
Abstract
Alzheimer’s disease (AD) is a common neurodegenerative disorder that mainly affects the aged population and is characterized by the progressive loss of the hippocampal and cortical neurons, which results in memory and cognitive impairments. Trigonella foenum-graecum (fenugreek) has been reported to have hypoglycemic, hypocholesterolemic, hyperinsulinemic and anti-diabetic properties. Traditionally, it was used as a galactagogue and to treat anorexia, fever gastritis, gastric ulcers, and various nervous disorders. However, the neuroprotective effect of fenugreek seed powder against aluminum chloride (AlCl3) induced AD rats has not been analyzed. The result of the present study indicated that the chronic administration of AlCl3 induced significant learning and memory impairments, oxidative stress, and alterations in the protein immunocontent patterns of IDE and CDK5 (enzymes involved in the metabolism of tau and amyloid proteins), pTau, GFAP and Iba-1, IL-1β, IL-6, TNF-α, iNOS, NF-κB, COX-2, CDK5, BDNF, and STAT3. Our behavioral, biochemical, and molecular studies revealed that the co-administration of fenugreek seed powder significantly attenuated the AlCl3 induced memory deficits, amyloid and tau pathology, oxidative stress, and inflammation in AD rats could be due to the synergistic action of its active components.
INTRODUCTION
Alzheimer’s disease (AD) is the most common form of dementia in the elderly population and is characterized by progressive memory loss and other cognitive impairments. According to the National Institute of Health, it has affected about 18 million people worldwide [1] and is predicted to increase to about 65.7 million by 2030 and 115.4 million by 2050 [2]. The disease is mainly characterized by two major pathological hallmarks including the deposition of amyloid-β (Aβ) plaques and neurofibrillary tangles with a dramatic loss of neurons and synaptic dysfunction [3]. Aluminum (Al), an environmental toxin, is considered as a potential etiological factor that causes neuropathological, neurochemical and neurobehavioral changes which are similar to AD [4–6]. It is the third most abundant element in the earth, and due to the ubiquitous presence in food, water, dust, and air, humans are heavily exposed. Additional sources include aluminum utensils and use of aluminum foil [7], utilization of its compounds in different processes including papermaking, water treatment, fire retardants, fillers, food additives, colors and pharmaceuticals [8]. It also enters into the human body by the consumption of various food products such as corn, shellfish, yellow cheese, dairy products, spices, salt, breads, pastries, cakes, glace fruits, sausages, sugar-rich foods, baking mixes, tea, herbs and cosmetics. Al compounds are also used in antacids, phosphate binders, buffered aspirins, vaccines, and allergen injections. People living near cement factories are more prone to Al exposure as dispersed particles contain high amounts of Al [8]. Since Al is an environmental neurotoxin, exposure is increased due to lifestyle changes and implicated in the pathogenesis of AD. On this basis, the current experiment was designed.
Al is known to increase brain Aβ burden in experimental animals by influencing anabolism and catabolism of Aβ [9]. Aβ fibrils induced tau phosphorylation in in vivo and in vitro models of AD [10]. The phosphorylation state of tau protein mainly depends upon a balance between kinase-mediated phosphorylation and phosphatases intervened through dephosphorylation [11]. Cyclin-dependent kinases 2 and 5 (CDK 2 and CDK 5), glycogen synthase kinase-3β (GSK-3β), mitogen-activated protein kinase (MAPK), extracellular signal-regulated protein kinase 1/2 (ERK1/2), c-Jun N-terminal kinase (JNK), Akt, protein kinase A (PKA), and calcium-calmodulin protein kinase 2 (CaMKII) are a few kinases that are involved in the phosphorylation process, whereas some phosphatases such as protein phosphatase 1 (PP1), PP2A, PP2B, and PP2C participate in the dephosphorylation process of tau proteins [12]. Moreover, tau phosphorylation-dephosphorylation processes are regulated by a brain-derived neurotrophic factor (BDNF), a ubiquitous neurotrophin vital for the differentiation of neurons, synaptogenesis and synaptic plasticity. BDNF induces the dephosphorylation of tau protein and the cumulative lack of its signaling during AD promotes hyperphosphorylation [13]. Aβ and tau deposits in the AD brain probably cause alterations in cellular signaling pathways that trigger generation of reactive species, excessive inflammatory responses, a sustained rise in intracellular Ca2+, activation of caspase and nuclear factor-kappa B (NF-κB) and finally, apoptosis [14–16].
Fenugreek (Trigonella foenum graecum) is an old traditional medicinal legume plant, found in India, North Africa, the Mediterranean region, and Yemen [17]. The pharmacologically active components of fenugreek seeds are polyphenolics, flavonoids, steroid saponins, and polysaccharides. Galactomannans and 4-hydroxyisoleucine are known to have anti-diabetic, anti-inflammatory, antineoplastic, antipyretic, anti-ulcerogenic, anti-tumor, and immunomodulatory properties [18]. An in vitro study by Satheeshkumar et al. [19] demonstrated the acetylcholinesterase (AChE) inhibitory potential of standardized extract of fenugreek seeds. Fenugreek seed powder (FSP) ameliorated AlCl3 induced hepatorenal [20] and neurotoxicity [21] by virtue of its antioxidant and anti-inflammatory properties. The role of FSP in AD, and especially against AlCl3-induced oxidative stress, inflammation, and tau pathology, has not been so far studied. To fill the lacuna, the current study was designed.
MATERIALS AND METHODS
Animals
All the experiments were approved by the Institutional Animal Ethics Committee (Reg. No. 160/1999/CPCSEA, Proposal No. 1125) according to the National Guidelines on the Proper Care and Use of Animals in Laboratory Research, Indian National Science Academy, New Delhi (2000). Male Albino Wistar rats (200–225 g; 10–12 weeks) were procured and housed in standard conditions with food and water ad libitum at the Central Animal House, Rajah Muthiah Medical College & Hospital, Annamalai University.
Chemicals
AlCl3, thiobarbituric acid (TBA), reduced glutathione, 5,5-dithiobis-(2-nitrobenzoic acid), Anti-insulin degrading enzyme (IDE) (SAB2500529) and horseradish peroxidase (HRP) conjugated goat anti-rabbit IgG were purchased from Sigma Aldrich, Bangalore, India. Anti-glial fibrillary acidic protein (GFAP) (#12389), anti-NF-kB (#8242),anti-cyclooxygenase-2 (COX-2) (#12282), anti-CDK5 (#14145), anti-Signal transducer and activator of transcription 3 (STAT3) (#12640), anti-total-Tau (t-Tau) (#4019), anti-phospho-Tau (p-Tau) (#20194), and anti-β-actin (#3700) were purchased from Cell Signaling technology, USA. Anti-interleukin (IL)-1β (sc-12742), anti-IL-6 (sc-57315), anti-tumor necrosis factor alpha (TNF-α) (sc-133193), anti-inducible nitric oxide synthase (iNOS) (sc-7271), anti-BDNF (sc-65514), and anti-ionized calcium binding adaptor molecule 1 (Iba-1) (sc-32725) were purchased from Santa Cruz Biotechnology, USA. All other chemicals used were of analytical grade.
Preparation of FSP
Trigonella seeds were purchased from the local market, Chidambaram, Tamilnadu, India, germinated and finely powdered. For the preparation of 5% of FSP, about 5 g of dried FSP were mixed with 95.0 g of ground rat food [21].
Experimental design and dosage fixation
In our lab, a pilot study was conducted with three different doses of FSP (2.5, 5, and 10%) to determine the dose-dependent effect of FSP in AlCl3-induced Alzheimer rats [22]. After scarification, the hippocampus and cortex were excised and utilized for the determination of Al and AChE. Rats induced with AlCl3 showed a significant decrease in body weight when compared with control rats. Oral treatment with FSP to AlCl3-induced rats significantly increased the body weight dose dependently. There are no significant changes in weight gain of FSP-only treated rats when compared with control rats. The water intake (mL/d) and food intake (g/d) of rats in the control and experimental groups also showed no significant differences. We found observed that 5 and 10% of FSP administration showed a significant reduction in the levels and activities of Al and AChE compared to a 2.5% FSP treatment. No significant changes were found in the levels and activities of Al and AChE between 5 and 10% FSP co-treated rats. Hence, 5% FSP is considered as the effective dose. As AChE inhibitors are the major class of drugs approved for AD in practice, affording symptomatic relief and improvement in cognitive function [23], its activity measurement is considered as a marker for the selection of the FSP dose. Five percent FSP treatment significantly attenuated the AlCl3 induced amyloid burden by diminishing the protein immunocontents of APP, Aβ1 - 42, β-, and γ-secretases [22].
Forty-eight rats were divided randomly into four (n = 12) groups: Group I animals received saline and were considered to be the control. Group II rats were administered with AlCl3 (100 mg/kg b.w., oral) for 6 weeks [24]. Group III rats received AlCl3 as group II and 5% FSP mixed in grounded standard rat feed (i.e., 5 g of dry ground FSP in 95 g of ground rat food) [22]. Group IV animals received 5% FSP mixed in ground standard rat feed for 6 weeks. Six animals in each group were used for the analysis of the elevated plus maze test and the estimation of thiobarbituric acid reactive substances (TBARS) and antioxidants. Another six animals were used for the analysis of the radial arm maze test and protein content studies by western blotting.
Modified elevated plus maze (EPM) test
The Modified EPM test is mainly utilized for the evaluation of spatial learning and memory [25]. The maze consisted of two opposite closed arms (50 cm×10 cm) with 40 cm high walls open arms and two open arms of similar size without walls. The arms were connected with a central square (10 cm×10 cm) and placed 50 cm above the ground. Animals were tested for acquisition memory at end of the experimental period (20th day). Each rat was placed individually at one end of the open arm. The time taken by the animal to move from the open to the closed arms in the maze was measured as the initial transfer latency (ITL). Animals were permitted to explore for 20 seconds after recording the ITL and were then returned to the home cages. The same experiment was repeated on the 42nd day by placing the rat in an open arm as the retention transfer latency (RTL) was assessed.
Radial arm maze test
The apparatus contained an octagonal center compartment connected with 8 equidistantly spaced arms (15×15×70 cm). 4 arms were baited with sweetened cereals in grooves, which were positioned 2 cm away from the arms end. During the training period, rats were allowed to explore till the end of the radiating arms. The baits were steadily limited to grooves. During the trial, timings began after the rat was placed in the central area of the maze and allowed to move freely. Arm preferences were recorded if the animal entered at least half the length of the arm. When the rat chose all baited arms, or spent 10 min, then it was considered that the trial was completed. The following parameters were calculated: working and reference memories measured by counting the number of repeated entries to baited arms and the number of entries to unbaited arms, respectively. The mean time taken to complete the test in all trials was also measured [26]. After performing the behavioral experiments, the rats were sacrificed to procure the hippocampus and cortex for the biochemical analysis and protein immunocontent studies.
Tissue preparation for biochemical estimations
Dissected tissues were homogenized in a mortar with a 10 mM Tris-HCl buffer (pH 7.0) containing a 10μl/ml protease inhibitor to get 5% (w/v) homogenate and centrifuged (800 xg for 5 min at 4°C) to obtain the nuclear fractions. The supernatant 1 (S1) was used for the estimation of TBARS content. The remaining pellet was further centrifuged (10500 xg for 30 min at 4°C) and the obtained post-mitochondrial supernatant was utilized for the estimation of antioxidants such as reduced glutathione (GSH), superoxide dismutase (SOD), catalase, and glutathione peroxidase (GPx).
Estimation of TBARS
To the brain tissue extracts, 0.2 ml phenyl methosulphate was added and kept in water bath shaker (37°C). After 1 h, 0.4 ml of 0.67% TBA and 0.4 ml of 5% trichloroacetic acid were mixed and the mixture was centrifuged at 3000 xg for 15 min. The collected supernatant was incubated in a hot waterbath for 10 min. Changes in optical density were measured spectrophotometrically at 532 nm after cooling [27].
Estimation of GSH
The level of GSH in the hippocampus and cortex was measured by the method followed by Jollow et al. [28]. The homogenate was centrifuged at 16000 xg for 15 min at 4°C and 0.5 ml of supernatant was added to a 0.1 mM solution of 5,5-dithiobis [2-nitrobenzoic acid] in a 1 M phosphate buffer (pH 8). The changes in optical density were measured at 412 nm.
Assay of SOD
Briefly, the hippocampal and cortical extracts were added to 960μl of sodium carbonate buffer (pH 10.2) containing xanthine, nitro blue tetrazolium, and ethylene-diamine-tetra acetic acid to which 20μl of xanthine oxidase was added. Changes in absorbance were measured at 560 nm [29].
Assay of catalase activity
To 20μl of the brain tissue supernatant, 50μl of 1 M Tris-HCl buffer (pH 8.0) containing 5 mM EDTA, 900μl of 10 mM H2O2, and 30μl of MQ water was added. The activity of catalase was observed at 240 nm and expressed as rate of the decomposition of hydrogen peroxide [30].
Assay of GPx
The assay mixture consisted of 20μl of the brain extract, 100μl of 1 M Tris-HCl (pH 8.0) with EDTA, 100μl of glutathione reductase (10 U/ml), 20μl of 0.1 M GSH, 10μl of 7 mM hydroperoxide, 100μl of 2 mM NADPH, and 650μl distilled water. Oxidation of NADPH was determined at 340 nm [31].
Western blot analysis
Brain tissues were homogenized in a cold suspension lysis buffer (20 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1 mM EDTA, 30μg/mL apoprotein, and 1 mM phenyl methyl sulfonyl fluoride) andthe homogenates were centrifuged (10,000 rpm/15 min/4°C). The protein content of the supernatant was assayed [32] and diluted to give an equal protein concentration of 50μg. Samples containing equal cellular protein were loaded on 10% SDS-polyacrylamide gel electrophoresis and resolved. The gel was transferred to a polyvinylidene difluoride membrane (Millipore). For the assessment of the immuno-content of specific protein, the membranes were incubated in GFAP, IL-1β, IL-6, TNF-α, iNOS, NF-kB, COX-2, CDK5, BDNF, STAT3, t-Tau, p-Tau, IDE, and β-actin IgG (monoclonal; 1:1000) in 5% BSA in Tris-buffered saline and 0.05% Tween-20 (TBST) with mild shaking overnight (4°C) [33]. Then the membranes were incubated for 2 h at room temperature with their corresponding HRP conjugated secondary antibodies and washed. Immunoreactive proteins were visualized by the chemiluminescence protocol (GenScript ECL kit, USA) and the densitometric analysis was performed with a gel image analysis program. Data were normalized by background subtraction and β-actin was used as an internal control.
Data analysis
All data were expressed as the mean±standard error (SEM) of the number of experiments. The statistical significance was calculated by one-way analysis of variance (ANOVA) using SPSS version 15.0 and the individual comparisons were obtained by Duncan’s Multiple Range Test (DMRT). A value of p < 0.05 was considered to indicate a significant difference between groups and the values sharing a common alphabet do not differ significantly with each other.
RESULTS
FSP rescue cognitive performance in Wistar rats
In the elevated plus maze task, mean ITL for each group showed no significant variation. AlCl3treated rats showed poor performance throughout the experiment and exhibited no significant changes in the mean retention transfer latencies (1st and 2nd) as compared to ITL, indicating memory impairment. Chronic co-administration of FSP to the AlCl3 treated rats showed significant reduction in the mean retention latencies on days 21 and 42 compared to AlCl3 alone treated group. There is no difference in the mean transfer latencies of FSP alone treated and control animals (Fig. 1).
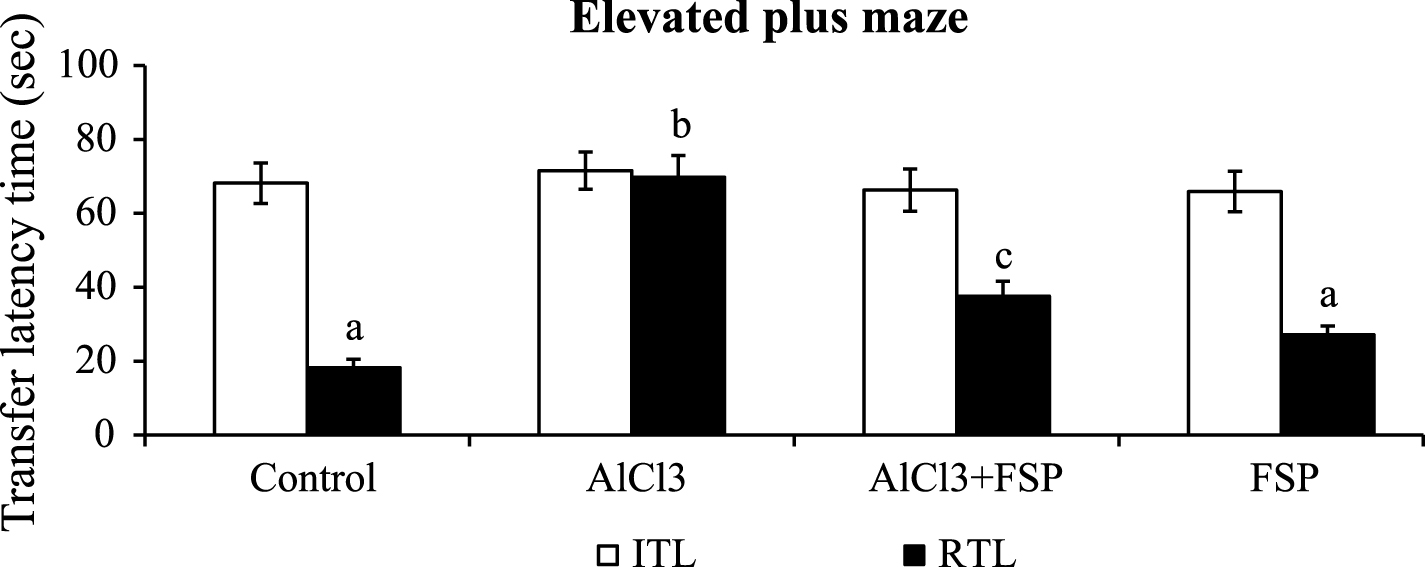
Effect of FSP on mean transfer latencies in modified elevated plus maze test. Both the initial and retention transfer latencies were assessed. Control group (n = 6); AlCl3 treated group (n = 6); AlCl3 + FSP group (n = 6); FSP group (n = 6). Data were presented as mean±standard error (S.E.). Values not sharing the same alphabet differ significantly: ap < 0.05 compared to AlCl3 and AlCl3 + FSP treated groups; bp < 0.05 compared to control; cp < 0.05 compared to AlCl3-treated group.
In the radial arm maze test, the animals administrated with AlCl3 showed more errors both in the reference and working memory task and took more time to finish the task when compared to control group. Co-administration of FSP to AlCl3-treated rats significantly (p < 0.05) attenuated the learning and memory impairment compared to AlCl3 alone treated animals. FSP alone treated rats showed no significant differences in behavioral patterns when compared to control animals (Fig. 2).
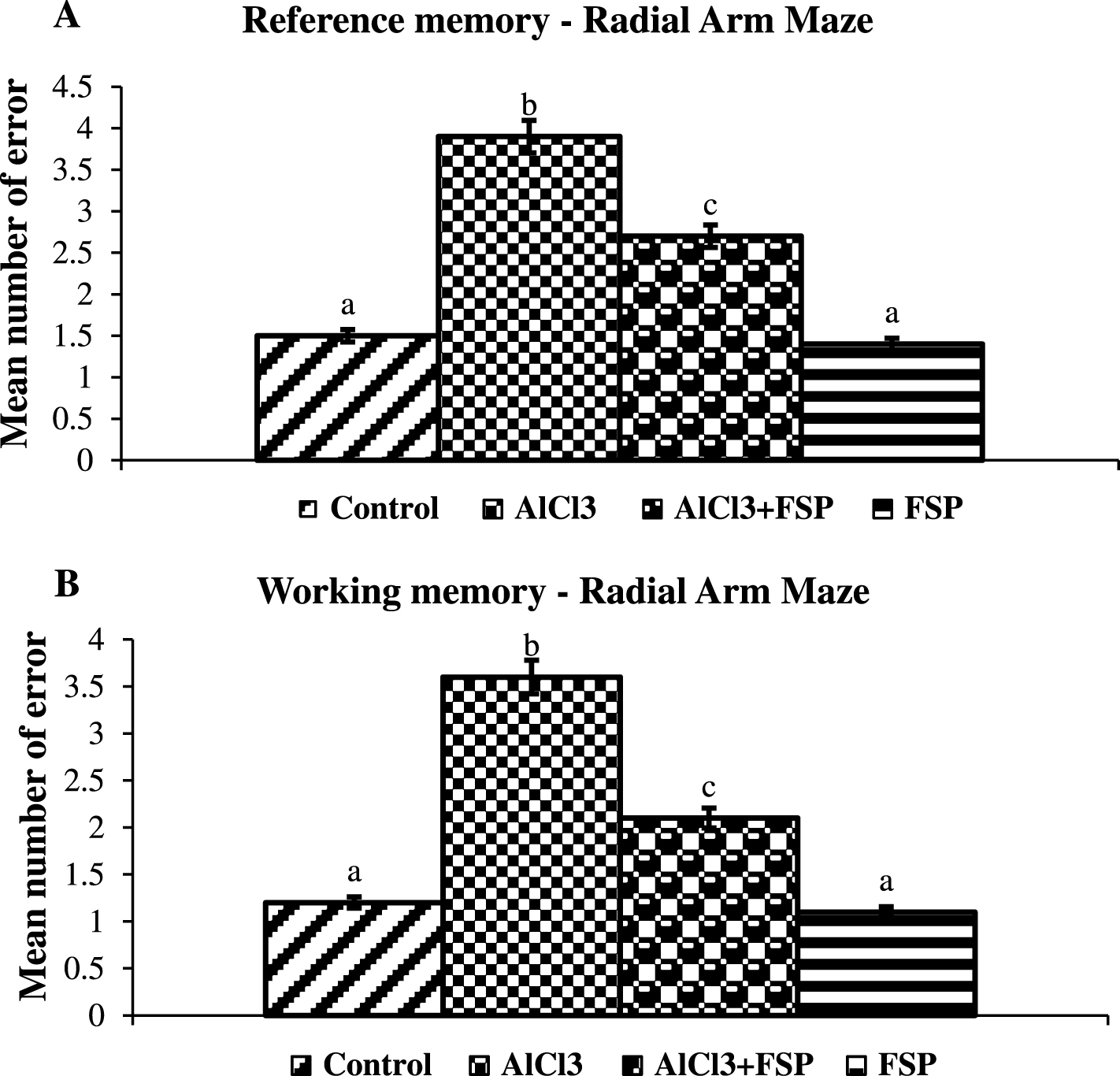
Effect of FSP on memory impairment in radial arm maze test. Both the working (A) and reference (B) memories were assessed. Control group (n = 6); AlCl3 treated group (n = 6); AlCl3 + FSP group (n = 6); FSP group (n = 6). Data were presented as mean±standard error (S.E.). Values not sharing the same alphabet differ significantly: ap < 0.05 compared to AlCl3 and AlCl3 + FSP treated groups; bp < 0.05 compared to control; cp < 0.05 compared to AlCl3treated group.
FSP nullifies oxidative stress
Figure 3A-E shows the oxidant and antioxidant imbalance in the hippocampus and cortex of control and experimental rats. In the AlCl3 treated rats, the levels of TBARS were significantly elevated and the levels of GSH and the activities of SOD, catalase and GPx were diminished significantly in the hippocampus and cortex compared to the control animals. Co-treatment of FSP significantly attenuated the oxidative stress by decreasing the levels of TBARS and enhancing the levels of GSH and the activities of SOD, catalase, and GPx in the above said brain regions as compared to AlCl3-treated animals.
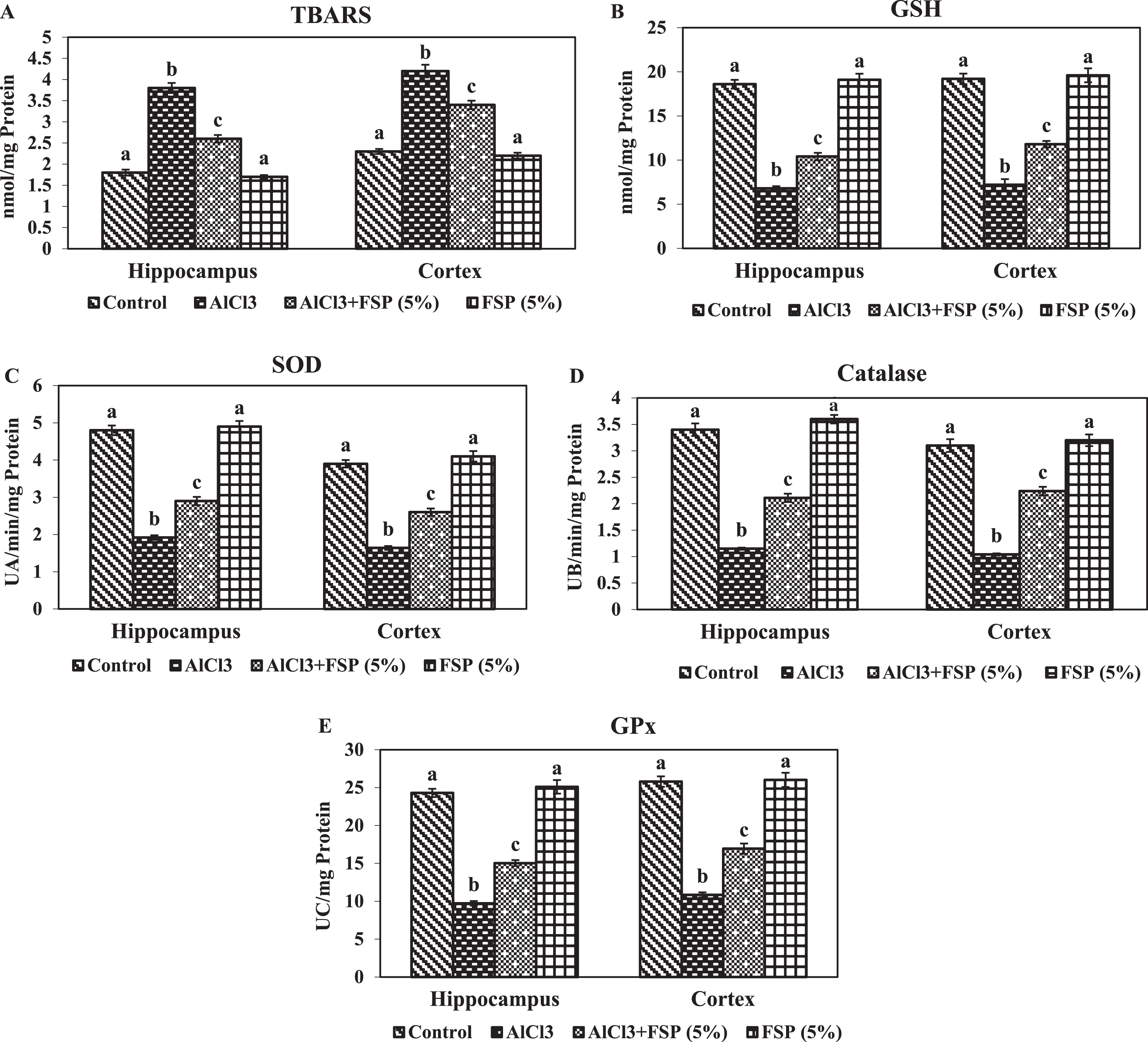
Effect of FSP on oxidative stress induced by aluminum chloride. Levels of the TBARS(A) and GSH (B) and the activities of SOD (C), catalase (D) and GPx (E) in hippocampus and cortex were assessed. Control group (n = 6); AlCl3 treated group (n = 6); AlCl3 + FSP group (n = 6); FSP group (n = 6). Data were presented as mean±standard error (S.E.). Values not sharing the same alphabet differ significantly: ap < 0.05 compared to AlCl3 and AlCl3 + FSP treated groups; bp < 0.05 compared to control; cp < 0.05 compared to AlCl3-treated group.
FSP attenuates tau pathology
Rats treated with AlCl3 exhibited significant increase in the protein immunocontents of IDE, CDK 5, and pTau in hippocampus and cortex, whereas their contents were significantly attenuated by co-treatment with FSP. FSP alone treatment did not exhibit significant changes in the expression of above said indices compared with the control group. In our study, FSP treatment effectively reversed the AlCl3-induced Aβ and tau pathology by altering the expressions of IDE and CDK 5 (Fig. 4A-E).
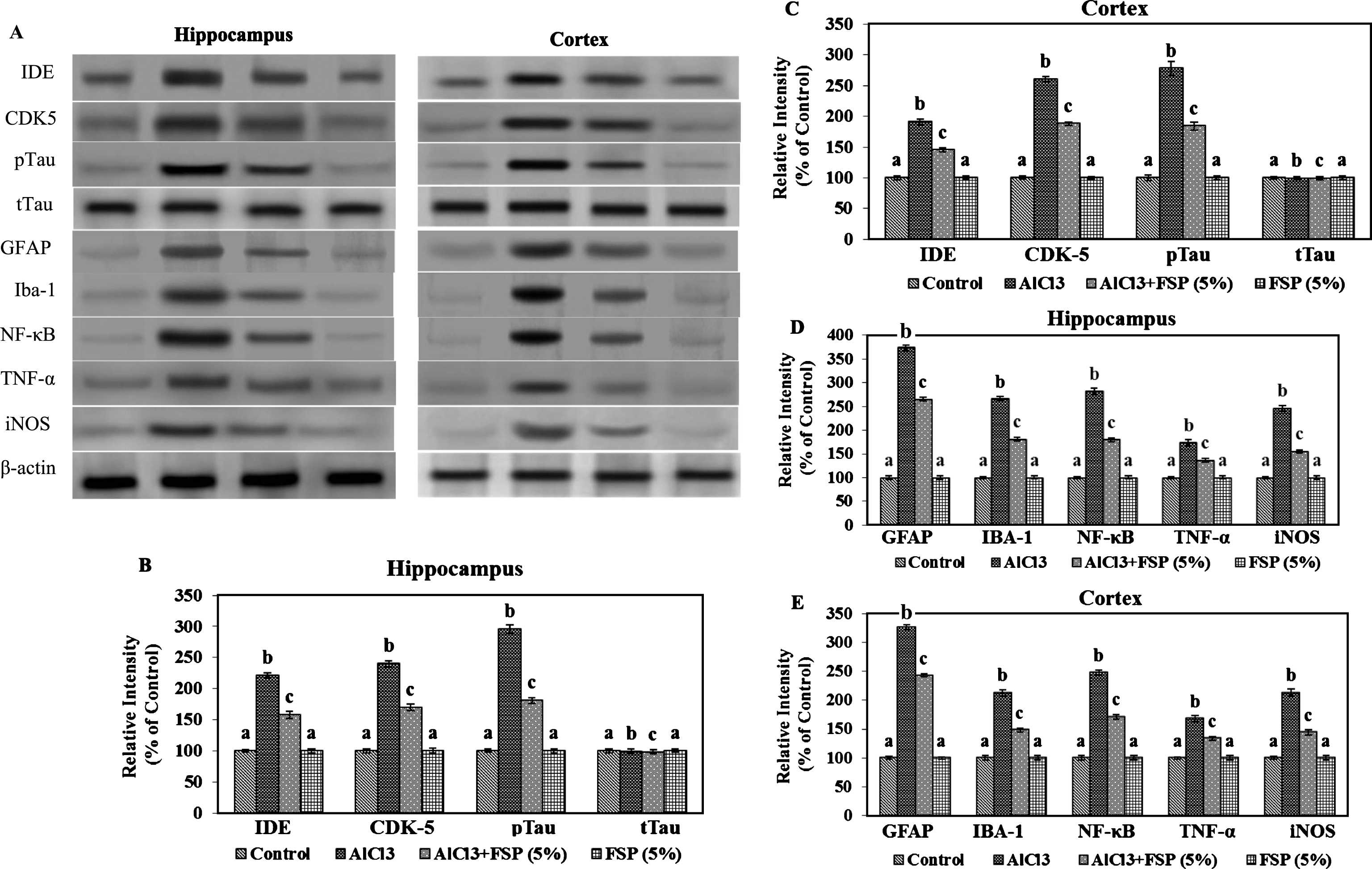
Effect of FSP on the expression (A) of IDE, CDK5, pTau, tTau, GFAP, Iba-1, NFκ-B, TNF-α, and iNOS in hippocampus (B,D) and cortex (C,E). Evaluation of protein expression by using western blot. Control group (n = 3); AlCl3 treated group (n = 3); AlCl3 + FSP group (n = 3); FSP group (n = 3). Data were presented as mean±standard error (S.E.). Values not sharing the same alphabet differ significantly: ap < 0.05 compared to AlCl3 and AlCl3 + FSP treated groups; bp < 0.05 compared to control; cp < 0.05 compared to AlCl3-treated group.
FSP ameliorates inflammation
To demonstrate the protective effect of FSP in AlCl3 induced gliosis, the protein immunocontents of GFAP and Iba-1 were studied in the hippocampus and cortex of control and experimental rats.The protein content of GFAP and Iba-1 were found to be enhanced in the AlCl3 treated rats, whereas the protein content was reduced by chronic FSP treatment. Activation of astrocytes and microglia leads to the production of pro-inflammatory cytokines and toxic compounds. Therefore, the protein contents of iNOS, NF-kB, TNF-α, IL-1β, IL-6, and COX-2 were performed by western blot technique. Western blot analysis indicated the enhanced protein contents of pro-inflammatory cytokines in AlCl3 treated rats, whereas FSP treatment attenuated these indices through its anti-inflammatory properties (Fig. 4A-E, Fig. 5A-C).
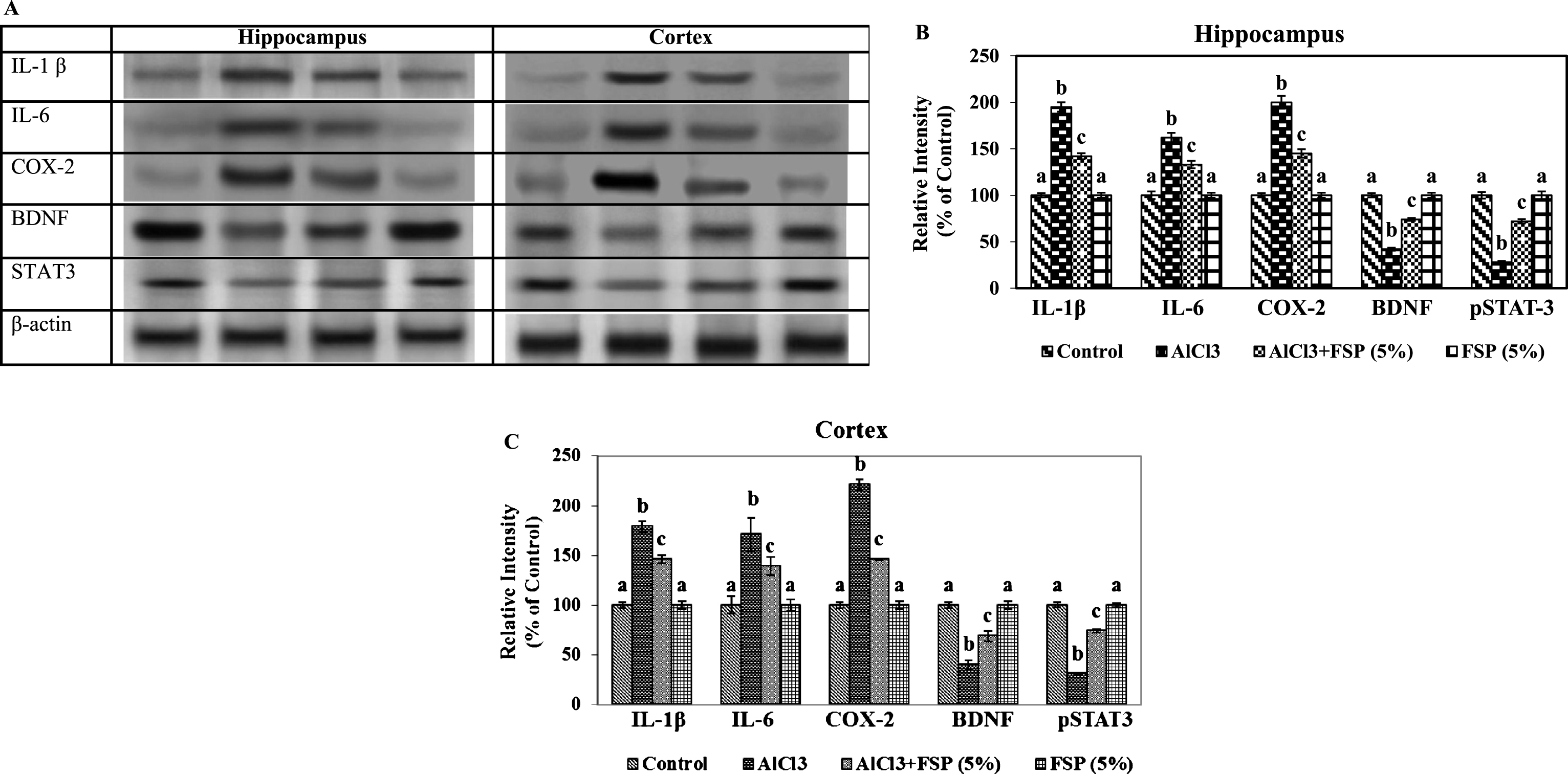
Effect of FSP on the expression (A) of IL-1β, IL-6, COX-2, BDNF and STAT 3 in hippocampus (B) and cortex (C). Evaluation of protein expression by using western blot. Control group (n = 3); AlCl3 treated group (n = 3); AlCl3 + FSP group (n = 3); FSP group (n = 3). Data were presented as mean±standard error (S.E.). Values not sharing the same alphabet differ significantly: ap < 0.05 compared to AlCl3 and AlCl3 + FSP treated groups; bp < 0.05 compared to control; cp < 0.05 compared to AlCl3treated group.
FSP modulates BDNF and STAT 3 expressions
We performed a western blot technique to investigate changes in the protein immunocontents of BDNF and STAT 3 in control and experimental groups. Animals treated with AlCl3 manifested significant reduction in the protein contents of BDNF and STAT 3 in hippocampus and cortex. Meanwhile, these alterations were significantly attenuated by co-treatment by FSP (Fig. 5A-C).
DISCUSSION
We found that FSP co-administration attenuated AlCl3 induced learning and memory impairments, Aβ accumulation, oxidative stress, tau hyperphosphorylation, and inflammation in the hippocampus and cortex of the rat. EPM is generally used to measure anxiety, whereas its modified form is observed to measure learning and memory impairments. The test is based on a conflict between the tendency of rodents to explore a novel environment and aversive behavior on open spaces and heights. If the animals faced a choice between walls enclosed arm versus unprotected open ones, they prefer to stay in the protected areas. The time required for animals to move from the open arm to either of the enclosed arms is called as transfer latency. Trained animals (repeated exposure of the animals to the open arms) showed a reduction in the transfer latency, as a result of learning acquisition and retention. The transfer latency is increased even after training in the memory impaired rats. Both the working and reference memories are assessed in rodents using radial arm maze paradigms [34]. Working memory is assessed by examining the entry of rats into each arm once and the reference memory is examined by counting entry into the arms containing food. If the rat re-enters the arm or fails to enter into the food-containing arm, then it is considered as a working and reference memory error. In the present study, AlCl3 treatment enhanced the retention transfer latency in the modified elevated plus maze test and declined the reference and working memories in the radial arm maze test that has been associated with the impairment in memory-related functions. Previous studies from our lab indicated that the reason for this disturbance in memory and learning of animals may be due to the influence of Al in the cholinergic system [5, 35]. In the central nervous system, acetylcholine (ACh) is a neurotransmitter that plays a vital role in motor and memory functions. AChE is the key enzyme, involved in the degradation of ACh. Acetylcholinesterase inhibitors increase the cholinergic transmission by inhibiting the loss of ACh and therefore used for nullifying the symptoms of patients with AD [36]. Satheeshkumar et al. [19] demonstrated the in vitro AChE inhibitory activities of crude fenugreek seed extracts, hydro alcoholic, ethyl acetate, chloroform and total alkaloid fractions, and trigonelline, which is its active component. Hence, it is possible that, the memory-enhancing activity of FSP might be partially through the inhibition of AChE.
AlCl3 is reported to induce the formation and accumulation of insoluble Aβ peptide and promotes the hyperphosphorylation of tau protein [37]. In the present study, AlCl3 administration enhanced the protein immunocontent of IDE, the rate-limiting Aβ-degrading enzyme, whereas FSP treatment attenuated the protein contents of IDE. The cause of increased IDE may be partly due to activated astrocytes as a result of Aβ-triggered neuroinflammation [38]. In several countries like Iran, Southern India, and Africa, fenugreek has been used as a traditional medicine or remedy for inflammation and its related effects for many years. The main chemical constituents responsible for the anti-inflammatory activity are alkaloids, saponins, and flavonoids [39]. Abnormal hyperphosphorylation of tau is dependent on both the enhanced activation of kinase, as well as the diminution of phosphatase activities [10]. Cyclin-dependent kinase 5 (CDK5) plays a major role in causing aberrant hyperphosphorylation. AlCl3 enhanced the protein immunocontent of CDK5 in the hippocampus and cortex and thereby favoring tau hyperphosphorylation, but this was significantly reduced after FSP co-treatment.
Accumulation of Al enhances reactive oxygen species (ROS) formation that depletes the normal antioxidant defense mechanism, thereby further enhancing oxidative stress and lipid peroxidation processes [40]. The interaction of redox transition metals, abnormal mitochondria and oxidative stress response elements, including inflammation, contributes to the generation of ROS in AD conditions [41]. Al causes progressive deterioration of mitochondrial functions due to excessive free radical generation. It is a non-redox active metal that is capable of increasing the oxidative stress by enhancing pro-oxidant properties of transition metals such as iron and copper [42]. This further damages other cellular molecules including DNA damage, nitration of protein residues, and lipid peroxidation [43]. TBARS is an in vivo biomarker that reflects the overall status of oxidative stress in brain and is an end product of peroxidation of polyunsaturated fatty acid. SOD, catalase, and GPx are the enzymatic antioxidants, which are capable of removing oxygen free radicals in the body. SOD represents the first line of defense against initially formed superoxide and dismutases it to less toxic H2O2. Catalase protects SOD by converting H2O2 into water and oxygen. Reduced glutathione is the most abundant intracellular antioxidant and is involved in direct scavenging of ROS or serving as a substrate for the GPx enzymes that catalyze the detoxification of H2O2. As oxidative stress and cognitive dysfunction are strongly correlated, the agent that modulates oxidative stress may be used as potential anti-dementia agent [44–46].
In our study, we demonstrated that FSP diminished the levels of TBARS, restored reduced glutathione, and increased SOD, catalase, and glutathione peroxidase activities in AlCl3 treated rats. Supplementation of fenugreek in the diet attenuated the AlCl3 [20], deltamethrin [47], and acrylamide [48] induced oxidative stress by lowering the lipid peroxidation processes and elevating the levels and activities of antioxidants in rats. Moreover, the treatment of FSP exhibited the antioxidant function against alloxan-induced neurotoxicity by enhancing the activities and protein immunocontents of SOD, catalase, and GPx in the brain of diabetic rats [49], which supports our findings.
Cognitive dysfunction is generally accompanied by a rise in pro-inflammatory cytokines and inflammation in the hippocampal and cortex regions [50]. Brain parenchyma comprises neurons and glial cells, in which the latter is classified into macroglia and microglia. Macroglia is further classified into oligodendrocytes, astrocytes, radial glia, and ependymal cells. Both the astrocytes and microglia are involved in the homeostatic functions like differentiation, development, and survival of neurons [51]. Reactive gliosis is a phenomenon during which both the astrocyte and microglia get activated in response to an array of toxins that contribute to the pathogenesis of neuro-inflammation and neurodegeneration [52]. Severe activation of astrocytes has been implicated in the pathogenesis of AD [53] and results in secretion of various neurotoxic substances and elevated the protein content of GFAP, which is considered as a marker protein for astrogliosis [54]. Microglia respond to any insult and injury faster than astrocytes [55]. Only microglial cells express Iba-1. Al plays a major role in the activation of inflammatory process. Our results showed that elevated protein content of GFAP and Iba-1 demonstrated both macro and microgliosis after AlCl3 treatment, whereas the FSP treatment attenuated the reactive gliosis in AlCl3–induced neurotoxicity by reducing the expressions of GFAP and Iba-1.
Although oxidative stress and inflammatory events are independent phenomena, they are inter-related. ROS can activate redox-sensitive transcription factors, such as NF-κB and p53 in glial cells, which are able to produce the pro-inflammatory cytokines, potentially neurotoxic reactive oxygen species and excitotoxins. Both microglia and astrocytes produce multiple pro-inflammatory factors, including cytokines (tumor necrosis factor-α (TNFα), interleukins (IL-1β and IL-6), chemokines, reactive oxygen species, and COX-2) [56, 57]. Experimental and clinical evidences have demonstrated the enhanced production of pro-inflammatory cytokines such as TNF-α, IFN-γ, IL-1β, IL-6, IL-18, and the upregulation of their related receptors in the AD brain [58, 59]. Chronic deposition of Al in the brain drives neuroinflammation by activating astrocytes and microglia, which are reported to be a major source of pro-inflammatory cytokines in AD [60]. The results of our present study demonstrated that AlCl3 administration induces the protein immune-contents of pro-inflammatory cytokine such as NF-κB, TNF-α, IL-1β, IL-6, and COX-2, which is consistent with a previous report [61].
BDNF signaling is also involved in learning and memory by binding to its main functional protein-tyrosine kinase-β (TrkB) receptor, present in the hippocampus, cortex, and basal forebrain. Reduced BDNF signaling leads to defective hippocampal and cortical synaptic plasticity and transgenic mice lack either BDNF or TrkB exhibited arborization impairment, defective synaptic sprouting, lowered synapse number, and impaired hippocampal long term potentiation (LTP). In our current study, diminished memory impairment in AlCl3 treated rats may be partially due to the lowered protein content of BDNF. BDNF promotes neuronal survival through activation of tyrosine receptor kinase B (TrkB) by JAK2/STAT 3 signaling [62]. STAT 3 may serve as a signal transducer for Trk receptors in neuronal cells and that BDNF-induced activation of STAT 3 may play crucial roles in the downstream functions of neurotrophin signaling. Phosphorylated STAT 3 dimerize and translocates into the nucleus where it induces the transcription of target genes critical for promoting neuronal survival such as Bcl-2 [63, 64]. FSP treatment induced upregulation of phosphorylated STAT 3, a key component of the cellular anti-apoptotic cascade, in the hippocampus and cortex of Al-treated rats, thereby enhancing neuronal survival. Oxidative/-nitrosative stress further diminishes the BDNF in the brain [65]. Enhanced protein immune content of BDNF and STAT 3 in FSP co-administered rats indicated that FSP shielded the protein contents of BDNF, which might be due to its antioxidative potential.
Conclusion
To conclude, FSP could be used as a possible therapeutic agent in the treatment of cognitive dysfunction-associated disorders such as AD. Our finding suggested that the neuroprotective role of FSP in the brain is multifaceted, which may be due to the synergistic action of some active constituents. However, further analysis of the neuroprotective ability of active components of fenugreek is warranted.