
Abstract
Background:
Although many studies have shown that arteries change diameter in response to chronic change in blood flow (BF), keeping wall shear stress (WSS) at physiologically normal levels, relatively little is known about the effects of flow restoration after flow reduction and also the role of vascular smooth muscle (VSM) during such a remodeling process.
Objective:
To elucidate the biomechanical responses of the arterial wall to the restoration of normal BF after flow reduction and compare the results with our previous results observed in response to decreased BF alone.
Methods:
Carotid artery BF in the Wistar rat was decreased by ligation and then restored to normal levels by release of the ligation. The effects of BF changes on the biomechanical properties of the carotid arterial wall were determined from measurements of diameters and pressures of excised artery segments.
Results:
During BF reduction and restoration, WSS was maintained at physiological levels by changes in the internal diameter. No significant changes in the incremental elastic modulus were found in response to changes in BF. VSM tone was significantly enhanced during the changes in BF.
Conclusions:
Arteries change diameters in response to BF reduction and also flow restoration to normal after flow reduction, keeping WSS at physiologically normal levels. The lack of changes in vascular elasticity suggests that there were no significant changes in major wall constituents, such as elastin and collagen. VSM may play the dominant role in observed arterial remodeling and adaptation.
Keywords
Introduction
Living organs, tissues, and cells functionally adapt to mechanical demands and efficiently change their geometry, structure, and properties [1]. This phenomenon is called “Tissue Remodeling” [2]. More than a hundred years ago, Thoma [3] hypothesized that the diameter of blood vessels is regulated by the magnitude of blood flow (BF), while the thickness of vessel wall changes depending on the magnitude of tension generated by blood pressure (BP) [4]. Thereafter, many studies have been performed on the arterial response to chronic changes in BF and BP [5], because the phenomenon is interesting and important not only in basic biomedical science but also in clinical medicine. The structure and properties of arterial wall are strongly affected by such hemodynamic forces as BF and BP. Since vascular pathologies are related to alterations in these hemodynamic loads, the sensitivity of arterial structure to these changes inevitably affects the initiation and progression of vascular diseases [6].
Regarding the effects of BF on arterial diameter, Kamiya and Togawa [7] proposed constant wall shear stress regulation for the first time on the basis of their experimental results obtained from an arterio-venous shunt model between the common carotid artery (CCA) and the external jugular vein in the canine. They observed that the carotid internal diameter increased with chronic increase in BF and vice versa, keeping wall shear rate, which is equivalent to wall shear stress, at a constant, physiologically normal level, and suggested that arterial diameters are controlled by a physiologic negative feedback system. Thereafter, Langille and his group performed a series of several important studies on BF-diameter relationships and roles of cells in the compensatory arterial response to chronic changes in luminal BF [ex. [8–12]]. Recently, we have performed a systematic experimental study on BF-diameter and BP-wall thickness relationships using normotensive and hypertensive animals, and confirmed that there are flow- and pressure-mediated control functions in the vascular system [13].
Arterial BF and BP induce two major hemodynamic stresses: (1) wall shear stress (WSS), and (2) wall tensile stress (wall hoop stress, WHS), respectively. Blood vessel wall is optimally designed to correspond well to these stresses at physiological state, and also changes its structure and properties so as to adapt to changes in stresses as mentioned above. For example, chronic changes in BF lead to changes in arterial diameter, while increased BP induces arterial wall thickening. We have considerable evidence indicating that such arterial remodeling occurs in order to restore WSS and WHS to their physiologically normal values, that is to preferred homeostatic levels [5,14]. The flow- and pressure-mediated adaptation and remodeling have become of major interest lately, because the phenomena are very interesting and important not only in basic biomedical science but also for clinical medicine [6,15]; in particular, the sensitivity and response of vascular structure to BF changes inevitably affect the development and progression of vascular diseases as stated above [16].
A number of studies have been performed on the arterial response to chronic changes in BF, which indicate that arterial inner diameters change in response to the alterations of BF so that WSS is maintained at constant. Many of these experiments have been done with increased BF. However, relatively little is known about the effects of the reduction of BF and also the restoration of normal BF after decrease. Understanding of the processes involved in arterial remodelling is prerequisite not only in clinical medicine but also for basic vascular physiology and mechanics. In addition, there is a paucity of information on the activities of smooth muscle cells (SMCs) during the remodelling process, although it has been estimated that their role is one of the key factors in the phenomenon. Based on this background, the present study was performed to experimentally determine the biomechanical response of the arterial wall to the restoration of normal BF after flow reduction. The results obtained were compared with those observed in the experiments of a decrease of BF alone.
Materials and methods
Materials and hemodynamic measurements
All animal experiments and procedures were carried out within the Animal Welfare Regulations and Guidelines for Animal Experiments, Graduate School of Engineering Science, Osaka University, by the approval of the Committee for Animal Experimentation in the School.
We used 42 male Wistar rats aged 8 weeks; 24 rats were used for the experiments on the effects of BF changes, while the remaining 18 animals were used to obtain age-matched, control data. First, the left CCA in each of the 24 experimental animals was surgically exposed under anesthesia with sevoflurane (1 ml/min of Sevofrane™). Then, for the measurement of BF in the CCA, a flow probe (1 mm in diameter and 7 mm in length) for an ultrasonic transit time system (T106-108, Transonic Systems, Ithaca, New York) was mounted on the artery between 5 and 13 mm proximal from the bifurcation to the external carotid artery (ECA). BP was monitored in the abdominal aorta using an implantable telemetric transducer system (PA-C40, Data Sciences International, St. Paul, Minnesota). A fine indwelling catheter (0.7 mm in diameter) having a pressure transducer at the tip and a signal transmitter (7.8 g in weight and 4.5 ml in volume) at the other end was inserted into the abdominal aorta from approximately 20 mm distal to the entrance to the left renal artery. The transducer was positioned in the aorta 40 mm proximal from the entrance to the right renal artery, while the transmitter was placed inside the peritoneal cavity. Then, to reduce BF in the left CCA, the artery was constricted between the flow probe and the bifurcation to the ECA. To make the same degree of constriction in all the experimental animals, an injection needle (0.55 mm in diameter) was placed in parallel with the CCA, and the artery and the needle were tied with braided silk suture, and then the needle was carefully removed. BF in the left CCA and BP were measured simultaneously before and soon after the constriction of the artery, recorded on a digital recorder (Omniace RT3200N, NEC San-ei, Tokyo, Japan), and fed to personal computers. After these procedures, the flow probe and the pressure transducer were removed, and the skin in the neck and abdomen was closed. All the animals were kept in cages during each prescribed period, being given a standard rat chow and tap water ad libitum.
At 2 weeks after the constriction of the CCA, the artery in each of the 24 animals was exposed again under anesthesia, and the flow probe and pressure transducer were attached to the CCA and the abdominal aorta, respectively, again by the procedures mentioned above. After BF and BP were measured as the data immediately before the release of constriction, left CCAs were excised from 6 animals out of 24 for biomechanical tests by the procedures described in the next section. In each of the remaining 18 rats, the braided silk suture used for carotid constriction was cut and removed for the restoration of BF, and then BF and BP were simultaneously measured again by the procedures mentioned above. From 6 animals out of 18, left CCAs were excised for biomechanical tests to obtain data at 0 week. The remaining 12 rats were kept alive after closing the skin. At 2 and 4 weeks after the release of constriction (6 animals for each period), they were euthanized after the measurements of BF and BP in vivo, and their left CCAs were excised for biomechanical tests.
Biomechanical testing
Following euthanasia by a lethal dose of pentobarbital sodium, the left CCA was carefully exposed and marked with gentian violet on the surface at several points 2.5 mm apart in the axial direction. Then, the CCA of approximately 15 mm in length was excised from the proximal part, and rinsed with a Krebs–Ringer solution. After the distances between the marks were measured with a scale to determine the in vivo axial extension ratio, each segment was stored in a fresh Krebs–Ringer solution kept at 4°C. An approximately 10 mm long specimen was obtained from the proximal site of the segment for biomechanical testing (pressure-diameter test, P-D test). The remaining distal segment with a length of approximately 5 mm was used for the measurement of arterial dimensions under no load conditions and also for future histologic studies.
For P-D tests, we used the apparatus and the procedures similar to those reported previously [17,18]. Briefly, each tubular segment was attached to a specimen holder, stretched to the in vivo length in reference to the marks previously put on the surface, and was set up in a test bath filled with a Krebs–Ringer solution of 37°C bubbled with 95% O2 and 5% CO2 gas mixture. Its external diameter, Do, and internal pressure, Pi, were measured with a video dimension analyzer (C3160, Hamamatsu Photonics, Hamamatsu, Japan) and a fluid-filled pressure transducer (MPU-0.5-290-0-3, Orientec, Tokyo, Japan), respectively. First, the artery was preconditioned by several times of inflation-deflation procedures between 0 and 200 mmHg at the rate of 3.3 mmHg/sec until we obtained reproducible Pi-Do curves. Then, the inflation-deflation was repeated once more at the rate of 1.7 mmHg/sec, and the data obtained during the inflation process were used as the data under “normal condition” with physiological vascular smooth muscle (VSM) tone (basal VSM tone). Pi was then increased to 100 mmHg. While the pressure was maintained at that level, papaverine was added to the bath to the concentration of 10−4 M. After relaxation became maximal, inflation-deflation loops were repeated between 0 and 200 mmHg at the rate of 1.7 mmHg/sec until reproducible Pi-Do curves were observed. Then, the final inflation curve was used as the data under “passive condition” (maximal VSM relaxation).
All the biomechanical tests were completed within 12 hours post-mortem for each vessel. We have so far performed many experiments on arterial mechanics, and confirmed that the basal tone and contractility of VSM and SMCs are preserved at least for 12 h after the excision of vascular specimens (for example, [13,17,18]).
Dimensional measurement
The external diameter,
Data analysis
Assuming that arterial wall is incompressible [18,19], internal diameter,
With the assumption of Poiseuille flow, mean wall shear stress (WSS), τ was calculated from [19]:
Mean wall stress in the circumferential direction (hoop stress, WHS),
Arterial contraction was evaluated from the diameter response,
To express the elastic properties of wall material, we used the following incremental elastic modulus in the circumferential direction of vessel [21]:
Statistical analysis
All data were expressed as means ± standard deviations (SDs). A one-way analysis of variance (ANOVA) was used to evaluate statistical dependences among experimental groups. If significant differences were confirmed with one-way ANOVA, then we applied Tukey–Kramer (T-K) test for post-hoc multiple comparisons between individual groups. Differences were evaluated to be significant, if the probability level (p) was below 0.05 (5%).
Results
Changes in blood flow and blood pressure
Mean BF in the left CCA was significantly decreased from 8.3 ± 1.6 ml/min (BC in Fig. 1) to 3.4 ± 1.1 ml/min (AC; 59% decrease from BC) by the constriction of the distal part of the artery. The low BF was maintained for the subsequent 2 weeks; it was 4.3 ± 1.6 ml/min after 2 weeks of constriction (BR; 48% decrease from BC). Immediately after the release of constriction at 2 weeks, BF was significantly increased and restored to 7.3 ± 2.5 ml/min (AR); there was no significant difference in BF between before constriction (BC) and soon after the release of constriction (AR).
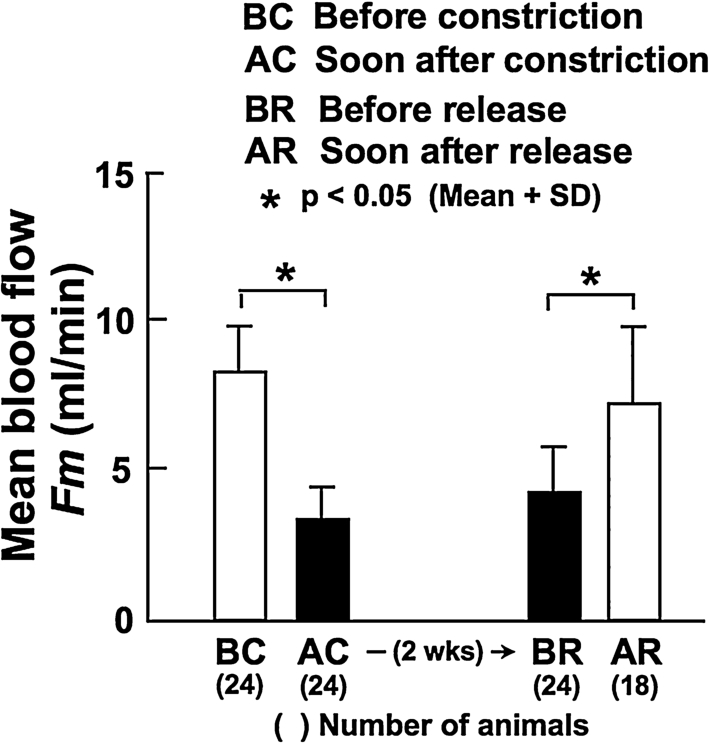
Mean blood flow in the CCA measured before (BC) and soon after (AC) the constriction of the ipsilateral, distal CCA, at 2 weeks after constriction (before the release of constriction, BR), and soon after the release of constriction (AR).
Figure 2 demonstrates mean BF measured in the animals of which left CCAs were harvested for biomechanical tests. There was a significant difference in BF in the left CCA between soon before (BR) and immediately after the release of contraction (0 wk) (3.8 ± 1.5 vs. 6.5 ± 1.2 ml/min). Also at 2 and 4 weeks after the release of constriction, BF was significantly larger compared with BF measured prior to the release (6.6 ± 1.2 and 8.1 ± 1.4 ml/min at 2 and 4 weeks, respectively). There were significant differences in BF between the experimental animals and the age-matched, non-treated ones at 0 and 2 weeks. However, the difference became insignificant at 4 weeks, which indicates that BF was recovered to the normal level at 4 weeks after the release of constriction.

Mean blood flow in the CCA measured before the release of constriction (BR), and soon after (0 wk) and at 2 and 4 weeks after the release of constriction. Data from age-matched, non-treated control animals are also demonstrated. All of these arteries were used for biomechanical tests.
In contrast, there were no significant differences in BP among all the experimental animals and controls (data not shown). For example, mean BP was 103.5 ± 7.3 and 83.4 ± 12.8 mmHg immediately before and after the release of constriction, respectively, and 101.0 ± 4.8 mmHg at 4 weeks after the release of constriction.
There was no significant increase in the internal diameter of the left CCA immediately after the release of constriction, and the diameter at that time was significantly smaller compared with the age-matched control animals (Fig. 3). Thereafter, however, the diameter progressively increased for the following 4 weeks; there were significant differences between the diameter before the release of constriction and those at 2 and 4 weeks, and the diameter difference from the age-matched control became insignificant at 4 weeks.
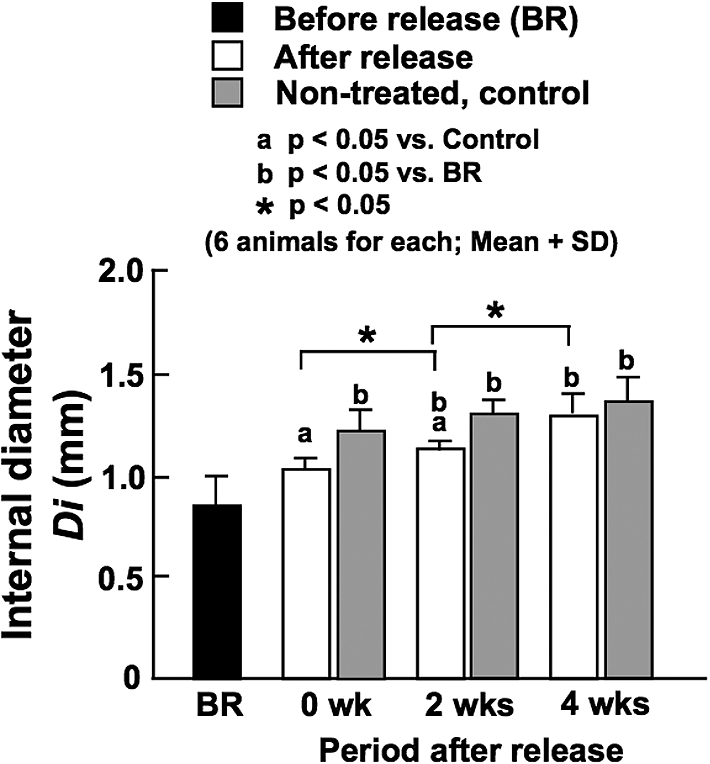
Internal diameter of the CCA determined at in vivo working blood pressure before the release of constriction (BR), and soon after (0 wk) and at 2 and 4 weeks after the release of constriction. Data from age-matched, non-treated control animals are also demonstrated.
After the release of constriction, WSS increased due to the increase of BF rate. However, the increases of WSS were not statistically significant, and the stress was not significantly different among all the experimental groups and age-matched controls; it ranged between approximately 2 and 4 Pa (Fig. 4).
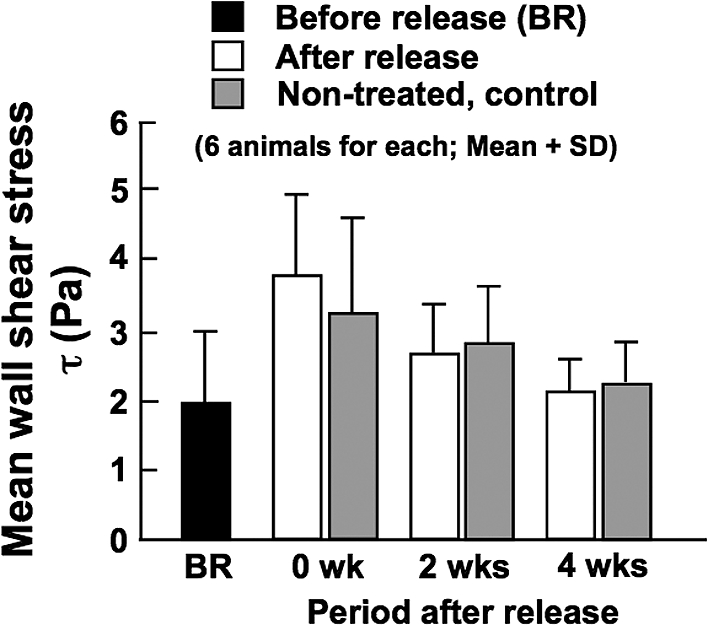
Calculated mean wall shear stress in the CCA before the release of constriction (BR), and soon after (0 wk) and at 2 and 4 weeks after the release of constriction. Stress values for age-matched, non-treated control animals are also demonstrated.
Like BP, there were no significant differences in wall thickness among all the experimental and control animals (data not shown). The ratio of wall thickness to internal radius at in vivo working BP was not changed much by the release of constriction (BR vs. 0 week in Fig. 5), but it gradually decreased with time and reached the control level at 4 weeks when the normal flow was restored. The ratio before (BR) and that soon after (0 wk) the release of constriction were larger compared with the control values. As there were no significant differences in BP among all the animals as mentioned above, the increased ratio of thickness to radius observed before and at 0 and 2 weeks after the release of constriction (Fig. 5) decreased WHS in these groups (Fig. 6). However, WHS was returned to the control level of approximately 130 kPa at 4 weeks after the release of constriction, indicating a blood pressure-mediated remodeling of arterial wall [14].
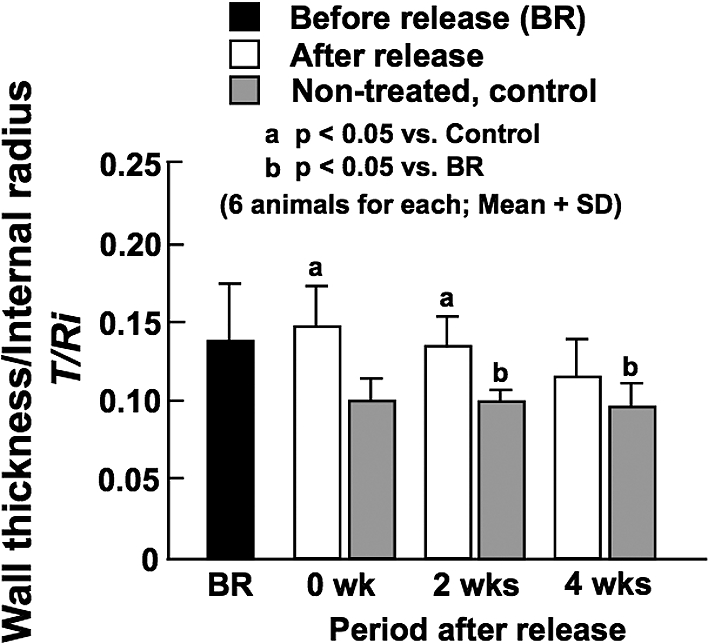
Ratio of wall thickness to internal diameter determined at in vivo working blood pressure before the release of constriction (BR), and soon after (0 wk) and at 2 and 4 weeks after the release of constriction. Data from age-matched, non-treated control animals are also demonstrated.
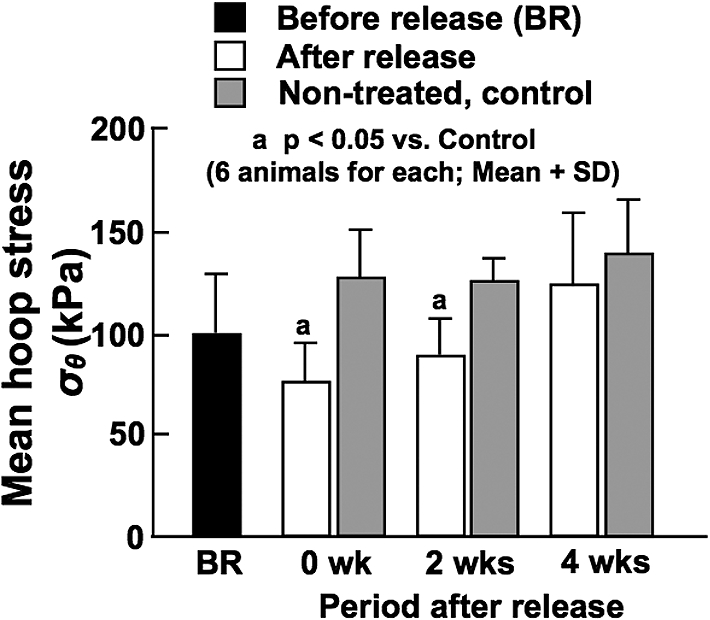
Calculated mean wall hoop stress in the CCA before the release of constriction (BR), and soon after (0 wk) and at 2 and 4 weeks after the release of constriction. Stress values for age-matched, non-treated control animals are also demonstrated.
Before the release of constriction, that is during the phase of reduced BF, diameter response at in vivo working BP under the normal condition, which represents the degree of basal VSM tone, was significantly larger compared with control arteries (Fig. 7). This continued until 2 weeks after the release of constriction. These results indicate that basal VSM tone was enhanced by BF changes, which may play an important role in the remodeling of arterial wall. The diameter response was returned to control level at 4 weeks after the release of constriction, which corresponded well to the restoration of BF. There were no significant effects of BF changes on the elastic modulus of arterial wall (Fig. 8).
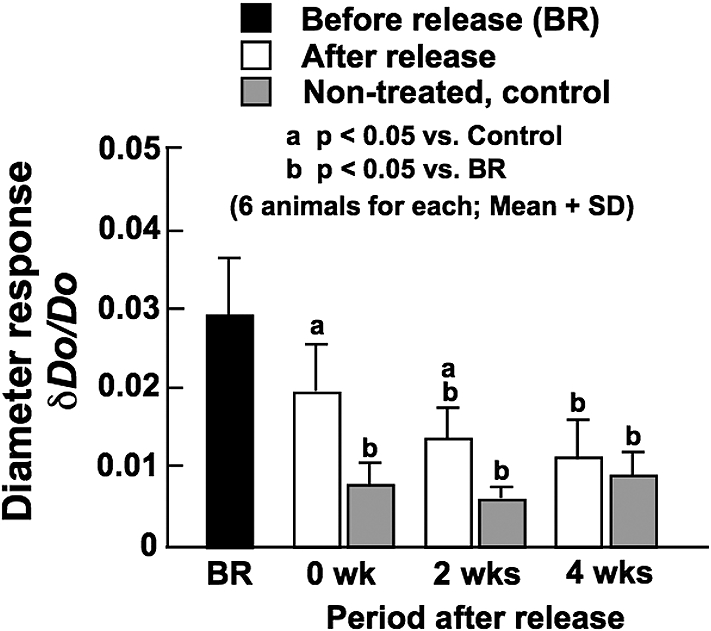
Diameter response of the CCA determined at in vivo working blood pressure under the normal vascular smooth muscle condition before the release of constriction (BR), and soon after (0 wk) and at 2 and 4 weeks after the release of constriction. Data from age-matched, non-treated control animals are also demonstrated.
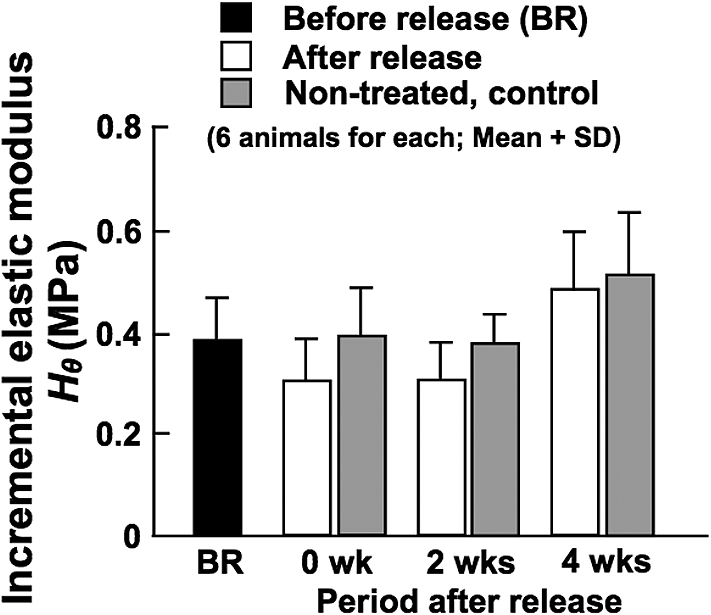
Incremental elastic modulus of the CCA determined at in vivo working blood pressure under the normal vascular smooth muscle condition before the release of constriction (BR), and soon after (0 wk) and at 2 and 4 weeks after the release of constriction. Data from age-matched, non-treated control animals are also demonstrated.
Experimental model
Arterial-venous fistula or arteriovenous shunt [ex. [7]] and arterial ligation/constriction [ex. [9]] have been commonly used to experimentally change arterial BF. The surgical procedures for the production of arterial-venous fistula/shunt are not so easy, and it is rather hard to restore BF after its reduction with this model. Therefore, no fistula/shunt models have been applied to the present purpose of BF alteration. On the other hand, Brownlee and Langille [23] applied a combined procedure of ligation and anastomosis for decreasing and recovering BF in the CCA. In the adult, male New Zealand white rabbit, they ligated the right ECA at its origin to reduce BF in the ipsilateral CCA. One month after, they transected the left CCA and performed an end-to-side anastomosis with the distal part of the right CCA for the restoration of BF in the CCA located proximally from the anastomosis. The ECA ligation decreased BF in the right CCA by 63% at 1 month after ligation, while the above-stated anastomosis immediately restored BF to control level. In contrast to such a laborious surgical procedure, our method of arterial constriction and its release is much simpler, and we could significantly reduce carotid BF by the constriction and then restore it to control level after its release. As far as the authors know, no studies except for the work reported by Brownlee and Langille [23] and the present experiments have been performed for the study on biomechanical effects of the reduction and restoration of BF. In the present study, BF in the rat CCA was decreased by 48% by 2-week arterial constriction (BR vs. BC in Fig. 1). In our previous study on the same rat artery [13], we observed a significant decrease of BF from the control level (34% decrease) at 8 weeks after the production of the same constriction as that in the present study. These results indicate that the constriction method used the present study can stably and reproducibly reduce carotid BF for a long period of time.
Arterial diameter and wall shear stress
The chronic reduction of BF in the CCA induced by constriction significantly decreased the internal diameter (BR data and Control data at 0 week in Fig. 3). After the release of constriction, the diameter progressively increased until 4 weeks, and the temporal change in diameter was in parallel with the change of BF (Fig. 2). At 4 weeks, the difference in the diameter between the experimental and age-matched control animals became insignificant similarly to the data of BF. These results indicate that it took 4 weeks for the restoration of luminal diameter and BF after the release of constriction.
In spite of significant changes in BF and luminal diameter observed after the production of arterial constriction and the subsequent release of the constriction, WSS did not change much, and there were no significant differences in the stress among all the experimental groups and age-matched controls (Fig. 4). As mentioned above, WSS is inversely related to the cube of internal diameter (see Equation (2)). Therefore, even if the change of diameter is small, it greatly affects the stress, and cancels the influence of large change in BF on WSS. Thus, WSS was kept at normal, control level between 2 and 4 Pa irrespective of BF changes. This is regarded as a phenomenon of functional adaptation and remodeling of arteries in the BF-mediated feedback system [13].
Brownlee and Langille [23] demonstrated in the rabbit that the 63% decrease of BF in the right CCA induced by the one-month ligation of the right ECA significantly decreased the internal diameter of the CCA, say up to 78% of control. Along with the decline in the diameter and BF, WSS was significantly decreased to 37–72% of control levels. This result of WSS is completely different from that obtained from the present study. The difference between the two studies is attributable to much larger BF reduction in the experiments done by Brownlee and Langille (63% after 1 month) compared with the present study (48% after 2 weeks) and also the differences of species and age (adult rabbits vs. 8-week old rats). As stated above, in our previous study, we observed a significant decrease of BF from the control level (approximately 34% decrease) at 8 weeks after the production of the same constriction as that in the present study [13]. However, there were no significant differences in the internal diameter and WSS between the low BF and the normal BF arteries. The percentage reduction of BF in the constricted CCA decreased from 48% at 2 weeks (present result) to 34% at 8 weeks (previous result), which induced approximately 30 and 5% reduction of the internal diameter, respectively, from age-matched control data. The much greater effect of diameter change on WSS compared with the effect of BF change, which is estimated from Equation (2) as already stated above, seems to have cancelled the effect of the large reduction of BF on WSS at 8 weeks after the production of constriction. In our previous study, we performed another experiment of significantly increasing BF (43%) in the CCA by the ligation of the contralateral CCA for 8 weeks, and observed an increase of internal diameter but no significant changes in WSS [13]. The increase of BF soon after the restoration of normal perfusion first produces outward remodeling, and increases WSS, although these changes were not statistically significant (Figs 3 and 4). This vascular expansion continued until the stress was restored to baseline. The homeostatic adjustment to maintain a constant, optimal WSS is effected by way of arterial enlargement in the case of increased BF. In both studies, WSS was controlled at approximately 2.5 Pa regardless of changes in BF. As these studies indicate, arteries change diameter in response to BF changes so as to maintain WSS at physiologically normal levels, which suggests a flow-mediated feedback system again [7].
Langille and O’Donnell reported that they did not observe the above-mentioned flow-mediated arterial remodeling in endothelium-denuded regions in the rabbit CCA [10], which indicates that the endothelium is essential for the homeostatic arterial response to chronic BF changes. The feedback control seems to govern the endothelial production of a number of vasoactive and mitogenic factors that play direct roles in the adaptation [ex. [24]]. Endothelial cells (ECs) are responsible for the detection of WSS, or more precisely fluid shear strain, because they are located at the interface between the blood and the vessel wall, and WSS is developed from the friction of the blood against the wall. There are several suggestions for possible mechanisms; shear stress may control the surface concentration of agonists like ATP that can be degraded at all the cell surface, shear-sensitive ion channels, the deformation of cell cytoskeleton, the mechanical deformation of transmembrane protein, and the activation of signal transduction pathways to focal adhesion sites [6]. The mechanisms for the flow-mediated arterial remodeling should be more extensively studied, for example to understand whether the phenomenon is endothelial-dependent or -independent.
Wall thickness and wall hoop stress
No significant changes were observed in the ratio of wall thickness to internal radius after the release of constriction (Fig. 5). However, the ratio seems to gradually decrease with time and reached the control level at 4 weeks when the normal BF and internal diameter were restored (Figs 2 and 3). The ratios before, soon and 2 weeks after the release of constriction were larger than those in non-treated, control arteries, possibly because of decreased internal diameter. Although there were no significant differences in BP among all the animals as mentioned above, BP slightly decreased from 103.5 ± 7.3 mmHg to 83.4 ± 12.8 mmHg by the release of constriction. On the other hand, BF significantly increased (Fig. 2), but the internal diameter remained unchanged (Fig. 3). These results suggest the increase of mean blood velocity, which may have reduced mean BP. Thereafter, the internal diameter gradually increased in parallel with BP changes (from 83.4 ± 12.8 at 0 week to 101.0 ± 4.8 mmHg at 4 weeks), which resulted in the gradual decrease of the ratio of wall thickness to internal diameter.
The wall thickness/radius ratios at 0 and 2 weeks after the release of constriction were significantly larger in comparison with age-matched controls, which corresponds to significantly smaller WHS during these periods (Fig. 6). However, the stress was restored to the control level of approximately 130 kPa at 4 weeks after the constriction release, which is considered to indicate a BP-mediated remodeling of arterial wall [14]. There are many studies demonstrating that hypertension significantly increases the thickness of arterial wall but does not change the internal diameter, resulting in the increase of the ratio of wall thickness to internal diameter [14,25]. Because of these changes in wall dimensions, WHS is not affected by the elevation of BP as estimated from Equation (3). There is evidence indicating that vascular SMCs sense such a tensile force as WHS and play important roles in BP-mediated vascular remodeling [6]. We have recently demonstrated that this feedback control is not affected by BF changes regardless of the magnitude of BP, namely both in hypertension and normotension [13].
Vascular tone and wall elasticity
Before and until 2 weeks after the release of constriction, the diameter response under the normal condition of VSM, which represents the degree of basal VSM tone, was significantly larger than in non-treated, control arteries (Fig. 7). During these periods, BF in the CCA was significantly lower than age-matched controls (Fig. 2). The diameter response reached the control level at 4 weeks after the constriction release, which corresponded to the restoration of BF. As mentioned above, we previously carried out the experiments of significantly decreasing (34%) and increasing (43%) BF in the rat CCA by the 8-week ligation of the ipsilateral ECA and the contralateral CCA, respectively, and observed no significant changes in the diameter response of the CCA under the normal VSM condition [13]. In this study, we obtained the diameter response at 2 weeks, which showed significantly greater values in the CCA exposed to low BF than in the age-matched control artery (0.021 ± 0.011 vs. 0.005 ± 0.001); the initial control value at time zero was 0.006 ± 0.004. These results were the same as those obtained from the present study. Therefore, the enhanced basal VSM tone induced by the 2-week reduction of BF was maintained for 2 weeks after the restoration of BF and returned to the normal level at 4 weeks after. The basal VSM tone enhanced by BF changes may contribute to the remodeling (ex. changes in internal diameter and wall thickness) and the functional adaptation (ex. constancies of WSS and WHS) of arterial wall. The activities of SMCs are directly reflected in basal VSM tone. On the other hand, the contractile function of SMCs was evaluated from their responses to vasoconstrictive substances such as norepinephrine [13,26]. We did not examine this response in the present study,.
Regarding wall elasticity, there was no significant influence of BF changes on the incremental elastic modulus (Fig. 8). This result was the same as our previous results which demonstrated that significant decrease and increase of BF for 8 weeks had no effects on the elastic modulus [13]. As Langille et al. suggested, no significant changes in the elastic modulus of wall material imply that there were no changes in such major wall constituents as elastin and collagen [8]. We are planning to confirm this by histologically analyzing the arteries used for the present study.
Concluding remarks
Living organs, tissues, and cells remodel by changing geometry, structure, and properties so as to functionally adapt themselves to mechanical demands [1]. For example, many studies have shown that arteries also respond to chronic changes in BF and BP and change their dimensions and properties [4,5]. In particular, there are a number of studies on arterial response to BF changes. Most of those studies have been done on the effects of either increase or decrease in BF. However, very little is known about the effects of the restoration of normal BF after reduction, whereas this BF change is clinically very important. Thus, the present study was carried out to know the dimensional and biomechanical response of arterial wall to this BF change.
In Wistar rats initially aged 8 weeks, BF in the CCA was decreased by the constriction near the distal end of the artery, and the reduced flow was maintained almost at constant for the following 2 weeks. Then, the constriction was released to restore BF. BF in the CCA was gradually increased and returned to normal level by 4 weeks after the release of constriction. Between 0 and 4 weeks after the release of constriction, the internal diameter gradually increased and the ratio of wall thickness to internal diameter decreased with the restoration of BF, and both of these returned to normal, control values at 4 weeks. Basal VSM tone represented by the diameter response under the normal condition was significantly enhanced before and until 2 weeks after the release of constriction; carotid BF was significantly lower than control data during this period. The VSM tone was returned to normal level at 4 weeks after the restoration of BF brought about by the constriction release. These results indicate that the enhancement of basal VSM tone, that appeared during the period of decreased BF after arterial constriction and also for 2 weeks after the release of constriction, induced arterial contraction, which was then released by the restoration of normal BF at 4 weeks after the removal of arterial constriction. BF changes produced by the arterial constriction and its release did not affect arterial wall elasticity, partly because there were no changes in such wall constituents as connective tissues [8], and partly because SMCs and VSM tone do not have much influence on wall elasticity [22]. Even if arterial contraction was induced by the enhancement of basal VSM tone due to stimuli developed by the reduction of BF as well as the restoration of BF, its effects on wall dimensions were of short period. And WSS was maintained at control, normal level during the processes, and WHS was returned to normal level after some period of time.
A limitation of the present study is that we did not study the structural constituents of arterial wall. As stated above, Langille et al. reported that BF changes did not change such major wall constituents as elastin and collagen, and therefore there were no significant changes in the elastic modulus of wall material [8]. However, they did not study these during the process of restoration of BF after a decrease of BF. The structural and histological studies of such wall components as elastin, collagen, and SMCs would be desirable.
Compliance with ethical standards. This work was financially supported in part by the Grant-in-Aid for Scientific Research (A) (2) (nos. 09358020 and 12308047) from the Ministry of Education, Science and Culture, Japan. The authors declare that they have no conflict of interest financially nor non-financially.
Footnotes
Acknowledgement
We greatly appreciate Dr. Hiroshi Miyazaki for his technical assistance. Moreover, one of the authors (KH) appreciates the generous support of Professor Fong Chin Su, National Cheng Kung University, Tainan, Taiwan, Professor Takashi Yamane, Kobe University, Kobe, Japan, and Professor Jiri Bursa, Institute of Solid Mechanics, Mechatronics and Biomechanics, Brno University of Technology, Czech Republic, for preparing the manuscript.