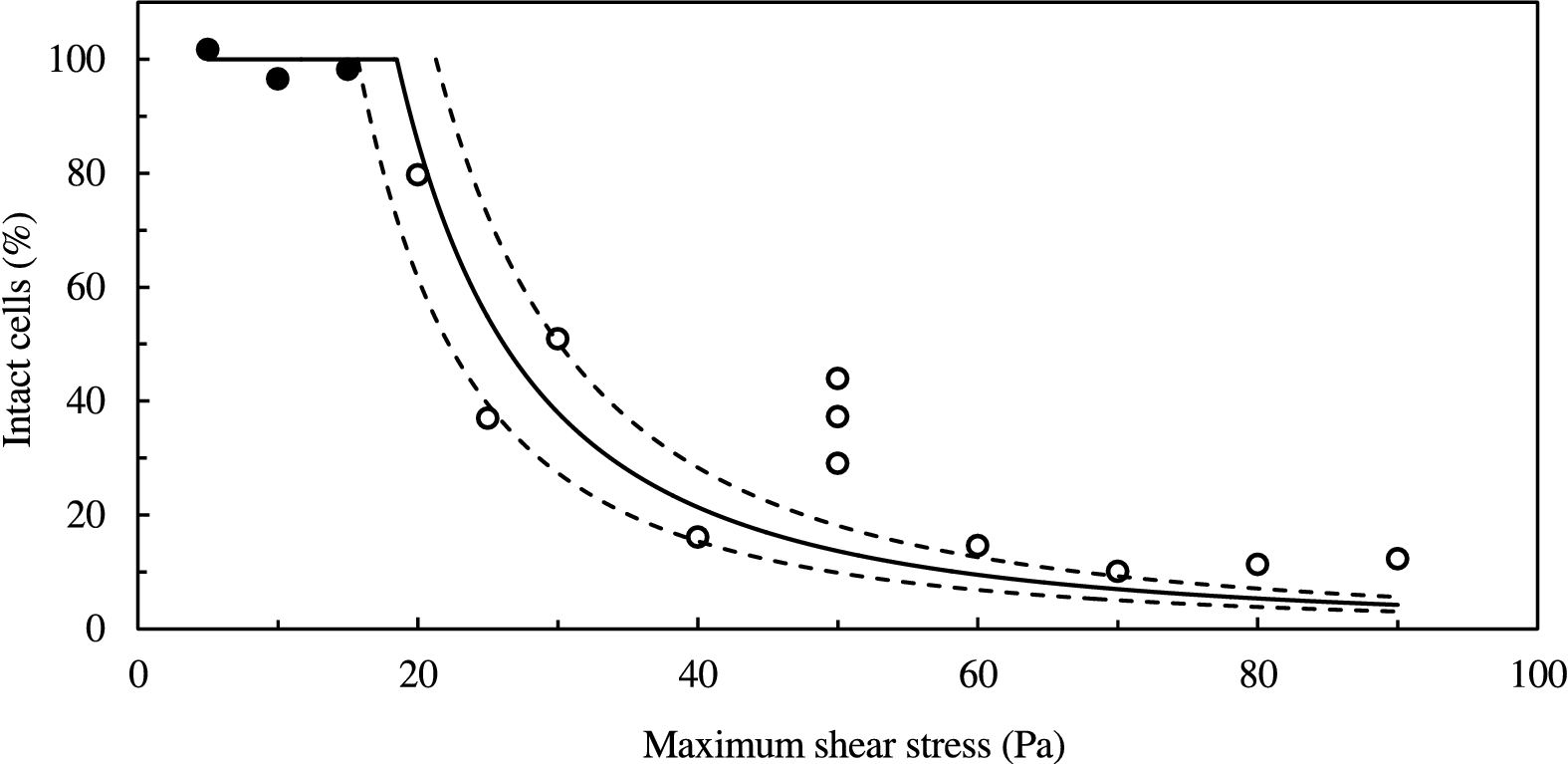
Effect of shear stress on the percentage of intact cells: (filled circles) experimental data of intact and motile cells; (empty circles) experimental data of intact but non-motile cells; (solid line) best least-square fit of Eq. (6); (dashed lines) 95% confidence interval. Maximum shear stress denotes the shear stress at the edge of the plate.
Both cell motility loss and cell deformation are considered indicators of cell damage. However, we consider the first as a weaker indicator, as it was shown for other flagellate microalgae that motility can be regained. More specifically, in the shearing experiments of Jaouen et al. [6] on the flagellate microalgae T. suecica, it was determined that 40% of the population of intact cells regained motility after a 5 h resting time. In our case, a significant impact on the cells is shown by both indicators for a maximum shear stresses higher than 15 Pa, additionally supporting the notion that the critical shear stress would be in the same order of magnitude.
The critical shear stress determined was 18 ± 3 Pa (with 95% confidence) for laminar flow, based on the critical shear stress criterion, as described in Eq. (6) and as shown in Fig. 4. As the critical shear stress criterion refers to cases where at least 20% deformed cells were observed after shearing, the non-linear regression curve was fitted to the data for a maximum shear stresses higher than 15 Pa. The fitting included some outliers, such as the measurements for a maximum shear stress of 50 Pa. However, we were unable to connect these outliers to some systematic error. The critical shear stress corresponds to a critical shear rate of 1.6 · 104 s−1, and a Reynolds number of 12. The estimated critical shear rate is twice as high as the one required to decrease the maximum specific growth rate of P. tricornutum [11]. Additionally, our estimation of the critical shear stress for D. salina is five orders of magnitude higher than the one for P. reticulatum [10], and one order of magnitude higher than the critical shear stress of C. muelleri [12]. Forming conclusions on the shear sensitivity of different strains would be ill-advised in cases where cells were subject to different flow conditions. This holds for both the cases of P. tricornutum and P. reticulatum, where cells were sheared in turbulent flow [10,11], as well as for C. muelleri, where death by shear stress was only observed in the samples where locust bean gum (LBG) was used as a thickener [12].
Time dependence of the shear deformation was not necessary for the critical shear stress criterion to adequately fit the experimental data. This indicates that 1000 s of shearing were suitable for the deformation of most D. salina cells. This is in line with the experiments of Michels et al. [12], where it was shown that constant high shear stress required only a few minutes to negatively affect the microalgae C. muelleri. The duration of shearing is negligible when compared to the usual cultivation time of D. salina, which lasts several days [17]. Therefore, reaching a damaging level of shear stress in the bioreactor would immediately impact the specific growth rate of the culture, which underlines the significance of determining the critical shear stress.
The energy dissipation rate corresponding to the critical shear stress is 1.7 · 105 W/m3, and the total energy dissipation for the critical shear stress is 1.7 · 108 J/m3. To put this energy dissipation rate into perspective, it is in the same order of magnitude as the maximum local energy dissipation rate of an impeller in a 10 l reactor operating at 700 rpm [29], the maximum local energy dissipation rate of an impeller in a 22 l reactor operating at 140 rpm [30], or a bubble with a diameter of 6.32 mm rupturing [31]. Dunlop et al. [25] used the total energy dissipation per unit volume as a measure for the effect of shear stress on carrot cells. They found that turbulent shear flow could lead to cell lysis for total energy dissipation up to four orders of magnitude lower than was required for cell lysis in laminar shear flow. This is a possible additional effect that should be taken into account when designing the cultivation process of D. salina. Damaging the cells occurred at maximum shear stresses between 20 and 90 Pa and at energy dissipation rates between 105 and 107 W/m3, knowledge applicable in the design of downstream processing for these cells, specifically downstream processing that is based on cell rupture by liquid shearing.
Conclusions
D. salina cells were sheared in a narrow-gap rheometer and observed in flow using a rheo-optical setup. No thickeners were required to achieve high shear stresses, which allowed shearing the cells in their natural environment. Cells assembled into strings for maximum shear stresses of 10 Pa or higher, and some cell clusters were visible in all cases. A laminar, maximum shear stress of 15 Pa was the highest maximum shear stress at which the D. salina cells remained fully intact and motile. For higher stresses, cells lost motility, and an increase in maximum shear stress led to an increase in the percentage of deformed cells. For a maximum shear stress of 60 Pa and higher, more than 80% of the cells were deformed.
Fitting a critical shear stress criterion to the experimental data, allowed for the determination of a critical shear stress of about 18 Pa, as the lower boundary of disruptive laminar shear stresses for D. salina. Though not including any time dependency for cell disruption, the shear stress criterion was well capable to represent the trend of the experimental data. The duration of shearing was 1000 s in all experiments and is negligible when compared to the usual cultivation time of D. salina. Thus, we can expect an immediate effect of shear stress on the specific growth rate of the cell culture, which increases the importance on designing cultivation systems that work below the critical threshold of shear stress.
The energy dissipation rate corresponding to the estimated maximum shear stress was 1.7 · 105 W/m3, and the critical total energy dissipation was 1.7 · 108 J/m3. Higher energy dissipation rates can be detrimental to the cells and result in lower specific growth rates, therefore they should be avoided when cultivating D. salina. Nonetheless, cell-damaging energy dissipation rates between 105 and 107 W/m3 might be relevant to the design of downstream processing for these cells.
Footnotes
Acknowledgements
The Bayerische Forschungsstiftung is gratefully acknowledged for the financial support of this work, carried out through grant DOK-147-12. Also, H.D. gratefully acknowledges financial support by the Deutscher Akademischer Austauschdienst (DAAD), Germany and the Ministry of Higher Education & Scientific Research, Iraq. Finally, we thank the Culture Collection of Algae at Goettingen University for providing the necessary microalgae strain.