
Abstract
BACKGROUND:
Cartilage tissue lacks the ability to heal. Cartilage tissue engineering using cell-free scaffolds has been increasingly used in recent years.
OBJECTIVE:
This study describes the use of a type I collagen scaffold combined with WNT5A plasmid to promote chondrocyte proliferation and differentiation in a rabbit osteochondral defect model.
METHODS:
Type I collagen was extracted and fabricated into a collagen scaffold. To improve gene transfection efficiency, a cationic chitosan derivative N,N,N-trimethyl chitosan chloride (TMC) vector was used. A solution of TMC/WNT5A complexes was adsorbed to the collagen scaffold to prepare a WNT5A scaffold. Osteochondral defects were created in the femoral condyles of rabbits. The rabbits were divided into defect, scaffold, and scaffold with WNT5A groups. At 6 and 12 weeks after creation of the osteochondral defects, samples were collected from all groups for macroscopic observation and gene expression analysis.
RESULTS:
Samples from the defect group exhibited incomplete cartilage repair, while those from the scaffold and scaffold with WNT5A groups exhibited “preliminary cartilage” covering the defect. Cartilage regeneration was superior in the scaffold with WNT5A group compared to the scaffold group. Safranin O staining revealed more proteoglycans in the scaffold and scaffold with WNT5A groups compared to the defect group. The expression levels of aggrecan, collagen type II, and SOX9 genes were significantly higher in the scaffold with WNT5A group compared to the other two groups.
CONCLUSIONS:
Type I collagen scaffold showed effective adsorption and guided the three-dimensional arrangement of stem cells. WNT5A plasmid promoted cartilage repair by stimulating the expression of aggrecan, type II collagen, and SOX9 genes and proteins, as well as inhibiting cartilage hypertrophy.
Introduction
Cartilage lacks the ability to heal, which makes treatment of traumatic and degenerative cartilage defects a challenge for orthopedic surgeons [1,2]. Surgical treatments for these defects, such as abrasion, microfracture, and autologous chondrocyte implantation, do not promote the formation of normal cartilage tissue, and may result in joint damage over time [3]. Tissue engineering to create cartilage is a promising technique for the treatment of cartilage defects. Current tissue engineering techniques for cartilage repair include the use of biocompatible scaffolds and stem cells. However, there are some limitations to the clinical use of conventional tissue engineering. The isolation and culture of stem cells is complex, and requires two surgeries. In situ cartilage tissue engineering simplifies the conventional tissue engineering process; it involves microfractures at the site of cartilage defect, which stimulate autologous bone marrow stem cells to repair the defect. This technique requires the presence of a biological scaffold and cytokines.
Collagen scaffolds are widely used because of their biocompatibility and degradability. Previous studies reported that human stem cells bind to type I collagen through various protein receptors, and type I collagen promotes osteogenic differentiation of stem cells [4,5]. Mizuno et al. [6] reported that integrins in type I collagen induce osteoblastic differentiation of bone marrow cells. Collagen was also reported to effectively control the release of growth factors [7]. Therefore, in this study, we used type I collagen to fabricate scaffolds to promote cartilage repair.
Cytokines induce the differentiation of stem cells into chondrocytes, and promote cartilage repair. However, because there is no blood supply to the cartilage defect, few nutrients are available for cartilage repair. Growth factors can modify the local environment and promote cartilage regeneration. Previous studies reported that only embryonic stem cells implanted in the cartilage defect (without cytokines) proliferated and hypertrophied; however, the regenerated cartilage did not integrate well with the normal surrounding cartilage [8–10]. Various cytokines may stimulate stem cell differentiation, such as bone morphogenic proteins (BMPs), transforming growth factor-β, and fibroblast growth factor. BMP-2 was widely used to promote stem cell differentiation to chondrocytes, but instead caused chondrocyte hypertrophy [11–14]. Transforming growth factor-β3 inhibited the expression of type I collagen, a marker of fibrocartilage, but did not prevent stem cell hypertrophy [15]. Parathyroid hormone-related peptide inhibited excessive chondrocyte proliferation, but also stem cell differentiation to chondrocytes [16], which reduces its applicability for cartilage regeneration. None of the currently available cytokines are ideal for the promotion of stem cell differentiation to articular chondrocytes. An ideal cytokine should enhance articular cartilage neogenesis by stimulating stem cells, without causing significant chondrocyte hypertrophy. WNT5A inhibits chondrocyte hypertrophy [17–19]. In another study, WNT5A inhibited the BMP-2-induced differentiation of human embryonic stem cells into chondrocytes, and chondrocyte maturation, thus suppressing the formation of fibrocartilage and articular cartilage [12]. Therefore, WNT5A plays an important role in stem cell differentiation to hyaline cartilage.
In this study, we introduced the use of a type I collagen scaffold combined with WNT5A plasmid to promote chondrocyte proliferation and differentiation in a rabbit model of osteochondral defects.
Materials and methods
Ethical approval
The experiments were approved by the Animal Ethics Committee of the Second Affiliated Hospital of Zhejiang University School of Medicine, China (no. 2020-46). The animals were obtained from the Zhejiang Academy of Medical Sciences (Zhejiang, China), and maintained at the Second Affiliated Hospital of Zhejiang University School of Medicine.
Collagen extraction and scaffold fabrication
Type I collagen was extracted from pig tendons obtained from butchers. The pig tendons were dissected, sliced, and washed with distilled water to remove plasma proteins. The tendons were then immersed in 0.5 M acetic acid (1.0% weight/volume), which was continuously stirred overnight at 4 °C; the aqueous extract was then obtained. The extract was precipitated with saline (0.9% NaCl) and dissolved in 0.5 M acetic acid. The solution was dialyzed with distilled water, frozen, and freeze-dried to obtain collagen. The collagen was dissolved in 0.05 M acetic acid to prepare a 10 mg/mL solution. The solution was poured into a mold (diameter: 3 cm; depth: 4 mm), which was placed in a fridge at −80 °C. The solution was then freeze-dried and sheared, and type I collagen scaffolds were fabricated. The scaffolds were elastic, and could be cut with scissors or a sharp knife to match the cartilage defect area. The scaffolds were sterilized with ethylene oxide in preparation for the experiments.
Preparation of N,N,N-trimethyl chitosan chloride/DNA complexes
N,N,N-trimethyl chitosan chloride (TMC) was dissolved in phosphate-buffered saline to prepare a 5 mg/mL solution. WNT5A DNA was dissolved in phosphate-buffered saline to prepare a 1 mg/mL solution. TMC and WNT5A DNA were filtered through a 0.22-μm filter for sterilization, and 0.2 mL DNA solution was added to 1 mL TMC solution. This solution was mixed using a vortex mixer for 30 s and incubated for 30 min at 37 °C. In previous studies, TMC/DNA complexes had the highest transfection efficiency in vitro when the theoretical charge ratio (molar ratio between amine and phosphate groups), denoted by the nitrogen/phosphate ratio, was fixed at 10 [20,21]. The solution containing TMC/DNA complexes was adsorbed onto the collagen scaffold to prepare a WNT5A scaffold.
Rabbit osteochondral defect model
After injecting 60 mg/kg of 0.7% pentobarbital sodium into the ear vein to induce general anesthesia, osteochondral defects were created in the femoral condyles of the bilateral hind limbs of 12 young adult female New Zealand White rabbits (mean weight: 2.5 ± 0.2 kg). Animals received subcutaneous carprofen (1.5 mg/kg) for analgesia after the surgery. The skin over the knees of the rabbits was shaved and disinfected with 5% Anerdian III. A midline longitudinal incision was made, and the knee joint was exposed using a medial parapatellar approach. The patella was dislocated laterally, and the knee was flexed at 90° to expose the non-weight-bearing area of femoral condyles. A steel drill with a diameter of 4 mm was used to create an osteochondral defect that extended to the subchondral bone, located in the center of the anterior aspect of the femoral condyle.
All rabbits were randomly divided into three groups (group 1: defect group; group 2: scaffold group, group 3: scaffold with WNT5A group). Each group comprised four rabbits (eight knee joints). In group 1, rabbits did not receive any special treatment. In group 2, a collagen scaffold of suitable size was implanted into the cartilage defect. In group 3, a collagen scaffold loaded with WNT5A plasmid was implanted into the cartilage defect.
The patella was relocated, the joint capsule was sutured using 4-0 Monocryl (Ethicon, Inc., Raritan, NJ, USA), and the skin was sutured using 5-0 Monocryl (Ethicon, Inc.). Animals were monitored by technicians during the surgery. All rabbits were allowed to move and feed freely. Two rabbits from each group were humanely killed using inhalational anesthesia 6 weeks after the surgery. Two other rabbits from each group were killed at 12 weeks. The knees were harvested for histological analysis.
Gross morphology and histology
Samples from the knees were evaluated using the macroscopic assessment scale for cartilage repair of the International Cartilage Repair Society [2]. The samples were fixed with 4% paraformaldehyde for 1 week and continuously decalcified with 10% ethylenediaminetetraacetic acid for 4 weeks. The ethylenediaminetetraacetic acid solution was changed every week. The samples were shaped so that the size was appropriate for the cartilage defect, and excess tissue was discarded. The samples were embedded in paraffin, cut into 7-μm sections, stained with hematoxylin and eosin to observe the distribution of cells, and then stained with safranin O to observe glycosaminoglycans.
Quantitative real-time reverse-transcription polymerase chain reaction
To detect chondrogenic gene expression, we collected total RNA from cartilage cells using the RNeasy Mini Kit (Qiagen, Hilden, Germany); 1 mg of the total RNA was reverse-transcribed into complementary DNA using the Omniscript RT kit (Qiagen). The expression levels of the target genes (Table 1) were analyzed using quantitative real-time reverse-transcription polymerase chain reaction (PCR). Real-time PCR was performed using the Applied Biosystems 7300 real-time PCR system (Thermo Fisher Scientific, Inc., Waltham, MA, USA) and SYBR Green PCR master mix (Thermo Fisher Scientific, Inc.) for 15 s at 90 °C and 60 s at 60 °C, and the fluorescence intensity was recorded for 40 cycles. All results were normalized to glyceraldehyde 3-phosphate dehydrogenase expression. The relative expression levels were calculated by comparison between the test and control groups using the 2−ΔCT method [22].
Real-time PCR primers for target genes
Real-time PCR primers for target genes
∗ S = Sense; A = Antisense. PCR: polymerase chain reaction.
The expression levels of target genes at 6 and 12 weeks after the operation were analyzed using the Wilcoxon signed-rank test. Post-hoc Bonferroni test was used to compare the data among the groups at the various time points. The level of significance was set at p < 0.05.
Results
Scaffold observation
Figure 1 demonstrates the structure and morphology of the scaffolds before treatment. Scanning electron microscopy revealed a microporous, sponge-like structure. The microporous structure facilitates adsorption of bone marrow cells and cytokines into the subchondral bone, and guides the three-dimensional arrangement of bone marrow cells.
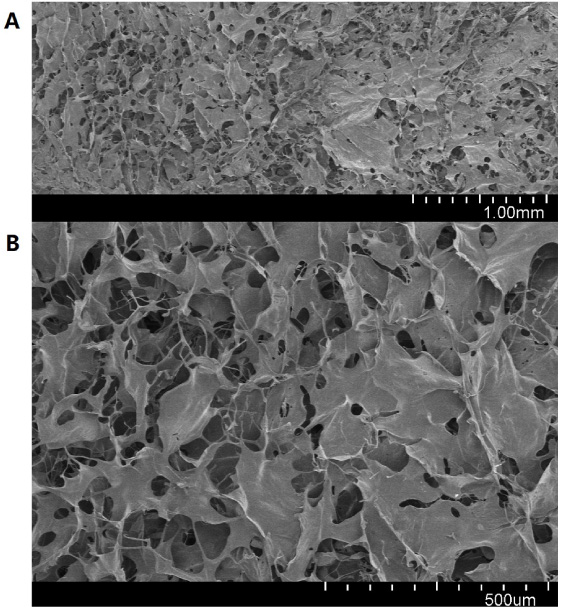
Scanning electron microscopy images of collagen scaffold, without treatment and before implantation, showing a nanoporous structure.
All rabbits recovered well and remained active after surgery, with no evidence of wound infection or pain. All joints showed full range of motion. Figure 2 displays images of explanted knees at 6 and 12 weeks after the surgery. These images demonstrate differences in color, consistency, and macroscopic repair at the defect site and surrounding cartilage among the groups. The surrounding cartilage did not show evidence of inflammation, damage, or erosion. Six weeks after surgery, samples from the defect group exhibited incomplete cartilage repair, with only small amounts of cartilage around the defect. Samples from the scaffold and scaffold with WNT5A groups exhibited “preliminary cartilage” covering the defect. Twelve weeks after surgery, samples from the defect group exhibited inadequate repair and a rough surface. Samples from the scaffold group exhibited regenerative tissue, which was raised above the joint surface and appeared white compared to the surrounding cartilage. Samples from the scaffold with WNT5A group exhibited adequate cartilage.
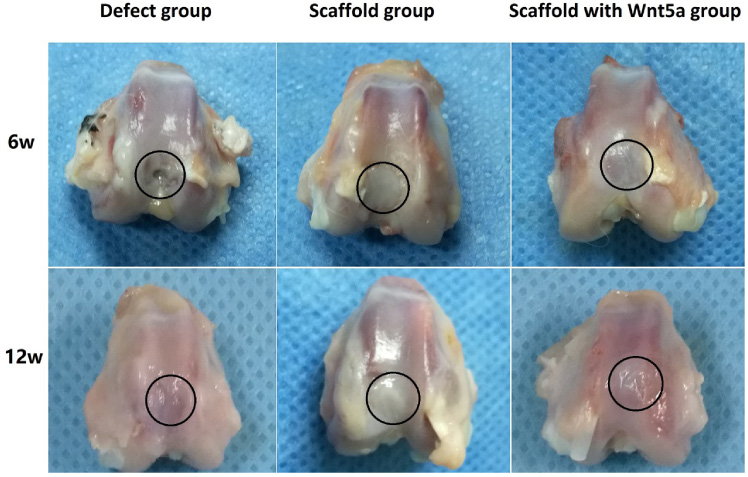
Photographs of rabbit knee specimens from the defect, scaffold, and scaffold with WNT5A groups 6 and 12 weeks after the operation. The cartilage defect is indicated by a black circle. None of the samples exhibited evidence of joint erosion in the surrounding cartilage. Defect group: macroscopic appearance demonstrating incomplete cartilage repair. Scaffold group: the repaired cartilage appeared white and was slightly raised compared to the surrounding cartilage. Scaffold with WNT5A group: regenerated cartilage appeared similar in color, consistency, and smoothness to the surrounding cartilage.
Figure 3 summarizes the results of histological observations. Six weeks after surgery, samples from the defect group exhibited thinner regenerative tissue and a disordered arrangement of cells. There were more cells in the defect compared to scaffold and scaffold with WNT5A groups. Safranin O staining demonstrated more proteoglycans in the scaffold compared to defect group. Twelve weeks after surgery, a greater degree of cartilage repair and more proteoglycans were observed in the scaffold and scaffold with WNT5A groups compared to the defect group. Cartilage hyperplasia was observed in the scaffold group. The regenerated cartilage tissue was at the same level as the surrounding cartilage tissue in the scaffold with WNT5A group. Samples from the scaffold with WNT5A group exhibited the greatest degree of repair, followed by the scaffold and defect groups.
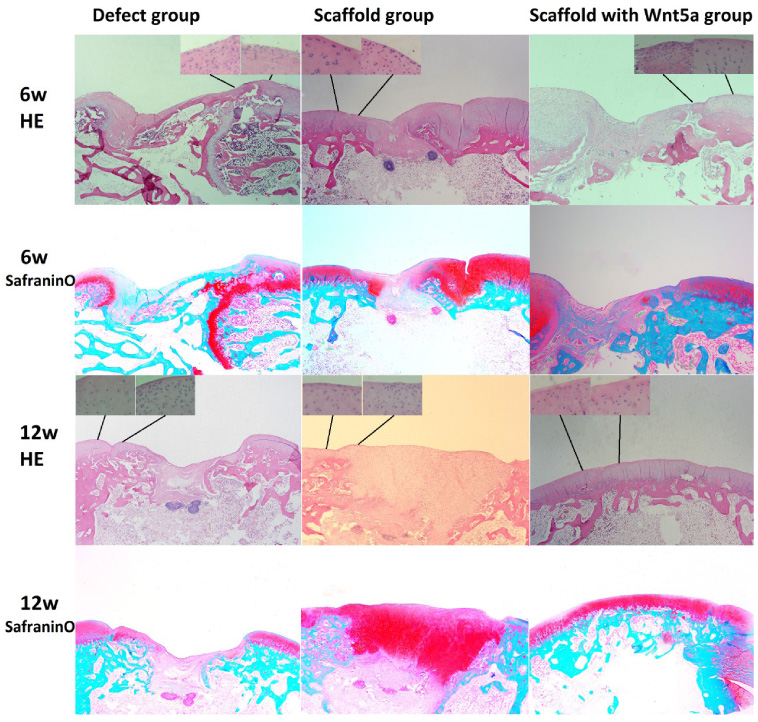
Representative histological images. Magnified images of cell morphology near the junction between the regenerated and normal cartilage, stained with hematoxylin and eosin. Representative images of cartilage sections from the defect, scaffold, and scaffold with WNT5A groups, stained with hematoxylin and eosin and safranin O (proteoglycan staining). Sections were obtained approximately midway and perpendicular to the defect site. Proteoglycans were detected in all repaired tissues. Less cartilage regeneration was observed in the defect group.
The gene expression levels of aggrecan, collagen type II, and SRY-related high-mobility group box 9 (SOX9) were significantly higher in the scaffold with WNT5A group compared to the other two groups, at both 6 and 12 weeks (Fig. 4). The gene expression levels did not differ between 6 and 12 weeks.
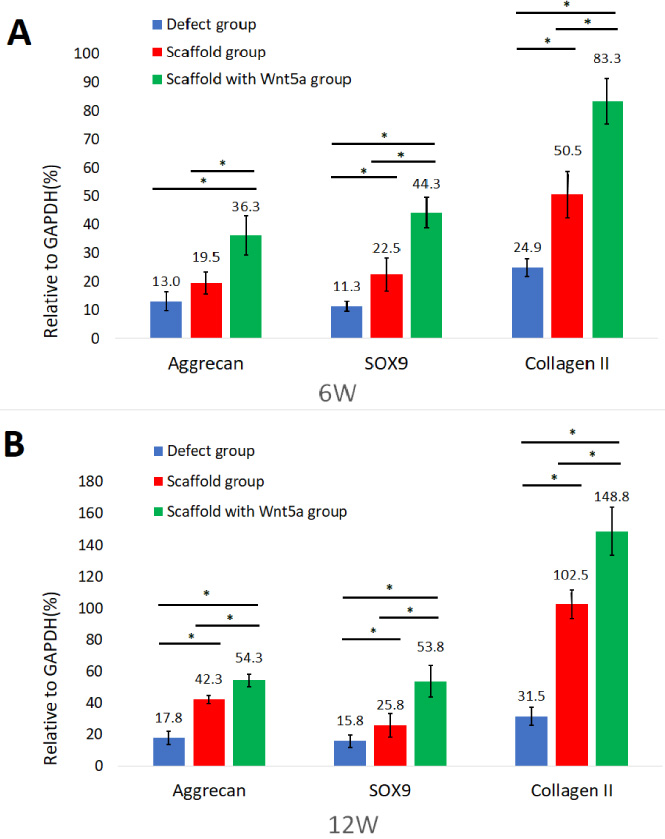
Gene expression profiles of tissues harvested at 6 and 12 weeks after transplantation using quantitative real-time reverse-transcription polymerase chain reaction. The gene expression levels were normalized to that of glyceraldehyde 3-phosphate dehydrogenase. Values are presented as means ± standard deviations for the three experiments. ∗Indicates p < 0.05.
Combinations of scaffolds and cells have been investigated in preclinical studies. However, limitations to cell culture techniques remain. Microfractures promote autologous stem cell recruitment, but this technique alone does not lead to satisfactory cartilage repair. Previous clinical studies have reported failed healing during long-term follow up after microfracture surgery [23,24]. The regenerated cartilage tissue after microfracture consisted of fibrocartilage, rather than hyaline cartilage, and showed a disordered cell arrangement; this may explain the failure of microsurgery in previous studies. The regenerated tissue had poor mechanical properties, resulting in degeneration of the cartilage defect. Fibrocartilage may form because of insufficient recruitment of stem cells to the cartilage defect [25], a lack of effective stimulation during chondrocyte differentiation, and local infiltration of fibroblasts. An ideal scaffold would adsorb the stem cells, thus preventing fibroblast invasion and fibrocartilage formation. Therefore, cell-free scaffolds are gaining popularity, and studies of these scaffolds have reported promising results [26,27]. Cartilage regeneration occurs due to migration of undifferentiated stem cells from the subchondral bone into the defect, which induces cartilage formation [26–28]. Type I collagen binds to stem cells and promotes osteogenic differentiation [29]. A clinical trial that involved the application of cell-free collagen scaffold to repair knee joint cartilage demonstrated satisfactory outcomes at the 2-year follow-up [27]. Although a subsequent follow-up study demonstrated increased degenerative wear of the repaired cartilage and a frequent need for revision surgery, the results demonstrated the potential of collagen scaffolds for cartilage repair [30].
In this study, a type I collagen scaffold was fabricated. The porous structure of this scaffold can absorb bone marrow stem cells from the cartilage defect and guide the three-dimensional arrangement of stem cells, which supports chondrocyte proliferation [31,32]. We used different sizes of the microporous collagen scaffold (range: 100–300 μm). Previous in vitro experiments demonstrated that scaffolds with smaller pores (100–300 μm) were associated with greater chondrocyte differentiation [33]. Moreover, scaffolds with smaller pores (150–300 μm) were more conducive to both chondrocyte growth and differentiation compared to larger pores (300–500 μm) [34]. Scaffolds with small porous structures (approximately 100 μm) create a hypoxic environment, which promotes chondrocyte formation [35]. Replacing the cartilage defect with collagen scaffolds prevents matrix leakage and provides a functional matrix facilitating chondrocyte proliferation [36]. In the current study, cartilage repair and proteoglycans were greater in the scaffold compared to defect group, as revealed by safranin O staining at 12 weeks after surgery. The mechanical properties of regenerated cartilage depend on the content of proteoglycans, which are the main proteins in hyaline cartilage [37]. In summary, the results of this study suggest that the application of type I collagen promotes cartilage restoration.
In this study, TMC/WNT5A complexes were integrated into the scaffolds, which resulted in satisfactory cartilage repair. More advantageous local gene expression is observed with gene therapy compared to the classic tissue engineering technology. A greater quantity of DNA is inserted with local application of growth factors compared to proteins, which makes the application of growth factors more stable. To achieve continuous gene expression, DNA plasmids may be transfected into seed cells. Non-viral transfection methods, including polymers and liposomes, are safe and efficient, and have a low risk of immunogenicity [38]. In this study, TMC/DNA complexes were used for cartilage regeneration because they encapsulate large quantities of plasmid DNA and have non-toxic degradation products. These complexes mask the negatively charged DNA, thereby facilitating their uptake across the cell membrane and increasing transfection efficiency [39].
Previous studies reported that WNT5A promoted early chondrogenesis and inhibited chondrocyte hypertrophy [17–19]. In the WNT5A/Ca+2 pathway, WNT5A induced SOX9 generation through the CaMK/NFAT pathway, thereby promoting chondrocyte production. SOX9 proteins are important transcriptional factors that regulate cartilage formation. Studies of heterozygous SOX9-mutant mouse embryos in different stages confirmed that SOX9 inhibited chondrocyte hypertrophy [40]. In the current study, the WNT5A gene was selected as a cytokine to promote early chondrogenesis and inhibit chondrocyte hypertrophy. Compared to the regenerated cartilage in the scaffold group, that in the scaffold with WNT5A group appeared more similar to normal cartilage. Previous studies reported that WNT5A promoted chondrocyte proliferation [41]. In the current study, increased expression of type II collagen and aggrecan in the scaffold with WNT5A group suggested greater regeneration of cartilage, with components similar to the normal hyaline cartilage [42,43]. Therefore, WNT5A affects chondrocyte differentiation. This study demonstrated that type I collagen scaffolds with WNT5A plasmid promote cartilage repair and reconstruction of the anatomical interface of cartilage.
Conclusions
In this study, type I collagen scaffolds with WNT5A plasmid were designed and used for osteochondral repair in a rabbit model. In addition, the biological properties of the scaffold were evaluated. Type I collagen scaffold effectively adsorbed and guided the three-dimensional arrangement of stem cells. WNT5A plasmid promoted cartilage repair by stimulating the expression of aggrecan, type II collagen, and SOX9 genes and proteins, as well as inhibiting cartilage hypertrophy. In conclusion, type I collagen scaffolds with WNT5A plasmid show promise for cartilage repair.
Footnotes
Acknowledgements
This study was supported by grants from the National Natural Science Foundation of China (nos 81772325 and 82072413).
Conflict of interest
The authors declare no conflict of interest.