
Abstract
Keywords
INTRODUCTION
The decline in cognitive function associated with Alzheimer’s disease (AD) is mainly induced by neuronal apoptosis resulting from the increase in amyloid-β peptide (Aβ) production. Evidence suggests that neuronal loss is one of the core events underlying both impaired cognitive and neuropsychiatric symptoms. The hippocampus, a key area for learning and memory, is considered a special target for neuronal apoptosis associated with many risk factors, such as cerebrovascular diseases [1], traumatic brain injury [2], and hyperlipidemia [3]. The brain, the second organ after adipose tissue with the highest lipid content in the body [4], is the most vulnerable to uncontrolled peripheral hyperlipidemia. It was reported that hyperlipidemia results in decreased general cognitive performance, memory impairment, and atrophy [4]. Persistent hyperlipidemia promotes the production of oxidized low-density lipoprotein (oxLDL) that plays an important role in lipid metabolism disorders [5], but it is not clear how abnormal lipid metabolism is involved in neurodegenerative diseases.
Proprotein convertase subtilisin/kexin type 9 (PCSK9) is a member of the subtilisin gene family of proprotein convertases, which is mainly involved in the degradation of LDL receptor (LDLR) in the liver [6]. With apolipoprotein B-100 and apolipoprotein E (apoE) as the ligands, LDLR plays a major role in the regulation of plasma cholesterol and intracellular levels. The gain of function mutations of PCSK9 are responsible for autosomal dominant hypercholesterolemia, whereas loss of function mutations of PCSK9 are associated with a reduction of LDL-cholesterol and the risk of coronary heart disease [7]. Although Shibata et al. [8] and Reynolds et al. [9] did not find direct evidence to suggest that PCSK9 and its two mutations are associated with AD patients, their results did not rule out the positive correlation between PCSK9 and AD. In our previous experiments, we discovered that a high-fat diet increases neuronal apoptosis in the hippocampus of mice, accompanied by increased expression of PCSK9 and activation of the apoptotic pathway [10], suggesting that PCSK9 might link abnormal lipid metabolismto AD.
In this study, our hypothesis is that PCSK9 might be associated with PC12 cell apoptosis. There have been several pieces of evidence to support our hypothesis. PCSK9 might regulate the expression of inflammatory factors induced by oxLDL through NF-κB [11], whereas the NF-κB/B cell lymphoma 2 (Bcl-2) signaling pathway is crucial in mediating apoptosis. The roles for PCSK9 in cerebellar granule neuronal apoptosis is also supported by the fact that the effects of wild-type and mutant PCSK9 on apoptosis are partially reversed by boc-Aspartyl(OMe)-fluoromethylketone, a caspase inhibitor. Moreover, it is possible that apoptotic mediators, such as caspase 3 and death receptor 6, are involved in neuronal apoptotic signaling induced by PCSK9 [12]. It was recently reported that PCSK9 promotes neuronal apoptosis through decreasing apolipoprotein E receptor 2 levels and deregulating anti-apoptotic signaling pathways [13]. However, further investigations are required to identify the precise PCSK9-related mechanism underlying neuronal apoptosis.
MATERIALS AND METHODS
Cell culture and treatments
Highly-differentiated rat PC12 pheochromocytoma cells that were induced by nerve growth factor were obtained from the Shanghai Cell Bank, Chinese Academy of Sciences (Shanghai, China) [14]. PC12 cells were cultured in Dulbecco’s modified eagle medium supplemented with 10% fetal bovine serum and 50 U/ml penicillin. Cultured cells were maintained at 37°C and 5% CO2 in a humidified environment. Human oxidized LDL was purchased from Guangzhou Yiyuan Biotech Co. Ltd., China. Sub-confluent PC12 cells were serum-starved for 24 h and then incubated with oxLDL for different time periods as indicated.
Oil red O staining
Intracytoplasmic lipid droplets were detected by oil red O staining. PC12 cells were stained with filtered oil red O solution at 37°C for 15 min after incubation with oxLDL for 24 h. After washing twice with distilled water, nuclear counterstaining was performed with hematoxylin solution for 30 s. Cells with positive staining (red) were observed via light microscope (Olympus) and then photographed using Image Pro-Plus 6.0 software (Media Cybernetics).
Morphological assessment of apoptotic cells
Morphological assessment of apoptotic PC12 cells was performed using Hoechst 33258 staining with the Boytime Staining Assay kit (Beytime Biotechnology Co., Ltd., China). After washing twice with phosphate-buffered saline (PBS), cells were stained with 0.5 ml Hoechst 33258 for 5 min at 37°C. The stained nuclei were washed twice with PBS, followed by fluorescence microscopy with an ultraviolet filter.
Reverse-transcription polymerase chain reaction (RT-PCR)
Total RNA was extracted from PC12 cells with HP Total RNA Kit (Omega, USA). cDNA was synthesized using a RevertAid First Strand cDNA Synthesis Kit (Thermo, USA). RT-PCR was prepared as follows: 95°C for 5 min, and then 95°C for 30 s, 57°C for 30 s, and 72°C for 30 s for 35 cycles, and 72°C for 5 min. Samples were separated on 2% agarose gel and analyzed using Gel-Pro Analyzer. Each sample was run and analyzed in triplicate. Primers were designed and synthesized by Shanghai Sangon Bioengineering Co., Ltd., China. These sequences of primers were: PCSK9 (forward) 5’- GGTGAGGGTGTCTATGCTGTC -3’, (reverse)5’- GGGCTCTAAGGTTTTCCACTTC -3’, and PCR product was 172bp; BACE1 (forward) 5’-GTTACAAGTTCGCCGTCTCAC-3’, (reverse)5’-CTCATCTGTCTGTGGAATGTTG-3’, and PCR product was 218 bp; and β-actin (forward) 5’- GAGAGGGAAATCGTGCGTGAC -3’, (reverse)5’ - CATCTGCTGGAAGGTGGACA -3’, and PCR product was 452 bp.
Western blot analysis
The cells were washed three times with PBS, incubated in 150 μL cell lysis buffer on ice for 30 min, followed by a collection of cell lysates and centrifugation at 10,000× g for 10 min at 4°C. The protein concentrations of cell lysates were measured using BCA protein assay kit (Beytime Biotechnology Co., Ltd., China) according to the manufacturer’s protocol. The protein was separated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis and electrotransferred to the polyvinylidene fluoride membrane. The membranes were incubated in a blocking buffer (20 mM Tris base, pH 7.6, 150 mM NaCl, 0.1% Tween-20, and 5% non-fat milk) for 2 h at 37°C and subsequently with the appropriate primary antibody [rabbit anti-rat Bax (1:400; Proteintech Group, USA), caspase3 (1:400; Proteintech Group, USA), caspase9 (1:400; Proteintech Group, USA), PCSK9 (1:400; Proteintech Group, USA), rabbit anti-rat BACE1 (1:3000; Abcam, USA), rabbit anti-rat Bcl-2 (1:400; Bioworld Technology, USA)] in blocking buffer at 4°C overnight. The membranes were then incubated with anti-rabbit horseradish peroxidase-conjugated secondary antibodies (1:1000; Auragene Bioscience, China), for 2 h. The proteins were visualized by chemiluminescence using BeyoECL Plus (Beytime Biotechnology Co., Ltd.,China).
siRNA transfection
The PCSK9 mRNA sequence was retrieved from GenBank (no. NM_199253). PCSK9 siRNA was synthesized by Guangzhou Rui Bo Biotechnology Co., Ltd., China. The PCSK9 siRNA sense strand was 5’- GGAGGAUAGCCUGGUUGAUdTdT-3’, and the antisense strand was 5’-dTdTCCUCCUAUCGGACCAACUA-3’. The negative control siRNA was provided by the same company. The PCSK9 siRNA was diluted in Opti-MEM I and added to the culture medium at a final concentration of 50 or 100 nM. The transfection of siRNA into PC12 cells was conducted using HiperFect Transfection Reagent (Qiagen, USA) according to the manufacturer’s instructions. After cy3-siRNA was transfected into PC12 cells for 6 h, the cells were rinsed twice with PBS to reduce background interference. Cy3 was scattered in the cytoplasm that could be detected by fluorescent microscopy. Successful transfection was indicated by red fluorescence. RT-PCR was performed 24 h after transfection to select the most effective siRNA concentration. The concentrations of PCSK9 siRNA for maximal effects were used for the subsequent experiments. PC12 cells were transfected with siRNAs for 48 h and then treated with 75 mg/L oxLDL for 24 h.
Flow cytometric analysis of apoptotic rates of PC12 cells by Annexin V/propidium iodide staining
Annexin V staining was performed with an apoptosis assay kit (Nanjing KeyGen Biotech Co., Ltd., China). The cells were gently washed twice with PBS, digested with 0.25% trypsin, and collected by centrifugation at 2,000× g for 5 min. The cells were incubated in 500 μl binding buffer containing 5 μl Annexin V-FITC and 5 μl propidium iodide for 15 min in a darkroom and immediately analyzed by flow cytometry (excitation wavelength 488 nm, emission wavelength 530 nm). Data were analyzed by flow cytometry software.
Enzyme-linked immunosorbent assay (ELISA)
ELISA kit (Fengxiang Biotechnology Co., Ltd., China) was used to detect the concentrations of Aβ40 and Aβ42 in the culture medium of PC12 cells. Briefly, the culture medium was collected and centrifuged at 2000× g at 4°C for 30 min. The supernatants were collected, diluted 1:5 in the standard dilution buffer of the kit, and assayed following the kit protocol. The fluorescent signal was detected in a microplate reader through 450 nm excitation filter. Each sample was measured in duplicate.
Statistical analysis
Data are presented as mean±SEM. Statistical analysis of data was performed with one-way ANOVA with Tukey-Kramer post-hoc testing. The results were plotted using GraphPad Prism5.0 software. Differences between the data sets are statistically significant at p < 0.05.
RESULTS
OxLDL treatment increased lipid accumulation, cell apoptosis, and PCSK9 expression in PC12 cells
Lipids were accumulated in the cytoplasm of PC12 cells after treatment with oxLDL. A gradual increase in Oil red O-stained lipids was observed in PC12 cells in response to treatment with increasing concentrations (25–100 mg/L) of oxLDL for 24 h (Fig. 1). This observation suggests that oxLDL induces lipid accumulation in PC12 cells in a concentration-dependent manner.
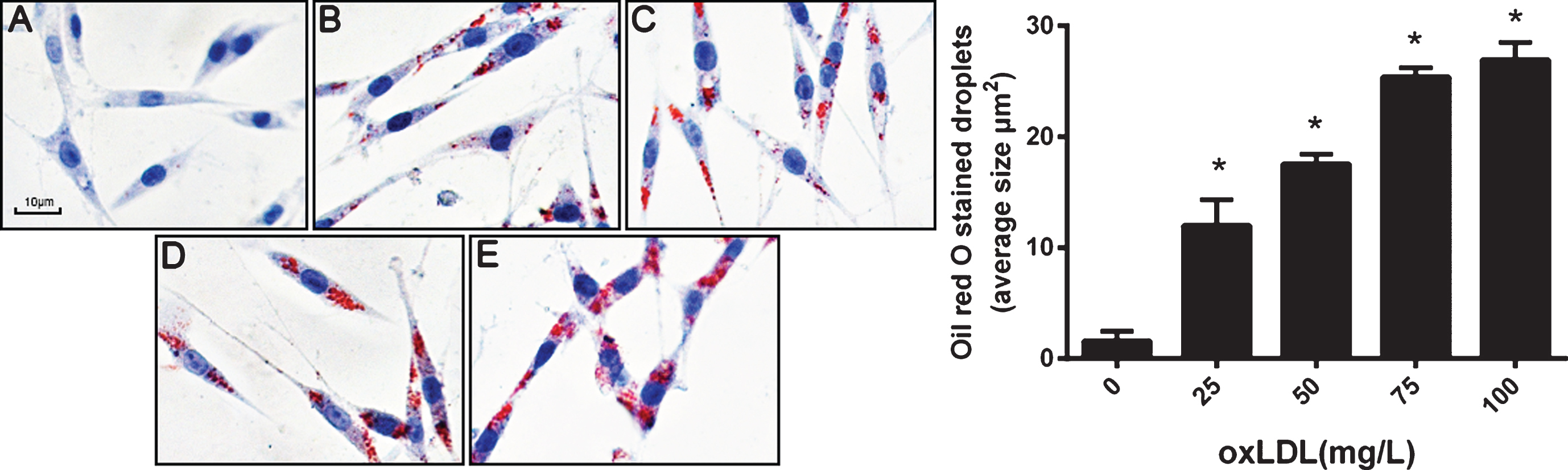
OxLDL increases the lipid accumulation in PC12 cells in a concentration-dependent manner. PC12 cells were treated with different concentrations of oxLDL and then subjected to oil red O staining. A–E images corresponding to PC12 cells treated with 0, 25, 50, 75, and 100 mg/L oxLDL (×400), respectively. Data represent three independent experiments (Five fields of vision were randomly selected for analysis in every independent experiment). One-way ANOVA with Tukey-Kramer post hoc test. *p < 0.05 versus 0 mg/L oxLDL.
PC12 cells were treated with oxLDL in a range of 25–100 mg/L for 24 h. Hoechst33258 staining showed a significant increase in the number of cells with nuclear condensation in response to treatment with oxLDL at 75 mg/L. After treatment with 100 mg/L oxLDL, apoptotic PC12 cell number was increased, but necrotic cell number was also increased likely due to its toxicity (Fig. 2). Therefore, in the subsequent experiments, only 75 mg /L oxLDL was used to treat PC12 cells.
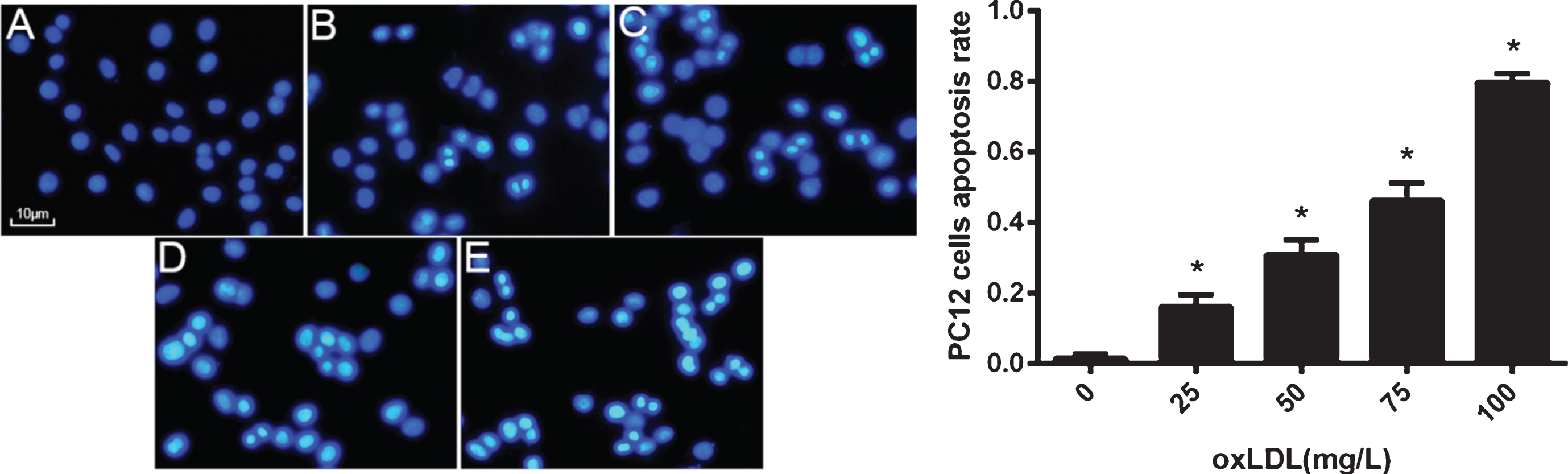
OxLDL induces PC12 cell apoptosis in a concentration-dependent manner. PC12 cells were treated with different concentrations of oxLDL and then subjected to Hoechst 33258 staining. A–E images corresponding to PC12 treated with 0, 25, 50, 75, and 100 mg/L oxLDL (×400), respectively. Data represent three independent experiments (Five fields of vision were randomly selected for analysis in every independent experiment). One-way ANOVA with Tukey-Kramer post hoc test. *p < 0.05 versus 0 mg/L oxLDL.
PC12 cells were treated with increasing concentrations of oxLDL (25–100 mg/l) for 24 h, followed by determining whether oxLDL upregulates the PCSK9 expression. The expression of PCSK9 mRNA and protein was analyzed using RT-PCR and western blotting, respectively. As shown in (Fig. 3), treatment with oxLDL increased the expression of PCSK9 mRNA and protein in a concentration-dependent manner. The peak of PCSK9 expression was observed at the oxLDL concentration of 100 mg/L (p < 0.05). The 75 mg/L oxLDL was selected for the subsequent experiments because of the high toxicity of oxLDL at 100 mg/L in PC12 cells. The above results showed that the increase in oxLDL concentrations resulted in increased lipid deposition and apoptosis in PC12 cells. Moreover, the expression of PCSK9 was also induced by oxLDL. A similar trend was observed among lipid deposition, cell apoptosis, and PCSK9 expression. Taken together, these results suggest the correlation between PCSK9 expression and PC12 cell apoptosis.
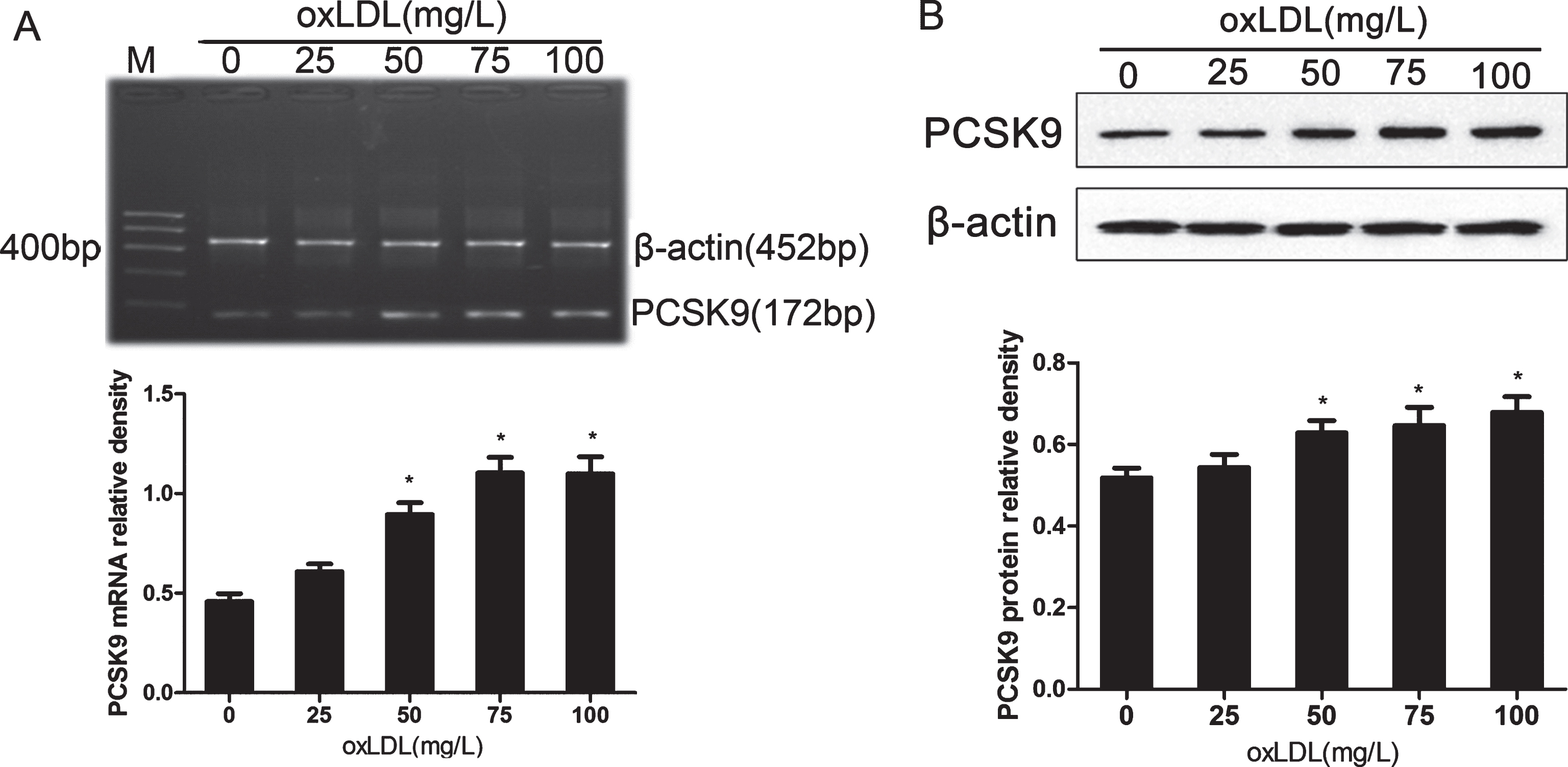
oxLDL upregulates PCSK9 mRNA and protein levels in PC12 cells in a concentration-dependent manner. A) Quantitation of PCSK9 mRNA in PC12 cells normalized to β-actin levels. B) Western blot analysis of PCSK9 levels in PC12 with β-actin as a loading control. Data are represented as mean±SEM from three independent experiments. One-way ANOVA with Tukey-Kramer post hoc test. *p < 0.05 versus 0 mg/L oxLDL.
PCSK9 siRNA inhibits oxLDL-induced PC12 cell apoptosis via the Bcl-2/Bax-Caspase (9, 3) signaling pathway
To investigate the roles of PCSK9 expression in BACE1 expression, amyloid-β protein precursor (AβPP)/Aβ metabolic pathway, and oxLDL-induced PC12 apoptosis, we developed the transfection of PCSK9 siRNA in PC12 cells. First, PC12 cells were transfected with different concentrations of siRNA for 24 h, followed by detection of PCSK9 mRNA with RT-PCR analysis. Our results revealed that PCSK9 siRNA at the concentration of 100 nM significantly inhibited the expression of the PCSK9 gene (Fig. 4).
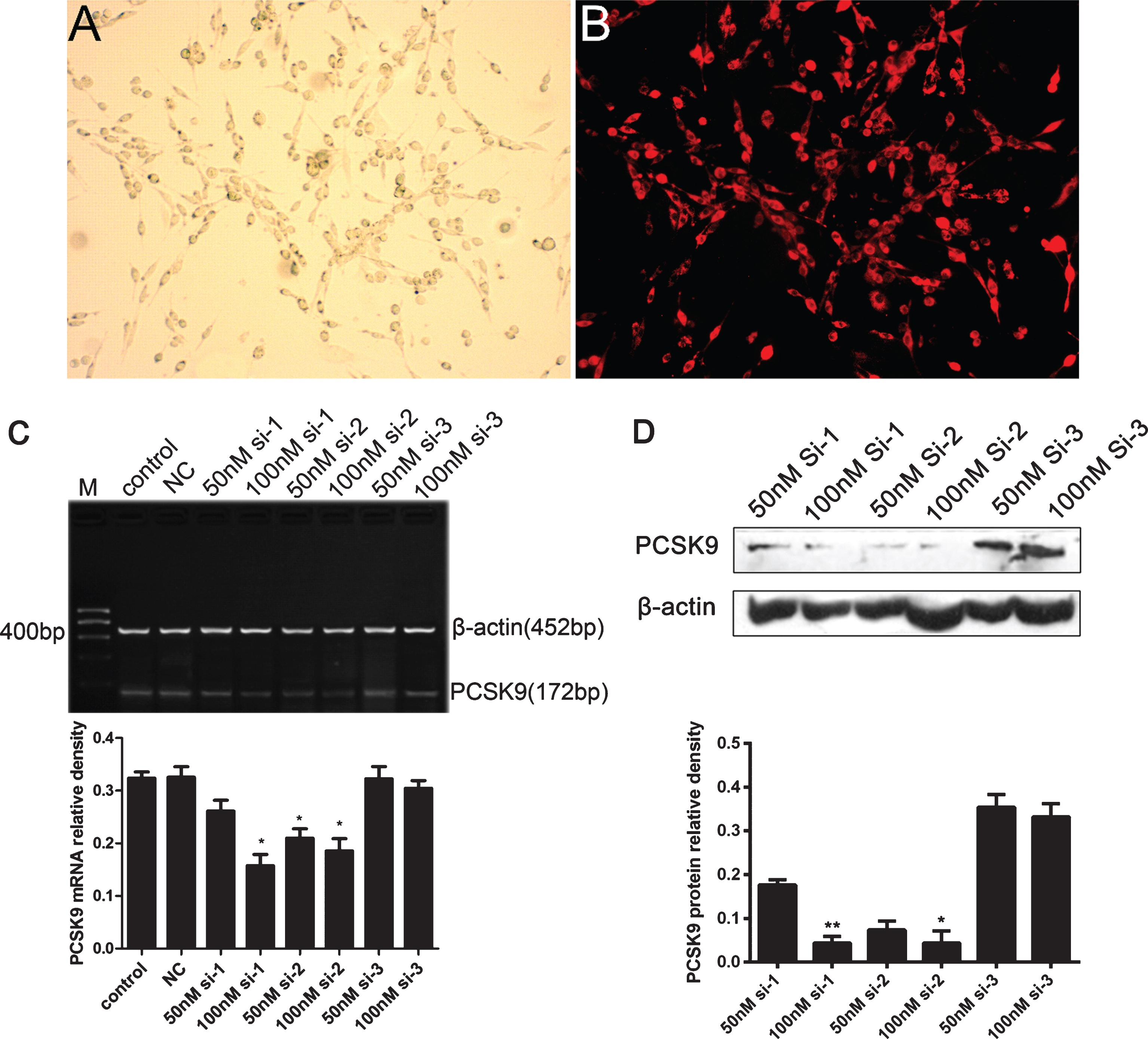
PCSK9 siRNA reduces the expression of PCSK9 in PC12 cells. A, B) PC12 cells were transfected with PCSK9 cy3-siRNA for 6 h. Transfection efficiency was then observed under the fluorescence microscope (×200). C) PC12 Cells were treated with control, NC, 50 nM, and 100 nM siRNA for 24 h and PCSK9 mRNA was detected by RT-PCR. *p < 0.05 versus control. D) PC12 Cells were treated with 50 nM, and 100 nM siRNAs for 24 h and PCSK9 protein was detected by western blot. **p < 0.01 versus 50 nM si-1, *p < 0.05 versus 50 nM si-2. Data are expressed as means±SEM from three independent experiments. One-way ANOVA with Tukey-Kramer post hoc test.
We then evaluated the effect of PCSK9 siRNA transfection on PC12 cell apoptosis. Pretreatment of PC12 cells with PCSK9 siRNA significantly reduced the number of apoptotic cells in response to oxLDL. These effects of PCSK9 siRNA were confirmed by Hoechst 33258 staining (Fig. 5) and flow cytometry analysis of Annexin V-FILC-stained apoptotic cells (Fig. 6).
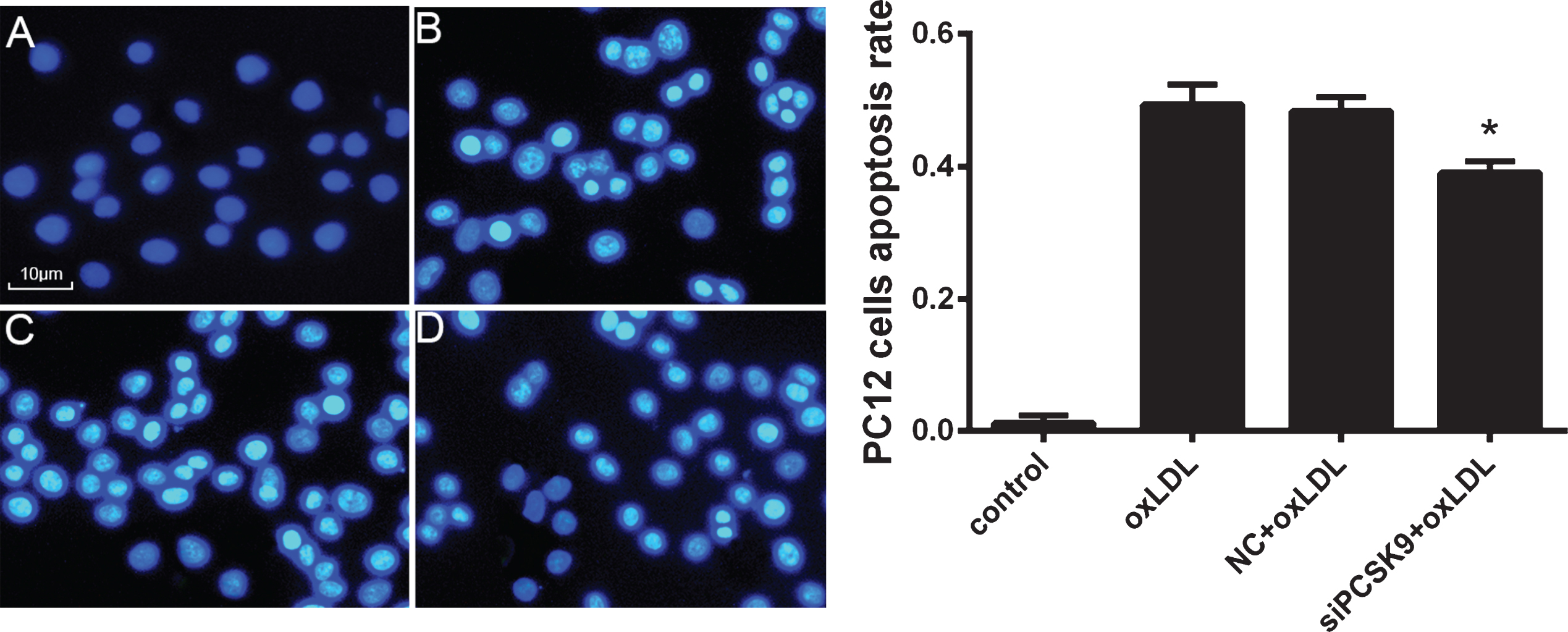
PCSK9 siRNA reduces the apoptosis of oxLDL-induced PC12 cells. After transfection with 100 nmol/l PCSK9 siRNA for 24 h, PC12 cells were treated with 75 mg/l oxLDL for 24 h. A-D represent PC12 treated with control, 75 mg/L oxLDL, NC +75 mg/L oxLDL, siPCSK9+75 mg/L oxLDL. *p < 0.05 versus NC+oxLDL. Data represent three independent experiments. One-way ANOVA with Tukey-Kramer post hoc test.
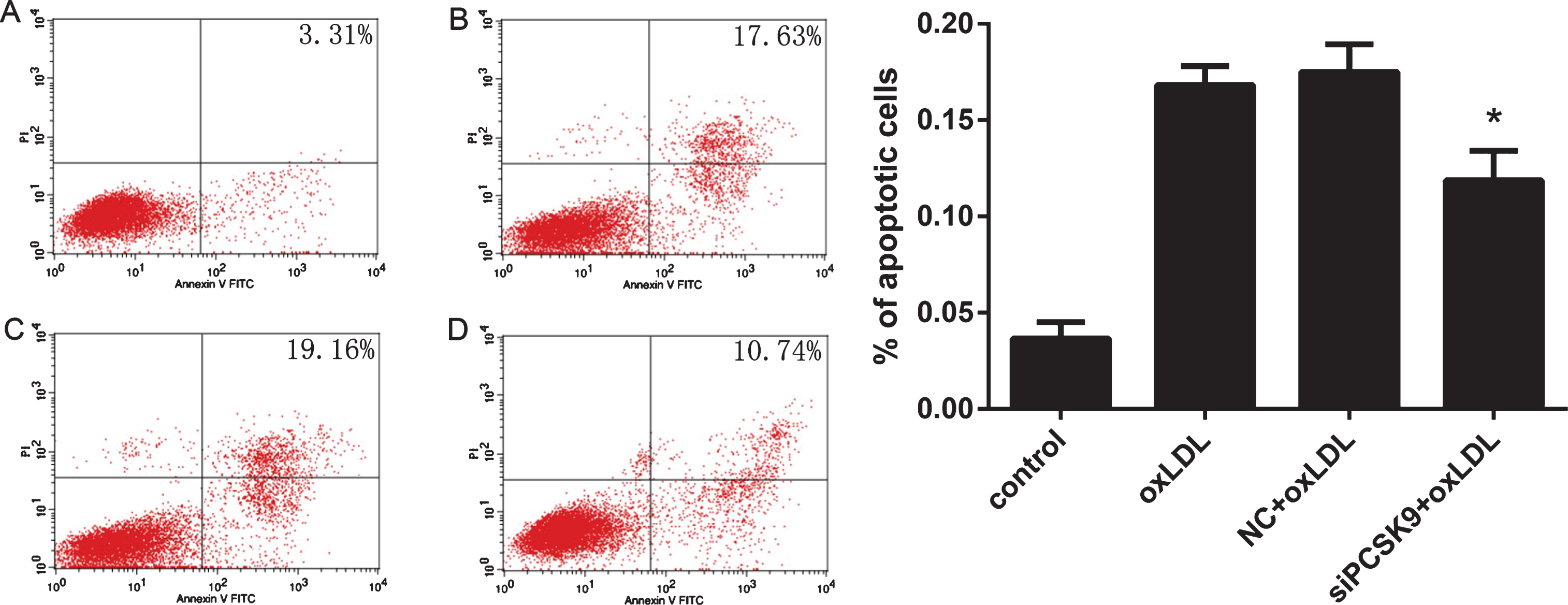
Detection of PC12 cell apoptosis by flow V-FITC Apoptosis Detection Kit for annexin V/propidium iodide cell labeling. A–D images corresponding to PC12 cells treated with control, 75 mg/L oxLDL, NC +75 mg/L oxLDL, and siPCSK9+75 mg/L oxLDL, respectively. Data represent three independent experiments. One-way ANOVA with Tukey-Kramer post hoc test. *p < 0.05 versus NC+oxLDL.
To further investigate the effect of PCSK9 siRNA on oxLDL-induced PC12 cell apoptosis, we determined the expression of apoptosis-related protein caspase3, caspase9, Bcl-2, and Bax by western blot analysis. The levels of caspase3, caspase 9, and Bax protein were significantly reduced in response to treatment with PCSK9 siRNA, whereas the expression of Bcl-2 was increased compared with that in the negative control (NC)+oxLDL group (p < 0.05) (Fig. 7). These results suggest that PCSK9 promotes apoptosis likely through the activation of the Bcl-2/Bax-Caspase (9, 3) signal pathway in PC12 cells.
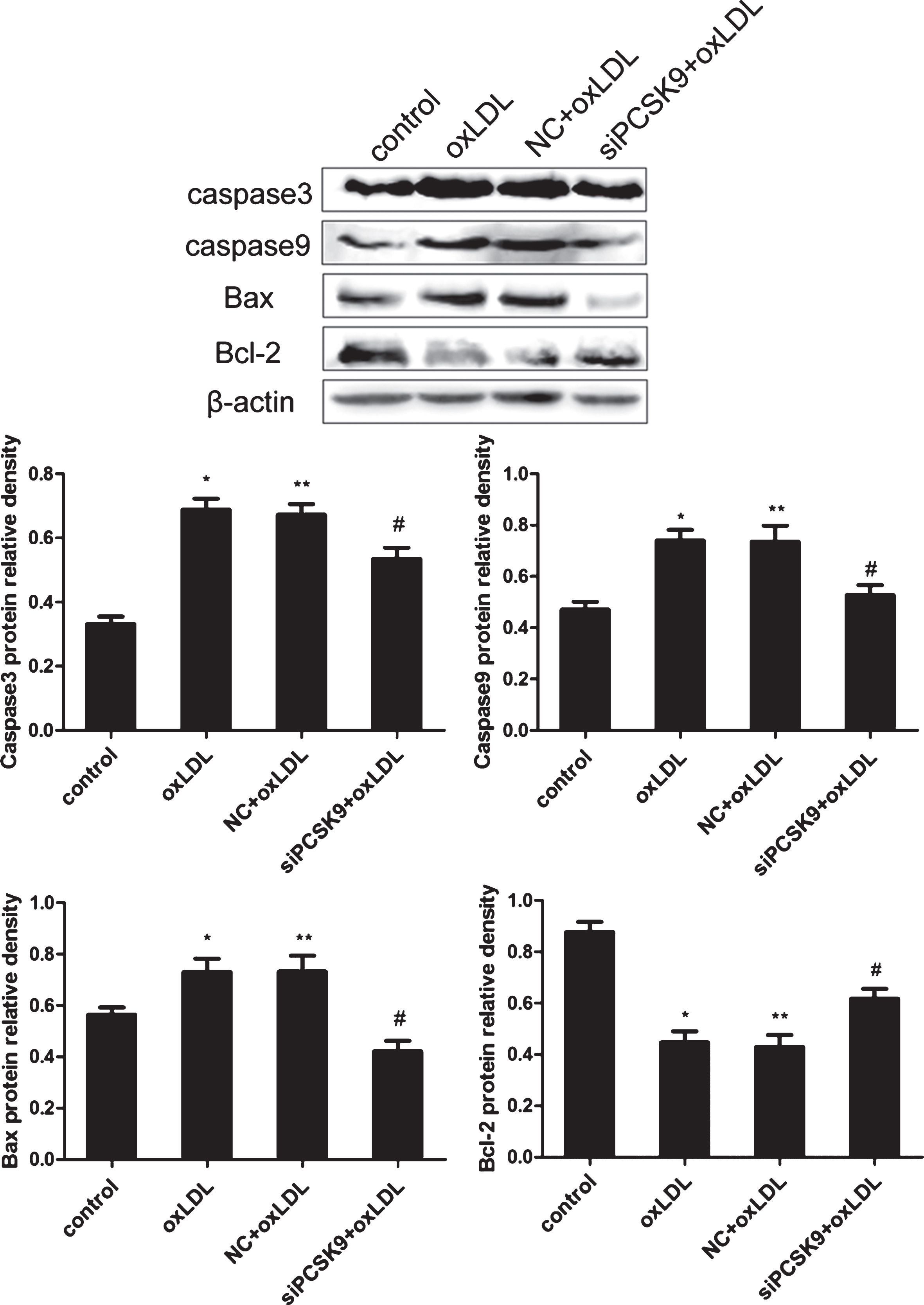
PCSK9 siRNA inhibits the expression of caspase3, caspase9, and Bax, while upregulates Bcl-2 in oxLDL-treatedPC12 cells. PC12 cells transfected with 100 nM PCSK9 siRNA were treated with 75 mg/L oxLDL for 24 h. The expression of caspase3, caspase9, Bcl-2, and Bax protein was determined by western blot analysis. Data are shown as mean±SEM of three or more replicate experiments. One-way ANOVA with Tukey-Kramer post hoc test. *p < 0.05 versus control; **p > 0.05 versus oxLDL; #p < 0.05 versus NC+oxLDL.
PCSK9 siRNA suppresses the oxLDL-induced downregulation of BACE1, Aβ40, and Aβ42 in PC12 cells
PC12 cells were incubated with various concentrations of oxLDL (0, 25, 50, 75, and 100 μg/ml) for 24 h. BACE1 mRNA and protein were then detected by RT-PCR and western blot analyses, respectively. The results revealed that oxLDL treatment reduced the expression of BACE1 mRNA and protein in a concentration-dependent manner (Fig. 8A, B). The contents of Aβ40 and Aβ42 in PC12 cells were downregulated with increasing concentrations of oxLDL as detected by ELISA (p < 0.05) (Fig. 9A). Treatment with a high concentration of oxLDL significantly increased PC12 cell apoptosis and PCSK9 expression but reduced the expression of BACE1 and Aβ contents. These results revealed a negative correlation between PCSK9 and BACE1 expression. We further tested the effect of PCSK9 siRNA on the expression of BACE1 in PC12 cells. PC12 cells were transfected with 100 nM PCSK9 siRNA, cultured in medium for 24 h and then treated with oxLDL (75 mg/L) for 24 h. Western blot analysis revealed a significant increase in BACE1 expression compared with that treated with only 75 mg/L oxLDL (Fig. 8C). Aβ40 and Aβ42 contents were upregulated by PCSK9 siRNA in oxLDL-treated PC12 cells as determined by ELISA (Fig. 9B). These results suggest that PCSK9 may inhibit the expression of BACE1 that controls the production of Aβ40 and Aβ42.
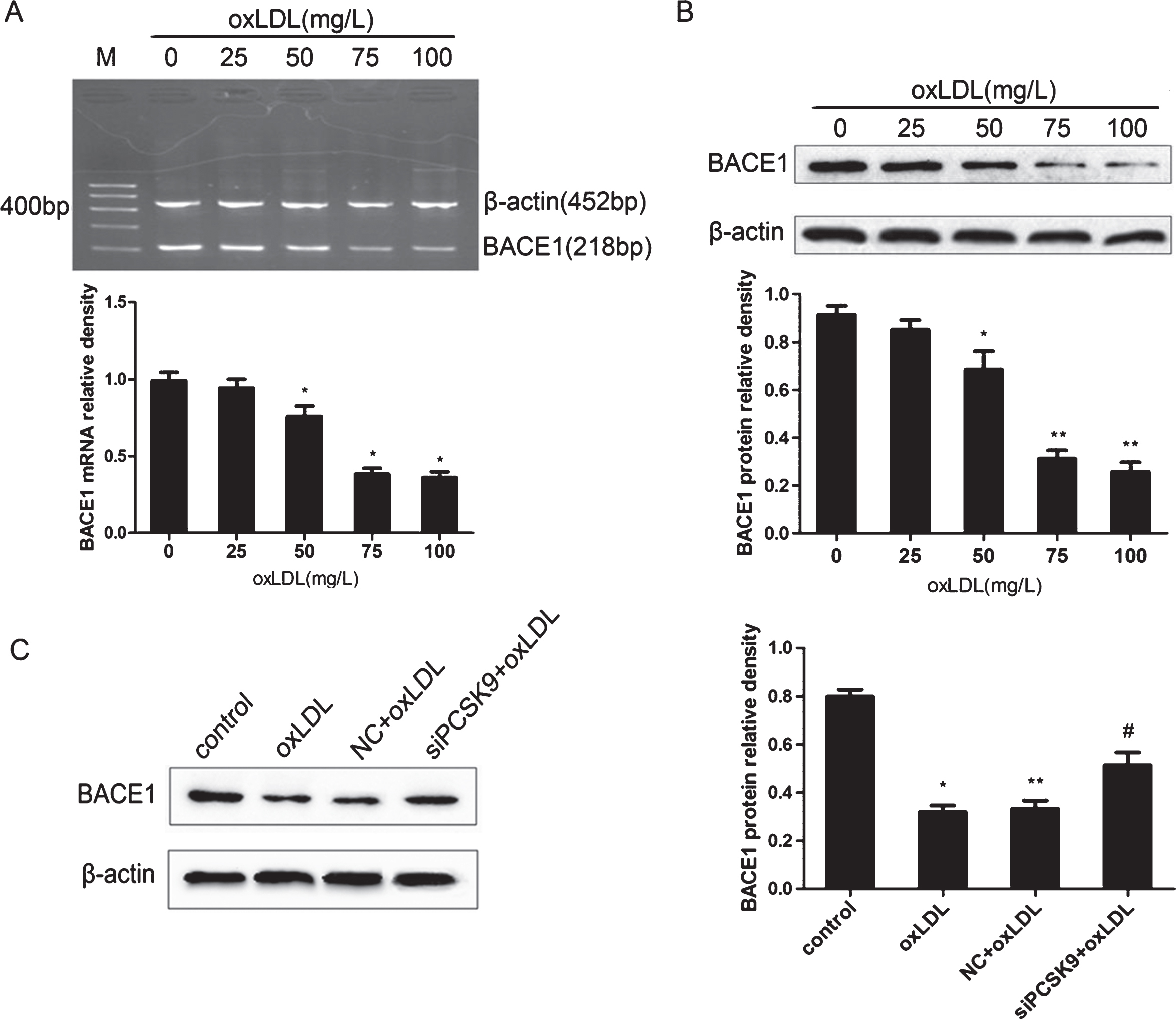
PCSK9 siRNA inhibits the expression of BACE1 in oxLDL-induced PC12. PC12 cells were treated with oxLDL at different concentrations (0, 25, 50, 75, and 100 mg/L) for 24 h. BACE1 mRNA and protein were detected by RT-PCR (A) and western blot analysis (B). After transfection with 100 nM PCSK9 siRNA, PC12 cells were treated with 75 mg/L oxLDL for 24 h. *p < 0.05 versus 0 mg/L oxLDL; **p < 0.01 versus 0 mg/L oxLDL. The protein levels of BACE1 were detected by western blot analysis (C). *p < 0.05 versus control; **p > 0.05 versus oxLDL; #p < 0.05 versus NC+oxLDL. Data are shown as mean±SEM of three or more replicate experiment. One-way ANOVA with Tukey-Kramer post hoc test.
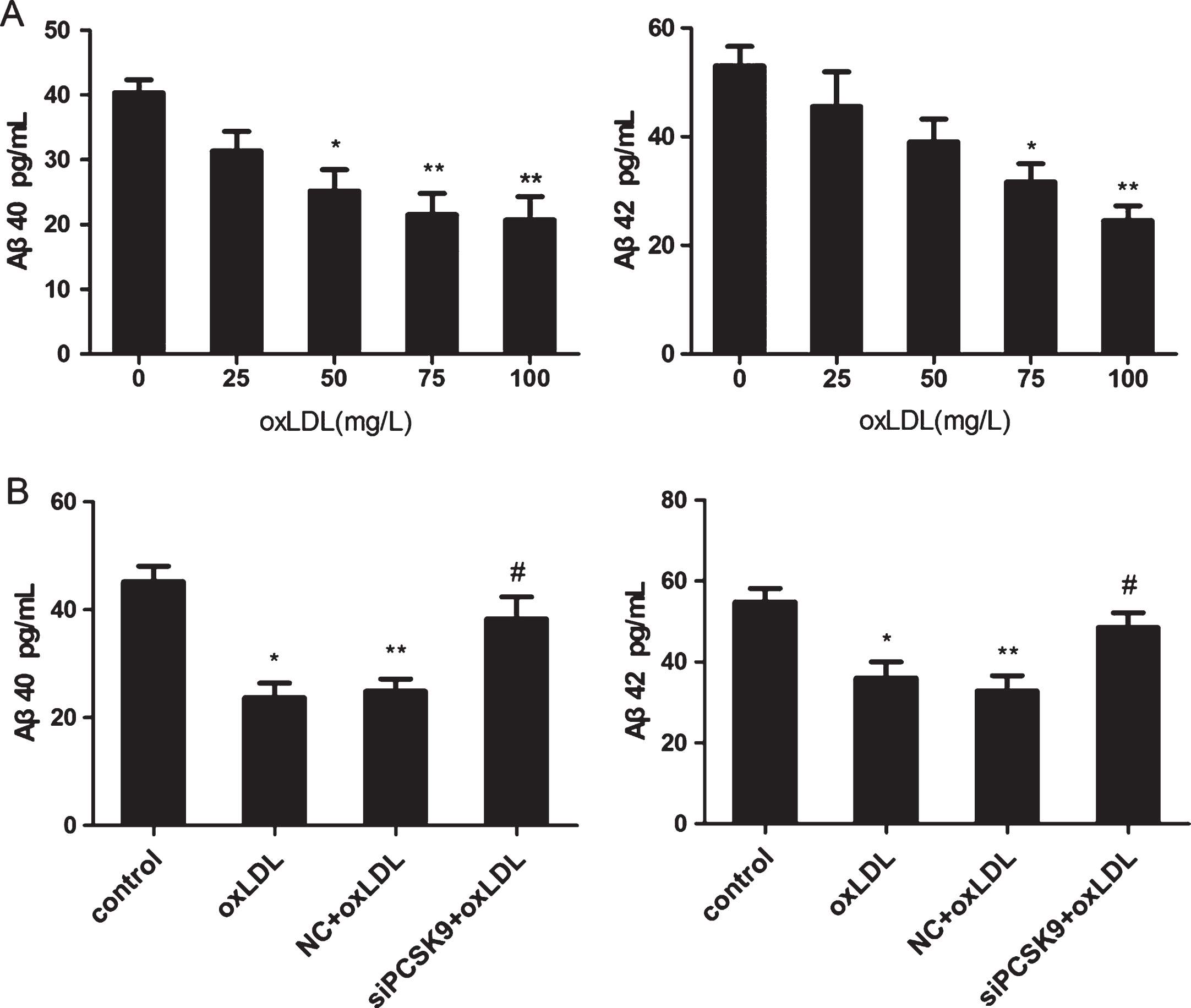
PCSK9 siRNA inhibits the Aβ40 and Aβ42 contents in oxLDL-induced PC12. A) PC12 cells were treated with 0, 25, 50, 75, and 100 mg/L oxLDL for 24 h. Protein levels of Aβ40 and Aβ42 were detected by ELISA. *p < 0.05 versus 0 mg/L oxLDL; **p < 0.01 versus 0 mg/L oxLDL. B) After transfection with 100 nM PCSK9 siRNA, PC12 cells were treated with 75 mg/L oxLDL for 24 h. The expression of Aβ40 and Aβ42 were determined by ELISA. *p < 0.05 versus control; **p > 0.05 versus oxLDL; #p < 0.05 versus NC+oxLDL. Data are shown as mean±SEM of three or more replicate experiments. One-way ANOVA with Tukey-Kramer post hoc test.
DISCUSSION
Recent clinical trials using antibodies against PCSK9 to block PCSK9 interaction with LDLR have shown a significant effect in lowering LDL cholesterol levels [15–17]. However, the results of several studies have suggested that neurocognitive adverse events may occur more frequently in individuals receiving PCSK9 antibodies [15, 18]. These neurocognitive adverse events suggest a potential role for PCSK9 in the nervous system. PCSK9 is also known as neural apoptosis-regulated convertase-1, which was identified via the cloning of cDNAs in primary cerebellar neurons after apoptosis induced by serum deprivationand was implicated in the differentiation of cortical neurons [19]. However, the precise role of PCSK9 in the nervous system still remains to be explored.
In ApoE–/–mice fed a high-fat diet, we found that PCSK9 expression was higher in the hippocampus than those in ApoE–/–mice fed a normal chow diet [10]. Correspondingly, neuronal apoptosis was markedly enhanced in the hippocampus of mice fed a high-fat diet when compared with mice fed a normal chow diet. In this paper, we tried to further elucidate the mechanism underlying hyperlipidemia-induced neuronal apoptosis that was found in animal studies. Therefore, PC12 cells were cultured and treated with different concentrations (25 to 100 μg/mL) of oxLDL for 24 h. As expected, oxLDL induced lipid accumulation and apoptosis in PC12 cells in a concentration-dependent manner. It also increased PCSK9 expression. The positive correlation between high PCSK9 expression and high apoptotic rate in PC12 cells suggests that PCSK9 may be involved in oxLDL-induced neuronal apoptosis.
Subsequently, we investigated the role of PCSK9 in oxLDL-induced apoptosis in PC12 cells using RNA interference technology and found that PCSK9 siRNA significantly reduced the apoptosis of PC12 cells induced with oxLDL. Moreover, the expression of pro-apoptotic protein caspase3, caspase9, and Bax was significantly decreased in response to treatment with PCSK9 siRNA, whereas the expression of anti-apoptotic protein Bcl-2 was increased. These results suggest that PCSK9 promotes apoptosis likely through the activation of the Bcl-2/Bax-Caspase (9, 3) signal pathway in PC12 cells. Remarkably, oxLDL upregulates the PCSK9 expression in human umbilical vein endothelial cells, whereas PCSK9 siRNA inhibits oxLDL-induced apoptosis of human umbilical vein endothelial cells through the Bcl/Bax–caspase 9/caspase 3 pathway [20]. In addition, Kysenius [13] reported that PCSK9 potentiated apoptosis of cerebellar granule neurons via modulation of ApoER2 levels and the related anti-apoptotic signaling pathways. In summary, PCSK9 may exert a pro-apoptotic effect in various cells.
AD is characterized by a progressive loss of neurons, which is closely associated with apoptosis of neurons induced by many factors, such as Aβ and oxidative stress. Two different metabolic pathways are involved in the processing of AβPP: (1) AβPP is cleaved by α-secretase to generate soluble AβPP alpha and the transmembrane C-terminal fragment alpha, which is further cleaved by γ-secretase to a C-terminal 83-amino acid fragment (C83); and (2) AβPP is cleaved by the β-site AβPP-cleaving enzyme 1 (BACE1) and γ-secretase to generate insoluble Aβ. The former pathway is generally the normal metabolism of AβPP. Under pathological conditions, the latter pathway is activated to produce insoluble Aβ, leading to AD. The BACE1 level is increased in many sporadic AD cases. Consistent with an increase in BACE1, higher concentration of Aβ produced by BACE1 cleavage may induce neuronal apoptosis and subsequently lead to AD. Jonas et al. [21] and Ko et al. [22] observed that the levels of BACE1 and Aβ were increased in the brains of PCSK9 (–/–) mice. Overexpression of PCSK9 decreased the level of BACE1 and small interfering RNA-mediated downregulation of PCSK9 increased the level of BACE1 in CHO cells. These results confirmed that secreted PCSK9 may stimulate the degradation of BACE1. These findings revealed that PCSK9 possibly reduced Aβ production to prevent neuronal apoptosis through degradation of BACE1 in nerve cells. However, Liu et al. [23] reported that there was no correlation between the levels of PCSK9 and BACE1 expression. Thus, further investigation is required to determine whether PCSK9 affects BACE1activity.
Our current study has revealed that PCSK9 may be negatively correlated with BACE1. Both BACE1 mRNA and protein levels and Aβ contents were reduced in a concentration-dependent manner in oxLDL-stimulated PC12 cells. At the same time, apoptosis and PCSK9 expression were significantly increased in oxLDL-stimulated PC12 cells. In addition, the levels of BACE1 and Aβ were significantly upregulated after transfection with PCSK9 siRNA. These results suggest that PCSK9 can inhibit the expression of BACE1 and affect the production of Aβ.
In summary, hyperlipidemia increases the lipid content in the brain, particularly oxLDL content. Increased oxLDL may upregulate PCSK9 expression in nerve cells. Neuronal apoptosis can be induced through the Bcl-2/Bax-Caspase (9, 3) signaling pathway when the PCSK9 expression is upregulated. However, through degradation of BACE1, PCSK9 decreases Aβ production via inhibiting the AβPP/Aβ metabolic pathway and ultimately decreases Aβ-induced neuronal apoptosis. Nevertheless, our results conclude that PCSK9 promotes PC12 cell apoptosis induced by oxLDL. We speculate that PCSK9 exerts an important regulatory effect on neuronal apoptosis, which may contribute to the slow progressionof AD.
Footnotes
ACKNOWLEDGMENTS
This work was supported by the grants from the National Natural Science Foundation of China (81200217, 81370376), the Natural Science Foundation of Hunan province (2015JJ4097), the Project of Science and Technology Department of Hunan Province (2015JC3081, 2014FJ3015), the Educational Department of Hunan Province Fund for Distinguished Young Scholars (16B228), the Hunan Provincial Innovation Foundation For Postgraduate (CX2016B477), and by the Construct Program of the Key Discipline in Hunan Province.