
Abstract
BACKGROUND:
In the pathogenesis of atherosclerotic cardiovascular disorders, vascular endothelium is crucial. A critical step in the development of atherosclerosis is endothelial dysfunction. Furin may play a factor in vascular remodeling, inflammatory cell infiltration, regulation of plaque stability, and atherosclerosis by affecting the adhesion and migration of endothelial cells. It is yet unknown, though, how furin contributes to endothelial dysfunction.
METHODS:
We stimulated endothelial cells with oxidized modified lipoprotein (ox-LDL). Endothelial-to-mesenchymal transition (EndMT) was found using immunofluorescence (IF) and western blot (WB). Furin expression level and Hippo/YAP signal activation were found using reverse transcription-quantitative PCR (RT-qPCR) and WB, respectively. To achieve the goal of furin knockdown, we transfected siRNA using the RNA transmate reagent. Following furin knockdown, cell proliferation, and migration were assessed by the CCK-8, scratch assay, and transwell gold assay, respectively. WB and IF both picked up on EndMT. WB and RT-qPCR, respectively, were used to find furin’s expression level. We chose the important micrornas that can regulate furin and we then confirmed them using RT-qPCR.
RESULTS:
EndMT was created by ox-LDL, evidenced by the up-regulation of mesenchymal cell markers and the down-regulation of endothelial cell markers. Furin expression levels in both protein and mRNA were increased, and the Hippo/YAP signaling pathway was turned on. Furin knockdown dramatically reduced the aberrant migration and proliferation of endothelial cells by ox-LDL stimulation. Furin knockdown can also suppress ox-LDL-induced EndMT, up-regulate indicators of endothelial cells, and down-regulate markers of mesenchymal cells. After ox-LDL stimulation and siRNA transfection, furin’s expression level was up-regulated and down-regulated.
CONCLUSION:
Our study demonstrated that furin knockdown could affect ox-LDL-induced abnormal endothelial cell proliferation, migration, and EndMT. This implies that furin plays an important role in endothelial dysfunction.
Introduction
Arteriosclerotic (AS) is the leading cause of acute cardiovascular events worldwide [1]. Endothelial dysfunction, as the initial link of atherosclerosis, has attracted much attention [2]. In healthy individuals, the endothelium responds to a range of chemical and biomechanical signals by secreting factors that regulate vascular tone, smooth muscle cell proliferation and migration, immune cell adhesion, thromboresistance, and vascular inflammation [3, 4]. In contrast, endothelial dysfunction initiates atherosclerotic progression by promoting inflammation, lipoprotein oxidation, platelet aggregation, and thrombosis [5], which triggers the production of cell adhesion molecules, the migration of macrophages under the intima of blood vessels and endothelial cell injury [6–10]. Oxidized modified lipoprotein (ox-LDL) is believed to play a key role in endothelial dysfunction [11].
One of the key processes in endothelial dysfunction, endothelial-to-mesenchymal transition (EndMT), is the process by which endothelial cells acquire a mesenchymal phenotype [12]. By tracking Cre-lox endothelial profiles in mice and examining human plaques, EndMT-derived fibroblast-like cells in endothelial atherosclerotic plaques are common and the unstable and rupture-prone properties of plaques may be closely related to EndMT [13]. Furin, a member of the protein convertase family, can activate TGF-β by cleaving the precursor of TGF-β [14]. TGF-β is a major regulator of EndMT [15]. It has been found that in both human atherosclerotic type cells and mouse atherosclerotic in human atherosclerosis-type cells and mouse atherosclerosis specimens, furin expression was significantly upregulated [16, 17], suggesting that furin may be associated with EndMT in atherosclerotic endothelial dysfunction. However, it is not clear whether furin directly affects the mechanism of atherosclerosis through its involvement in EndMT and endothelial dysfunction.
Yes-associated protein 1 (YAP1) is a transcriptional cofactor and effector of the Hippo pathway, and its activity is critical for cell proliferation and vascular development [18]. Studies have found that SMAD7, a downstream protein of TGF-β, interacts with YAP1 [19]. Furin is closely related to TGF-β. However, whether YAP1 is downstream of it and thus involved in the regulation of EndMT remains to be clarified. miRNAs are able to reverse regulate transcription by inhibiting translation or degradation of proteins [20], for example, miR-133a, and miR-22-3p can inhibit furin expression [21, 22]. Therefore, the possibility that miRNAs are involved in furin regulation of EndMT as upstream of furin also needs to be considered.
In the present study, we treated ECs with ox-LDL to investigate whether it leads to the formation of EndMT and whether the protein furin as well as the hippo/yap signaling pathway are involved in this process. We further explored the efficacy of knocking down furin in alleviating the process of EndMT. We also conducted preliminary exploration on whether there are microRNAs upstream of furin that regulate furin. The study of the potential mechanisms of the above processes will not only deepen the understanding of the pathogenesis of atherosclerosis but also contribute to the discovery of therapeutic targets that effectively inhibit the alleviation of atherosclerosis.
Material and methods
Cell culture and treatments
Experiments were performed using primary cardiac microvascular endothelial cells (CMVECs), which are given away from the Chinese Academy of Sciences cell bank (Shanghai, China). CMVECs were cultured in high-glucose DMEM (Gibco, Shanghai, China) containing 10% fetal bovine serum (FBS; Sijiqing Bioengineering Material Co., Ltd, Zhejiang, China) and 1% penicillin/streptomycin (Beyotime, Shanghai, China) in a man-made environment of 5% CO2 at 37°C. The culture medium was replaced every 1–2 days, and cells at 85–90% confluence were passaged at a ratio of 1 : 3 confluence. All of the cells were seeded in the tissue culture plates. The cells were used between passages two and five in all experiments. CMVECs of 75% confluence were incubated with various concentrations of ox-LDL (100 μg/mL; Yiyuan Biotech. Co. Ltd, Guangdong, China) for 24 h. In subsequent functional assay Ox-LDL was used to establish the model.
CMVECs were incubated in 96/24/12/6 well plates after trypsin digestion. After the cells were attached to the wall, ox-LDL was used to establish the cell model for 24 h according to the previous experimental data. The treated cells were rinsed with PBS and prepared for some experiments.
Cell viability assay
The cell viability of CMVECs was detected by the CCK-8 kit according to previous reports. To put it simply, CMVECs were seeded on 96-well plates and treated with ox-LDL (100 μg/mL), with siRNA or negative control (10 nM) for 24 h. Then, CCK-8 was mixed with a medium at a ratio of 1 : 10 and added to each well for 1–2 h incubation at 37°C. Next up, absorbance was measured at 450 nm by EnSpire (PerkinElmer, USA). For per experiment, 3 duplicate wells were examined for each group.
Immunofluorescence
The stimulated CMVECs were seeded on 24-well plates rinsed with phosphate-buffered saline (PBS) three times for five minutes and then fixed with 4% paraformaldehyde for 30 min, and then were permeabilized with 0.3% Triton X-100 in PBS for 30 min at room temperature. The cells were treated with 10% goat serum for 30 minutes at room temperature to block the particular antibody. Primary antibodies were diluted 1 : 800 for α-SMA (Abcam ab124964) and diluted 1 : 1000 for ve-cadherin (Abcam ab205336) in PBST (0.1%) with 1% BSA. The samples were then incubated at 4°C for an entire night.. The cells were treated with goat anti-rabbit IgG H&L Alexa Fluor® 488 for 30 minutes following three PBS washes. After three PBS washes, the cells were stained with DAPI for five minutes at room temperature. The fluorescence microscope (80i, Nikon, Tokyo, Japan) was used to capture the fluorescent images. The IMAGE-PRO PLUS 6.0 (Version 6.0, Media Cybernetics, Bethesda, Maryland, USA) was used to evaluate the data. Transwell Migration Assay.
A 24-well transwell plate (SPL Life Sciences, Korea) was used to seed CMVEC cells (104 cells/well) into the upper chamber. The growth media was then switched to a starvation medium for another 24 hours. After twice cleaning the well with PBS, fix it for 15 minutes with 4% paraformaldehyde. A membrane with hole sizes of 8 μm is traversed by migratory cells on the underside. Using a cotton swab, nonmigratory cells were eliminated from the upper chamber. Migratory cells were then counted and stained for ten minutes using Crystal Violet Staining Solution.
Wound migration assay.
A 6-well culture multi-plate was seeded with 104 CMVECs per well, and the cells were grown for 48 hours until confluence. With a 200 uL pipette tip, one perpendicular scratch was formed in each well, and the samples were cultured for 24 hours with normal DMEM, ox-LDL, ox-LDL with siRNA, or ox-LDL with NC. Axio Observer equipment was used to capture the images, which were then processed using ImageJ software 24 hours later.
RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR).
Following the manufacturer’s recommendations, total RNA from CMVECs was extracted using Trizol reagent, combined with 0.2 mL of chloroform (Beyotime, Shanghai, China), and centrifuged at 12,000 xg for 15 minutes at 4°C. After plus 0.4 mL of 100% isopropyl alcohol, waiting for 10 minutes at 4¡?centrifuge at 12,000 xg for 10 minutes at 4°C. The extracted RNA pellets were washed with 75% ethanol and centrifuged at 7,500×g for 5 min at 4°C. After dissolving the dried pellets in 20 μL of water treated with diethylpyrocarbonate (DEPC), RNA was measured and verified using Nanodrop 2000 (Thermo Fisher Scientific, MA, USA). Complementary DNA (cDNA) was prepared from RNA using an Evo M-MLV RT kit (Accurate Biology, China). The RNA levels were ascertained using a quantitative real-time polymerase chain reaction (qRT-PCR). A 384-well qRT-PCR plate was filled with the used primers after they had been combined with distilled water. After adding the template (cDNA) and SYBR green (Accurate Biology, China), the PCR equipment (lightcycler 480II, Rohce, USA) was used to validate the results. The specific primer sequences are shown in Table 1.
Primers for RT-qPCR
Primers for RT-qPCR
Primers for micrna
Cell lysis buffer for Western and IP, which contains proteinase and phosphatase inhibitors (Beyotime, Shanghai, China), was used to scrape the cells in order to isolate the proteins from the CMVECs. The cells were then incubated for five minutes in an ice container. The supernatants were collected in a fresh tube and protein concentrations were measured using a bicinchoninic acid assay kit (BCA kit; Beyotime, Shanghai, China) after centrifuging at 10,000×g for 5 min at 4°C. For ten minutes, the entire cellular protein was boiled at 98°C to denature the fractions. Western blotting was done to verify protein expression. Proteins from the lysate were sorted into equal parts using polyacrylamide gel electrophoresis at 10% or 12% sodium dodecyl sulfate. The proteins were then transferred to PVDF membranes, which were treated overnight at 4°C with suitable diluted primary antibodies (Table 1).
The membranes were washed three times with tris buffered saline (TBS) containing 0.1% Tween 20 before being incubated with secondary antibodies for 1 hour at room temperature. BeyoECL Plus (Beyotime, Shanghai, China) was used to create membranes via improved chemiluminescence.
Statistical analysis
All findings are given as Mean±SD. The Student’s t-test was used to make comparisons between the two groups. ANOVA was used to evaluate comparisons within various groups, followed by the Dunnet t-test. Differences with a P value of 0.05 were considered significant. All of the tests were carried out at least three times.
List of primary antibodies used for western blots
List of primary antibodies used for western blots
ox-LDL induced EndMT in CMVECs
To mimic lipid accumulation-disrupted normal functionality of the endothelium endothelial in atherosclerosis, we treated CMVECs with 100 μg/mL doses of ox-LDL for 24 h. Immunofluorescence staining showed that incubation of CMVECs with ox-LDL decreased expression of the endothelial marker VE-cadherin (Fig. 1A and 1C) and increased expression of the mesenchymal marker α-SMA (Fig. 1B and 1D). Consistently, western blot showed that. (Fig. 1E and 1F).
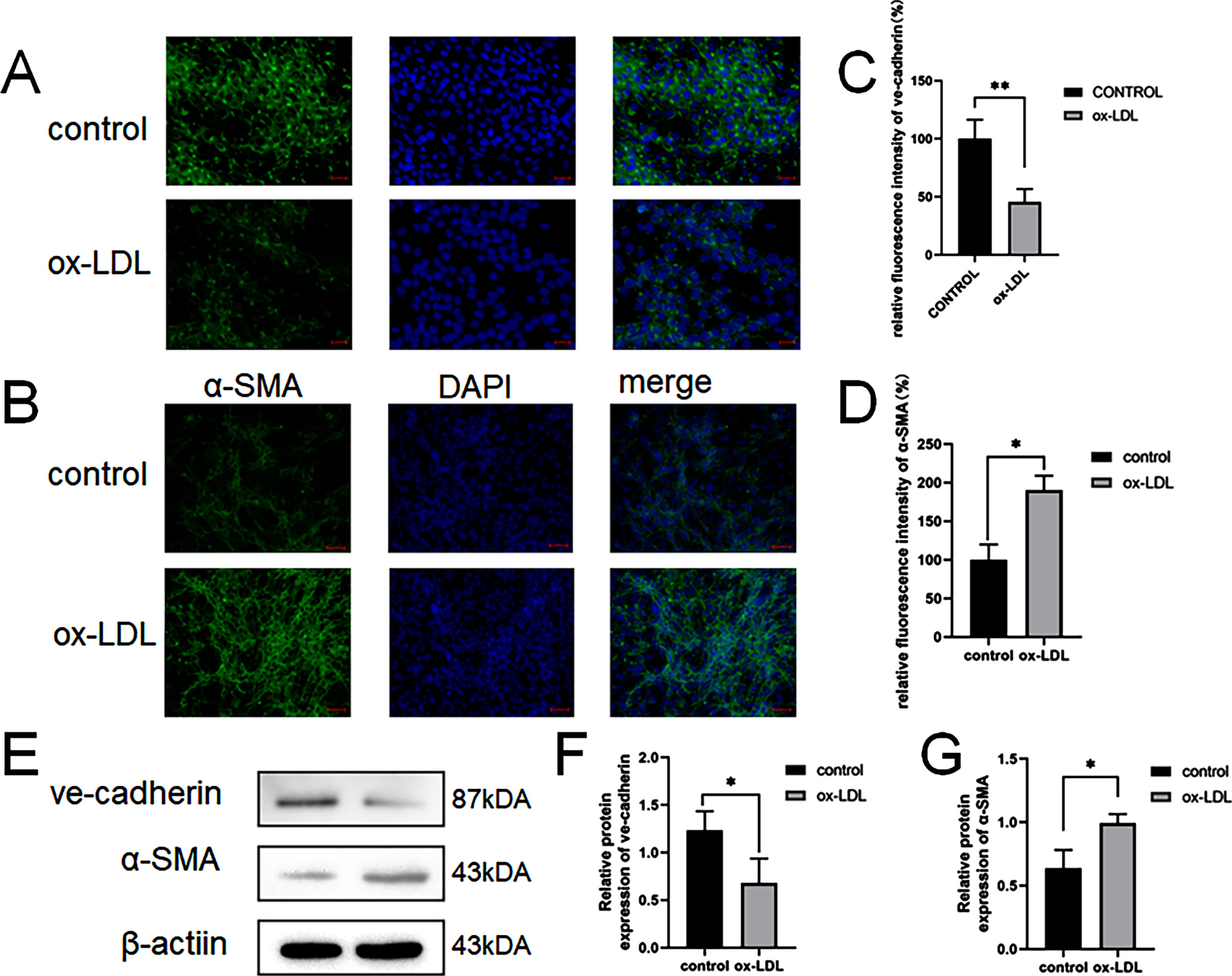
Immunofluorescence analysis of the protein expression of ve-cadherin and α-SMA.
Scale bar: 100 μm. (A, C) Quantification of Immunofluorescence analysis of ve-cadherinand α-SMA. CMVECs were treated ox-LDL (100 μg/mL) for 24 h. (B, D) The western blot analysis results revealed that the expression of EndMT molecules induced by ox-LDL. (E). Western blots indicating total ve-cadherin, α-SMAand β-Actin levels in whole cell lysates (FandG). Two asterisks (**) denote P value < 0.01, and one asterisk (*) denotes P value < 0.05 by one-way ANOVA analysis (n = 3, >1,000 cells per trial).
In addition, we used CMVEC cells, incubated with ox-LDL, to establish an in vitro AS model. We found that the furin protein and mRNA expression was significantly increased in the ox-LDL group as compared with the control group (Fig. 2a, b). To further explore the mechanism of furin in AS, the activation of the Hippo/Yap pathway was investigated. As shown in Fig. 2, the protein expressions and mRNA of yap, lats were significantly increased in the ox-LDL group compared with the control group.
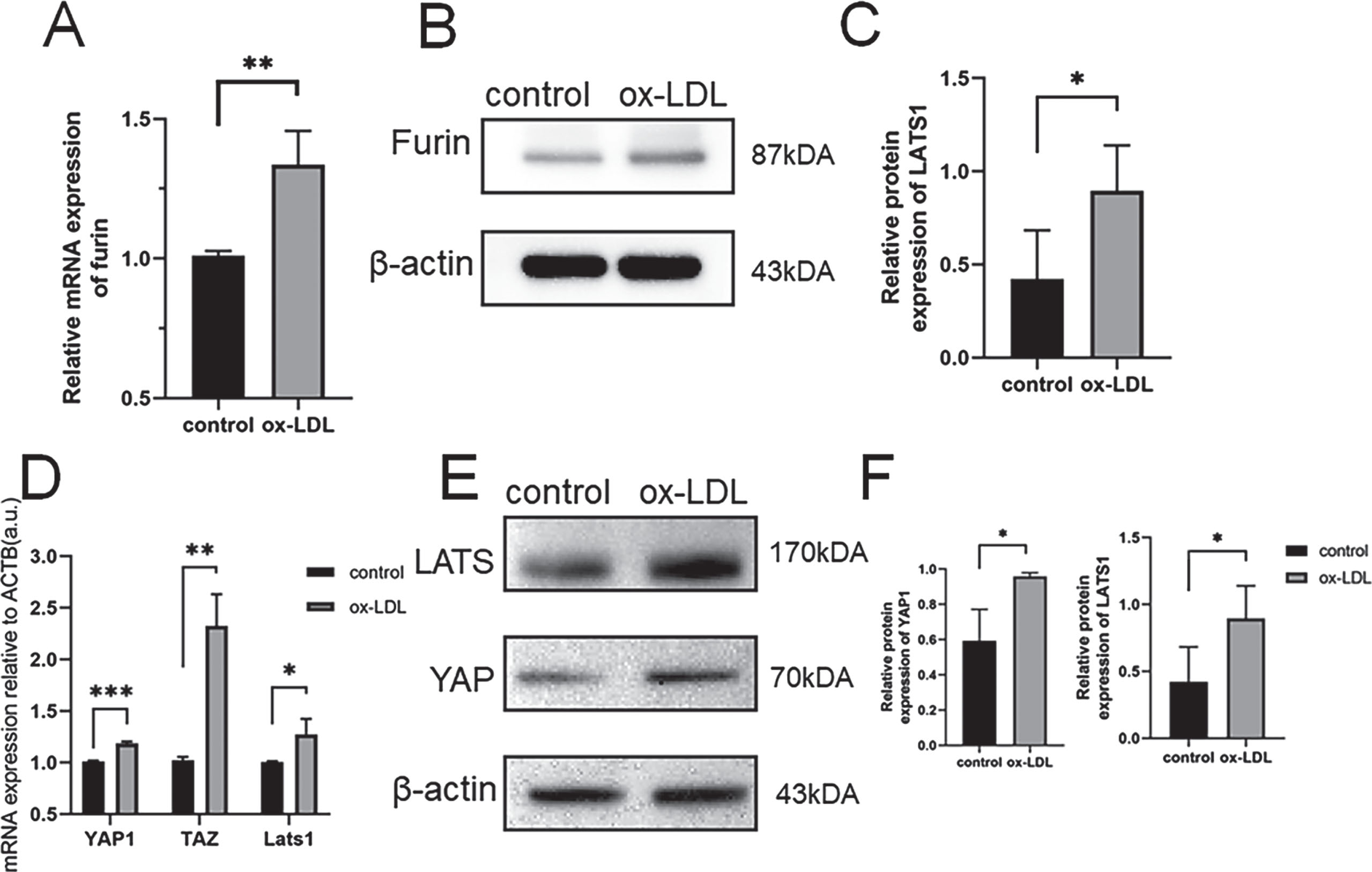
Quantification of furin mRNA levelsinducedbyox-LDL(A).
The western blot analysis results revealed that the Up-regulation of furin expression induced by ox-LDL. (BandC). Quantification of YAP1/LATS1mRNA levels induced by ox-LDL(D). The western blot analysis results revealed that the Up-regulation of YAP1/LATS1expressioninduced by ox-LDL. (EandF). Three asterisks (***) denote P value < 0.0.
The results of the growth curve (Fig. 3a), wound healing (Fig. 3b), and transwell (Fig. 3c) assays also indicated that the knockdown furin attenuates endothelial dysfunction caused by ox-LDL.
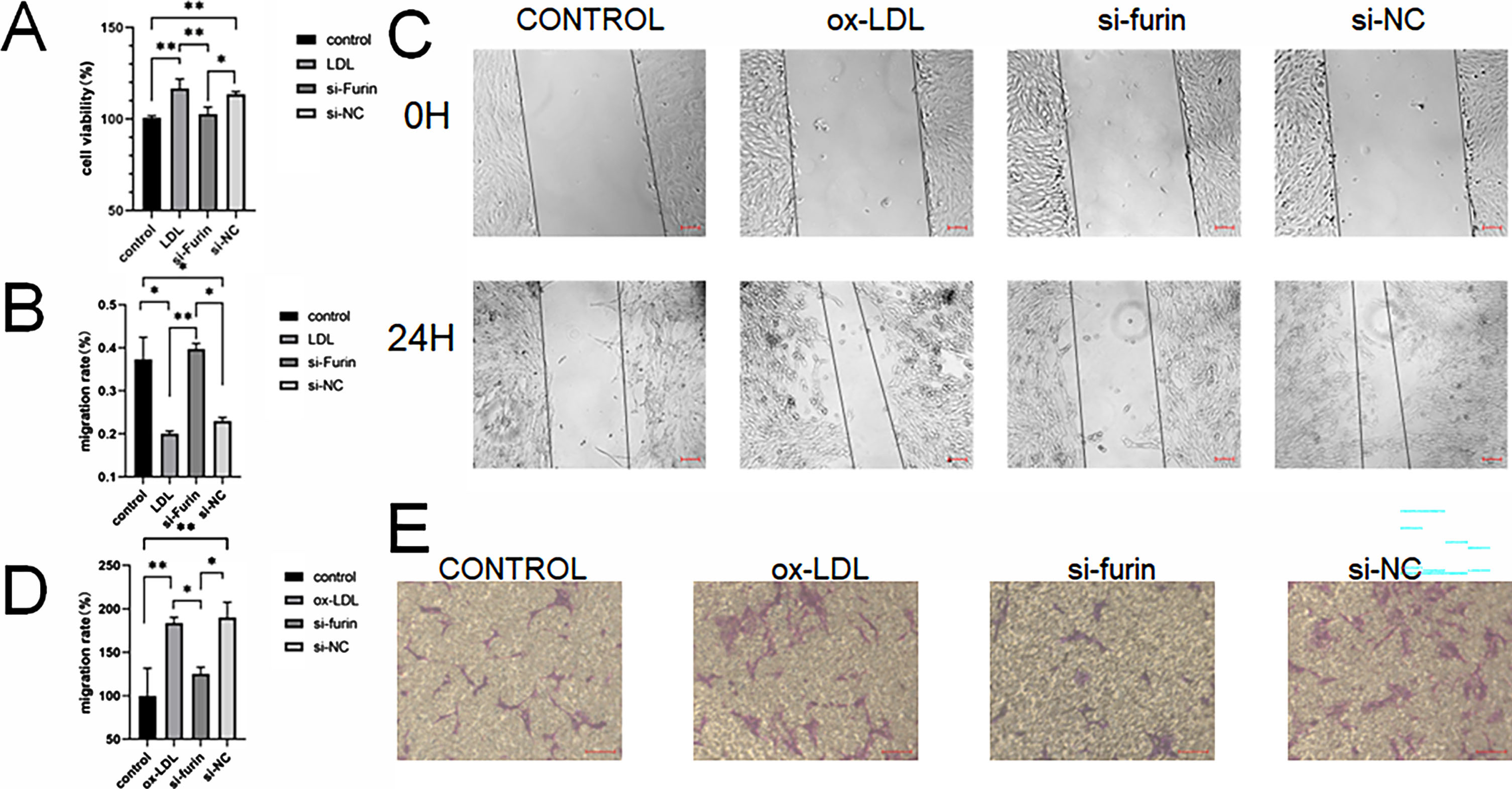
Ox-LDL promoted the proliferation of CMVECs, as shown by CCK8 (A), The migration abilities of CMVECs were assessed by wound healing (Band C scale bar 100 μm) and transwell (D and Escale bar 100 μm) assays. Two asterisks (**) denote P value < 0.01, and one asterisk (*) denotes P value < 0.05 by one-way ANOVA analysis (n = 3, >1,000 cells per trial).
We treated CMVECs with 100 μg/mL doses of ox-LDL for 24 h with transfection of furin inhibitor or negative control. Immunofluorescence staining showed that incubation of CMVECs with ox-LDL decreased expression of the endothelial marker VE-cadherin and increased expression of the mesenchymal marker α-SMA, and after transfection furin inhibitor, the progression of EndMT in CMVECs was slow down (Fig. 4A and 4B). Consistently, western blot showed that. (Fig. 4C and 4D). We found that the furin mRNA and protein expression was significantly increased in the negative control group as compared with the control group (Fig. 4E, 4F and 4G). furin protein and mRNA expression of siRNA group was significantly decreased compared with negative control.
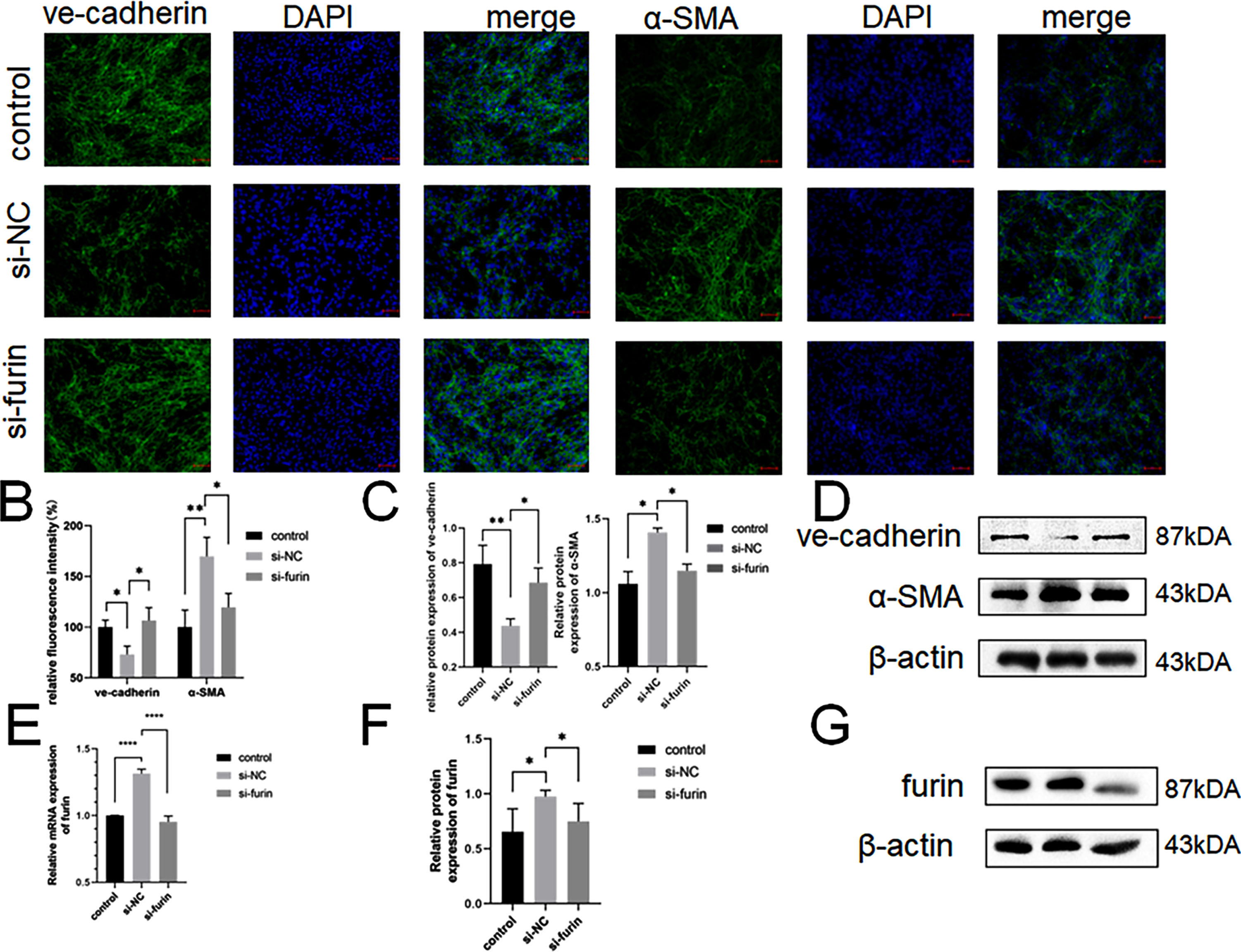
CMVECs were treated with normal medium, ox-LDL (100 μg/mL) with si-furin or si-NCfor 24 h. Immunofluorescence analysis of the protein expression of ve-cadherin and α-SMA.
Scale bar: 100 μm. (A) Quantification of Immunofluorescence analysis of ve-cadherin and α-SMA(B). The western blot analysis results revealed that the expression of EndMT molecules induced by ox-LDL. (C). Western blots indicating total ve-cadherin, α-SMA and β-Actin levels in whole cell lysates(D). Quantification of furin mRNA levels(E). The western blot analysis results revealed that the Up-regulation of furin. (F and G). Four asterisks (****) denote P value < 0.0001. Two asterisks (**) denote P value < 0.01, and one asterisk (*) denotes P value < 0.05 by one-way ANOVA analysis (n = 3, >1,000 cells per trial).
Venny analysis with Encori, TargetScan and miRDB was performed to screen the critical genes regulating endothelial cell function. Then, ten miRNAs were singled out (Fig. 5A). Hence, qRT-PCR were conducted to further detect the four miRNAs expression. The results indicated that the level of miRNA-486a-5p and miRNA-6413 in CMVEC cells was lower in Control group (Fig. 5B).
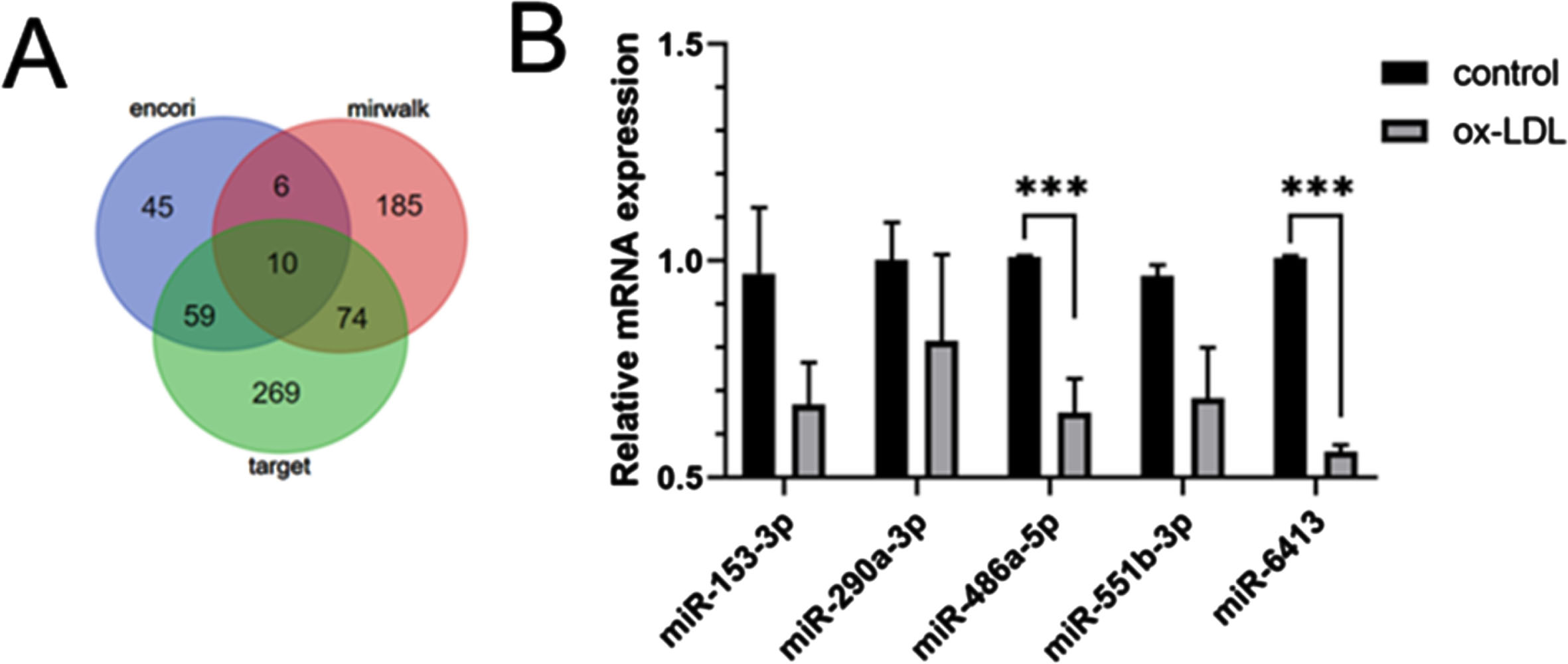
Venny analysis with Encori, TargetScan and miRDB was performed to screen the critical genes regulating furin (A). The level of the 5relativemiRNAs was evaluated by qRT-PCR analysis (B).
Endothelial dysfunction is often found in arterial vascular disease-prone areas, resulting in a series of lesions that are among the earliest detectable changes in the atherosclerotic process [1, 2]. As a key component of endothelial dysfunction, EndMT is a complex process of cell differentiation that involves the separation and migration of endothelial cells from the endothelium, with varying degrees of reduction of endothelial properties and acquisition of mesenchymal characteristics [12]. Endothelial cells transdifferentiate to form mesenchymal cells, which are myofibroblasts, smooth muscle cells, and osteoblasts, respectively. Accumulation of mesenchymal cells is central to plate formation and atherosclerosis [25]. And ox-LDL acts as an essential factor in the atherosclerotic process [26]. Therefore, in the present study, we stimulated the endothelial cells with ox-LDL and found that VE-cadherin and α-SMA coexisted in stimulated endothelial cells and that, after stimulation with ox-LDL, VE-cadherin expression decreased and α-SMA expression increased, confirming the presence of EndMT in the atherosclerotic process, which is also validated with previous studies [27].
Furin (PCSK3), on the other hand, is a member of the family of proprotein convertases, which are characterized by the conversion of a protein precursor into biologically active proteins and peptides through protein hydrolysis, thus allowing them to exert their biological effects [14]. Furin is known to activate TGF-β by cleaving the precursor of TGF-β1, and TGF-β is able to regulate EndMT [28, 29]. Therefore, we hypothesized that furin may be involved in the progression of atherosclerosis by regulating EndMT through cleavage of TGF-β. In our study, we also found that furin expression was elevated in the ox-LDL-induced endothelial cell injury model. In contrast, other studies have also found through genetic analysis that furin may play a role in vascular remodeling, inflammatory cell infiltration, and regulation of plaque stability, thereby affecting the early and late atherosclerotic process; studies at the cellular level have shown that the level of furin in endothelial cells affects the adhesion and migration of endothelial cells, whereas the progression of atherosclerosis in mice can be inhibited through the use of furin inhibitors [30]. Whereas in vivo and in vitro assays demonstrated that furin expression is directly related to the atherosclerotic process [31]. The formation of atherosclerotic plaques is characterized by the accumulation of mesenchymal stromal cells [32]. In this article, it was also found that cells showed abnormal proliferation and migration after ox-LDL stimulation, and that knockdown of furin was able to inhibit this abnormal proliferation and migration. This is verified by some experimental results [33, 34]. However, other studies showed that ox-LDL blocked the proliferation of HUVECs and induced apoptosis [10, 35–38]. The differences are subject to further research.
No previous study has confirmed whether furin can influence the progression of atherosclerosis by participating in the process of EndMT. In this paper, by knocking down furin, we found that VE-cadherin was up-regulated and α-SMA was down-regulated in cells with EndMT, which means that furin can participate in the process of EndMT and affect endothelial dysfunction, and the above facts prove that furin plays an important role in endothelial dysfunction and the formation of EndMT. ox-LDL stimulation in endothelial cells leads to aberrant proliferation and migration, and knockdown of furin reverses the proliferation and migration caused by ox-LDL.
The Hippo-YAP signaling pathway interacts with TGF-β downstream and thus may be involved in EndMT formation as a downstream of furin. Existing studies have demonstrated that disturbed blood flow enhances YAP activity, which triggers endothelial activation and atherosclerosis, whereas targeted inhibition of YAP attenuates endothelial inflammation and atherosclerosis [39]. Inhibitors of YAP also inhibit endothelial activation as well as potential mechanisms of atherosclerosis by suppressing YAP activity [40]. In the case of the hippo-/YAP signaling pathway that may be involved in furin’s downstream involvement in EndMT formation, our study also found that the Hippo/YAP signaling pathway was activated in a model of endothelial cell injury caused by ox-LDL, and the expression of two proteins, YAP1 and Lats1, in endothelial cells, was markedly upregulated after ox-LDL stimulation of endothelial cells. Moreover, the expression of YAP1 was down-regulated after the knockdown of furin. We speculate that this may imply that furin can affect endothelial dysfunction and EndMT by regulating the Hippo-YAP signaling pathway.
Finally, to explore whether there are microRNAs that can regulate furin in EndMT, we found several microRNAs that may be able to regulate furin through bioinformatics analysis, and experiments proved that the expression of these microRNA was down-regulated in the endothelial cell model that we established with ox-LDL stimulation, and among them, the one that was down-regulated most significantly was miRNA-486a-5p, which is most likely to regulate furin from influencing EndMT. miRNA-486a-5p is most likely to regulate furin from affecting EndMT, but the exact regulatory process needs to be further explored.
In summary, we found that furin participates in EndMT by regulating the Hippo/YAP signaling pathway, and concluded that the furin-Hippo/YAP axis affects endothelial dysfunction and participates in atherosclerosis, and briefly explored the upstream of furin in microRNA. However, how microRNA regulates the furin process needs to be further verified.
Footnotes
Acknowledgments
None.
Author contributions
Rui Zeng conceived and designed the experiments, performed the experiments, analyzed the data, prepared figures and/or tables, and approved the final draft. Yimin Wang performed the experiments, prepared figures and/or tables, and approved the final draft. Jun Chen conceived and designed the experiments, performed the experiments, authored or reviewed drafts of the article, and approved the final draft. Qiang Liu conceived and designed the experiments, analyzed the data, authored or reviewed drafts of the article, and approved the final draft.
Conflict of interest
All other authors have no conflict of interest.
Ethical approval
Not applicable.
Funding
This work was supported by the Natural Science Foundation of Zhejiang Province (No. LY22H270007), and the clinical basic application project of Zhejiang Rehabilitation Medical Center (ZKJC2202). The funder had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Data availability
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.