
Abstract
Administration of sevoflurane (SEVO) may induce learning and memory deficits, which increases the chances of developing Alzheimer’s disease. Here, we studied the effects of SEVO exposure on rats with a focus on the role of insulin-like growth factor (IGF) signaling. SEVO exposure significantly increased neuron cell apoptosis, and caused poor performance of the rats in behavior tests, by suppressing IGF-1 receptor (IGF1R). Bioinformatic analysis predicted microRNA(miR)-223-3p as an IGF1R-binding miRNA, the level of which increased in neurons after exposure to SEVO. In vitro, miR-223-3p suppressed the translation of IGF1R in neural cells. Moreover, transfection with antisense of miR-223-3p significantly attenuated SEVO exposure-induced neuron cell apoptosis. Taken together, these data suggest that SEVO-induced miR-223-3p upregulation suppresses IGF1R to impair IGF signaling, which subsequently leads to learning and memory impairments.
INTRODUCTION
The insulin-like growth factor-1 (IGF-1) receptor signaling pathway initiates with binding of IGF-1 and its cell-surface receptor IGF-1 receptor (IGF1R), which activates phosphatidylinositol–3 kinase (PI3K)/Protein kinase B (Akt) or extracellular signal-regulated kinase (ERK)/mitogen-Activated Protein Kinase (MAPK) signaling pathway, to promote cell replication and suppress apoptotic cell death [1–3]. Interaction among cells using IGF-1/IGF1R signaling cascades plays critical roles in the neuronal cell survival against stress and damage [4, 5]. Besides IGF-1, IGF-2 also binds to and function through IGF-1R.
Sevoflurane (SEVO) has an adverse effect on the development of the brain and causes learning and memory impairments, which increase the chances of developing Alzheimer’s disease [6–12]. The detrimental effects of SEVO have been widely reported. For example, gamma-aminobutyric acid (GABA) was shown to be induced by SEVO, which subsequently suppressed N-methyl-D-aspartate (NMDA) receptors to cause extracellular calcium ion inflow, overload and retention of neuronal calcium, both leading to establishment of neurotoxicity and long-term impairment of learning and memory [13–15]. On the other hand, SEVO-induced suppression of NMDA receptors also resulted in decreases in extracellular glutamate concentration and formation of synapses [16].
Currently, the effects of SEVO on IGF signaling, which plays critical roles in neuronal cell survival against apoptotic cell death, is unknown. Also, a role for microRNAs (miRNAs) in these processes has not been acknowledged. Here, we studied the effects of SEVO exposure on rats, with a focus on the role of IGF signaling.
We found that SEVO exposure significantly increased neuron cell apoptosis, and caused poor performance of the rats in behavior tests, through suppression of IGF-1R. Bioinformatic analysis predicted miR-223-3p as an IGF1R-binding miRNA, the level of which increased in neurons after exposure to SEVO. In vitro, miR-223-3p suppressed the translation of IGF1R in neural cells. Moreover, transfection with antisense of miR-223-3p significantly attenuated SEVO exposure-induced neuron cell apoptosis.
MATERIALS AND METHODS
Protocol approval
All the experimental methods and ethic animal studies have been approved by the research committee of Shanghai Jiao Tong University.
Animals
All experiments were performed according to the Experimental Guideline of Shanghai Jiao Tong University. Female Sprague-Dawley rats were used at 12 weeks of age (Shanghai Laboratory Animal Center, Shanghai, China) in the current study. The control rats were placed in a chamber with continuously flushed air, while the SEVO-grouped rats were placed in a chamber with 5% SEVO and air. For both control and experimental group, a low rate of 2 L/min and a duration of 4 h of flushing was applied.
Cell culture and treatment
A human cortical neuron cell line, HCN-2 (ATCC, Rockville, MD, USA), was maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 15% fetal bovine serum (Invitrogen, Beijing, China) in a 37°C humidified chamber flushed with 5% CO2. For in vitro studies, HCN-2 cells were exposed to SEVO of certain concentrations in a gas-tight chamber placed in the incubator.
Plasmids and viruses
The miR-223-3p or null or antisense for miR-223-3p (as-miR-223-3p) plasmids were produced using the corresponding sequence. Sequence for miR-223-3p: 5’-ugucaguuugucaaauacccc-3’; Sequence for as-miR-223-3p: 5’-ggggtutttgucuuuctgucu-3’.
ELISA
The cells or rat hippocampus were homogenized in protein lysis buffer, after which the concentration of the total protein was determined in a BCA protein assay (Bio-rad, Shanghai, China). ELISA for IGF-1 and IGF1R was performed, using specific ELISA kits (R&D Biosystems, San Jose, CA, USA).
Cleavage Caspase 3 staining and quantification
The rat hippocampus or cultured HCN-2 cells were stained with a polyclonal rabbit anti-Cleavage Caspase 3 (casp3) antibody, a polyclonal rabbit anti-Glut1 (Abcam, Beijing, China). The nuclei were stained with DAPI (Sigma-Aldrich, Shanghai, China). Quantification was done manually by two independent researchers.
Quantitative PCR (RT-qPCR)
Total RNA was extracted using miRNeasy mini kit (Qiagen, Hilden, Germany), and then reversely transcribed to complementary DNA (cDNA) using miScript II RT Kit (Qiagen). Quantitative PCR was performed ABI 7700 System (TaqMan, Beijing, China). Values of genes were determined by sequential normalization to β-actin and the experimental controls.
Bioinformatics and dual luciferase-reporter assay
On line prediction of the miRNA-binding targets used TargetSan website [17]. Luciferase-reporters including the wildtype and the mutate 3’-UTR of IGF1R were constructed (Promega, Shanghai, China) and were applied in a dual-luciferase reporter gene assay kit (Promega).
Morris water maze test
For Morris water maze test, rats were tested for their capability of seeking and perching on the hidden platform. The time for the trained rats to reach the platform (latency) and the path lengths that they took were both recorded by software.
Plus-Maze discriminative avoidance task (PM-DAT)
In a PM-DAT, the rats were exposed to both light illumination and blowing cold air. After the training, rats were placed in the same position in the same room for 3 min without any aversive stimuli. The percentage of time spent in the aversive enclosed arm during training and testing was recorded respectively for assessment of learning and memory.
Statistics
Using GraphPad Prism 7 (GraphPad Software, San Diego, CA, USA), statistical analysis was done using one-way ANOVA with a Bonferroni correction, followed by Fisher’s Exact Test upon necessity. All values are depicted as mean±standard deviation from 5 individuals and are considered significant if p < 0.05.
RESULTS
SEVO exposure causes poor behavior and increased neuronal cell apoptosis of the rats
Rats were randomly separated into two groups. For a duration of 4 h, the control group (CTL; n = 5) was exposed to normal gas, while the SEVO group was exposed to 5% SEVO. At different time points after treatment, Morris water maze test was performed, showing that SEVO exposure caused the rats to show worse performance in both escape latency (Fig. 1A) and path length (Fig. 1B) gradually with time. At 6 weeks after treatment, the rats were also examined in an PM-DAT, when we found that SEVO exposure significantly increased the time spent in aversive arm at training period (Fig. 1C), and in time spent in open arm at testing period (Fig. 1D). These results suggest that both learning and memory of the rats are compromised by SEVO exposure. Cleavage Caspase 3 (casp3) is the most reliable marker for apoptotic cells. Therefore, we performed casp3 and Glut1 (for determining different brain tissue) staining on rat hippocampus at weeks after SEVO treatment. We detected significant increases in casp3 + apoptotic neuronal cells the hippocampus, shown by quantification (Fig. 1E), and by representative images (Fig. 1F). Thus, SEVO exposure causes poor behavior and increased neuronal cell apoptosis of the rats. Interaction among cells using IGF-1/IGF1R signaling cascades plays critical roles in the neuronal cell survival against stress and damage. In order to examine if IGF-1/IGF1R signaling cascades may be involved in the response of the body to the SEVO exposure, we examined the levels of IGF-1 and IGF1R by ELISA, showing that both significantly reduced in brain tissue after SEVO (Fig. 1G).
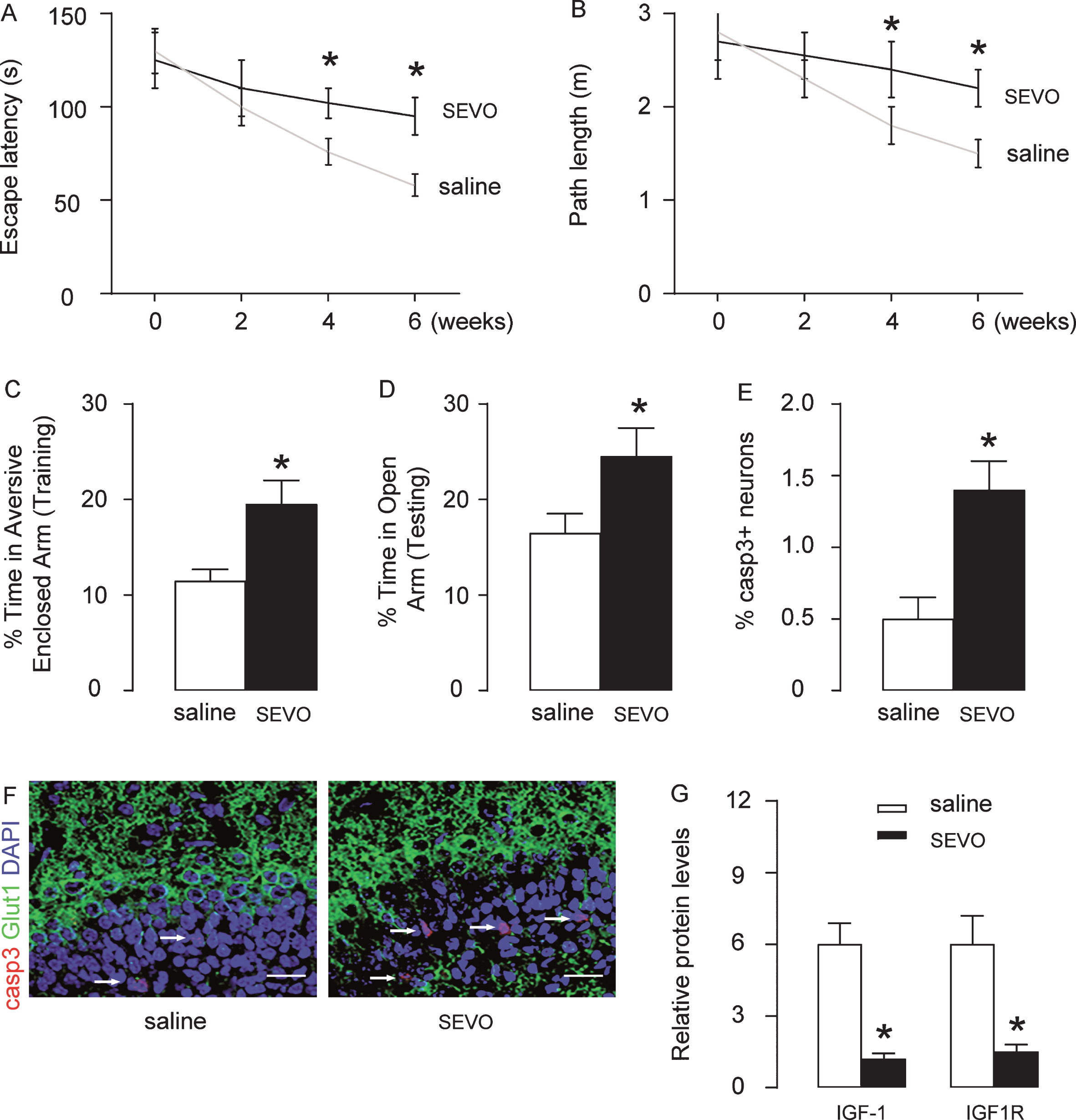
SEVO exposure causes poor behavior and increased neuronal cell apoptosis of the rats. Rats were randomly separated into two groups. For a duration of 4 h, the control group (CTL; n = 5) was exposed to normal gas, while the SEVO group was exposed to 5% SEVO. Morris water maze test: A) Escape latency; B) Path length. PM-DAT at 6 weeks after SEVO exposure: C) Time spent in aversive arm at training period; D) Time spent in open arm at testing period. Casp3 staining in the hippocampal area of rat brain, 6 weeks after SEVO exposure, by quantification (E) and by representative images (F). G) ELISA for IGF-1 and IGF1R in brain tissue, 6 weeks after SEVO exposure. *p < 0.05. N = 5. Scale bars are 20μm.
SEVO increases neuronal cell apoptosis in vitro and reduces IGF1R
In order to examine if SEVO-induced changes in IGF-1/IGF1R levels occurred in neuronal cells, we treated HCV-2 neural cells with different doses of SEVO in vitro for 48 h. We found that SEVO dose-dependently increased the casp3 + cells, shown by representative images (Fig. 2A), and by quantification (Fig. 2B). Moreover, SEVO dose-dependently reduced the total number of the cells (Fig. 2C). IGF-1 and IGF1R were both examined by ELISA, and showed no alteration in protein levels of IGF-1 by SEVO (Fig. 2D), but dose-dependent reduction in protein levels of IGF1Rby SEVO (Fig. 2E). Hence, the SEVO-induced changes in IGF-1 levels may occur in cells other than neurons, while IGF1R levels may be directly affected at neurons. Next, we examined mRNA level of IGF-1R, and surprisingly found that the mRNA levels of IGF1R remained unchanged by SEVO (Fig. 2F), suggesting that SEVO may alter post-transcriptional control of IGF-1R.

SEVO increases neuronal cell apoptosis in vitro and reduces IGF1R. HCV-2 neuronal cells were exposed to different doses of SEVO for 48 h and then for analysis. Casp3 + staining on HCN-2 cells, shown by representative images (A), and by quantification (B). C) The quantification of total cell number. ELISA for IGF-1 (D) and IGF1R (E) in treated HCN-2 cells. F) RT-qPCR for IGF1R mRNA. *p < 0.05. NS, non-significant. N = 5. Scale bars are 20μm.
SEVO upregulates miR-223-3p in neuronal cells
In order to determine the exact post-transcriptional control of IGF1Rby SEVO, we did a miRNA assay on 5% SEVO-treated HCN-2 cells versus control cells and highlighted IGF1R-binding miRNAs (Fig. 3A). We found that among these IGF1R-binding miRNAs, only miR-223-3p significantly increased after exposure to 5% SEVO (Fig. 3B). Moreover, miR-223-3p seemed to be an IGF1R-binding miRNA cross rats (Fig. 3C) and humans (Fig. 3D). Thus, our data on miR-223-3p should be applicable to both human and rat neuronal cells.
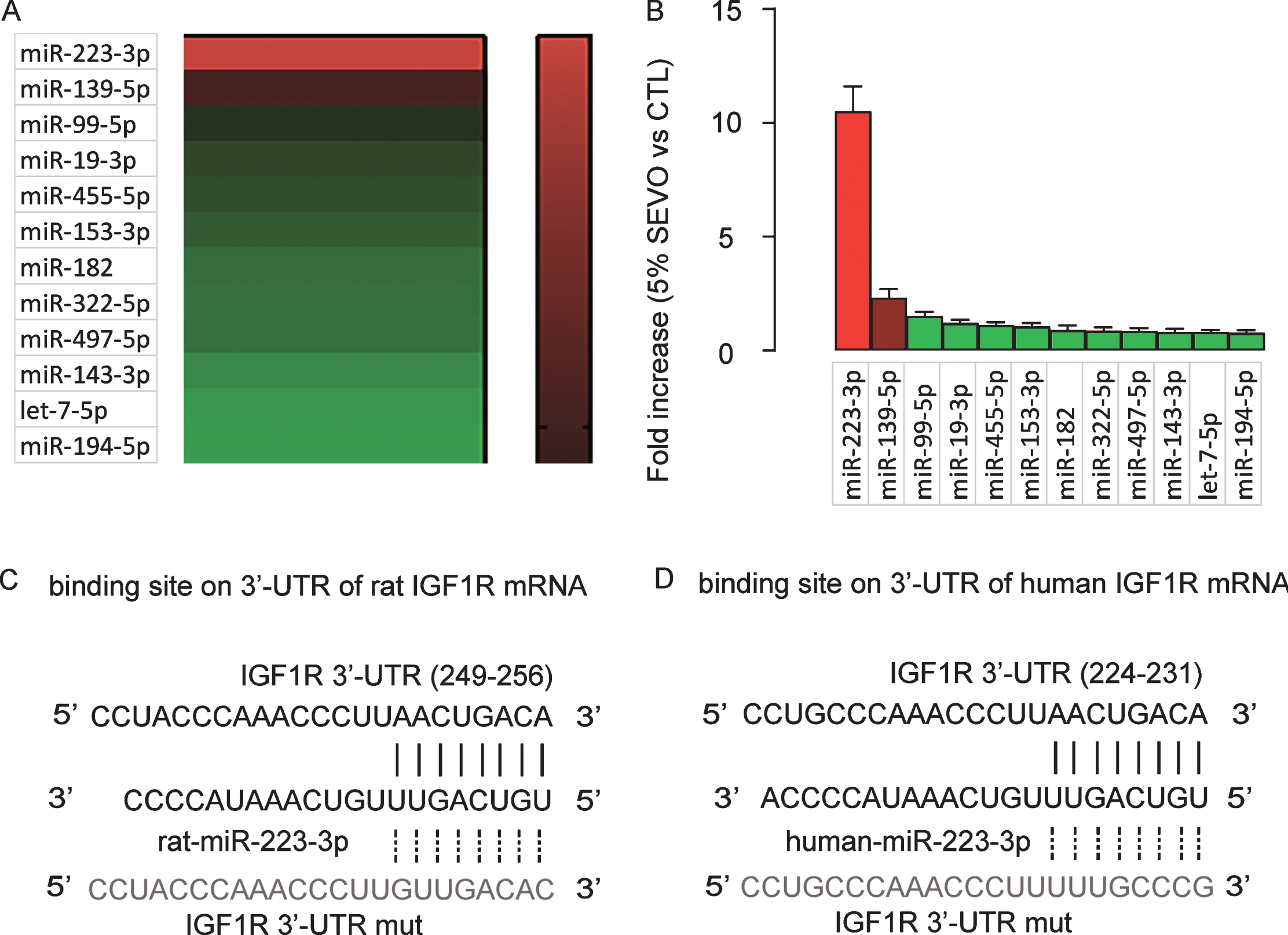
SEVO upregulates miR-223-3p in neuronal cells. A) Heat map for IGF1R-targeting miRNAs in HCN-2 cells exposed to either 5% SEVO or CTL gas in a miRNA assay. B) RT-qPCR for IGF1R-targeting miRNAs in HCN-2 cells exposed to either 5% SEVO or CTL gas. Bioinformatic analysis for predictive binding sites of miR-223-3p to 3’-UTR of IGF1R mRNA of rat (C) and human (D). N = 5.
MiR-223-3p inhibits translation of IGF1R in neuronal cells
Next, we examined how the binding of miR-223-3p to IGF1R affects the expression of IGF1R. First, miR-223-3p was either overexpressed or inhibited by plasmids carrying a miR-223-3p mimic or an as-miR-223-3p in HCN-2 cells. Transfection with a null sequence was used as a control. The RT-qPCR for miR-223-3p levels confirmed the alteration in miR-223-3p levels in these cells (Fig. 4A). Next, a wildtype 3’-UTR of IGF1R mRNA (WT-IGF1R 3’-UTR) and an 3’-UTR of IGF1R mRNA with a mutant at the miR-223-3p-binding site (MUT-IGF1R 3’-UTR) were generated to be used in a dual luciferase reporter assay together with miR-223-3p-modifying plasmids. The data showed that the specific binding of miR-223-3p to 3’-UTR of IGF1R mRNA inhibited IGF1R protein translation in HCN-2 cells (Fig. 4B). The mRNA levels of IGF1R were unchanged (Fig. 4C), but the protein levels of IGF1R significantly altered by modification of miR-223-3p levels (Fig. 4D). Together, these data suggest that miR-223-3p inhibits translation of IGF1R in neuronal cells.
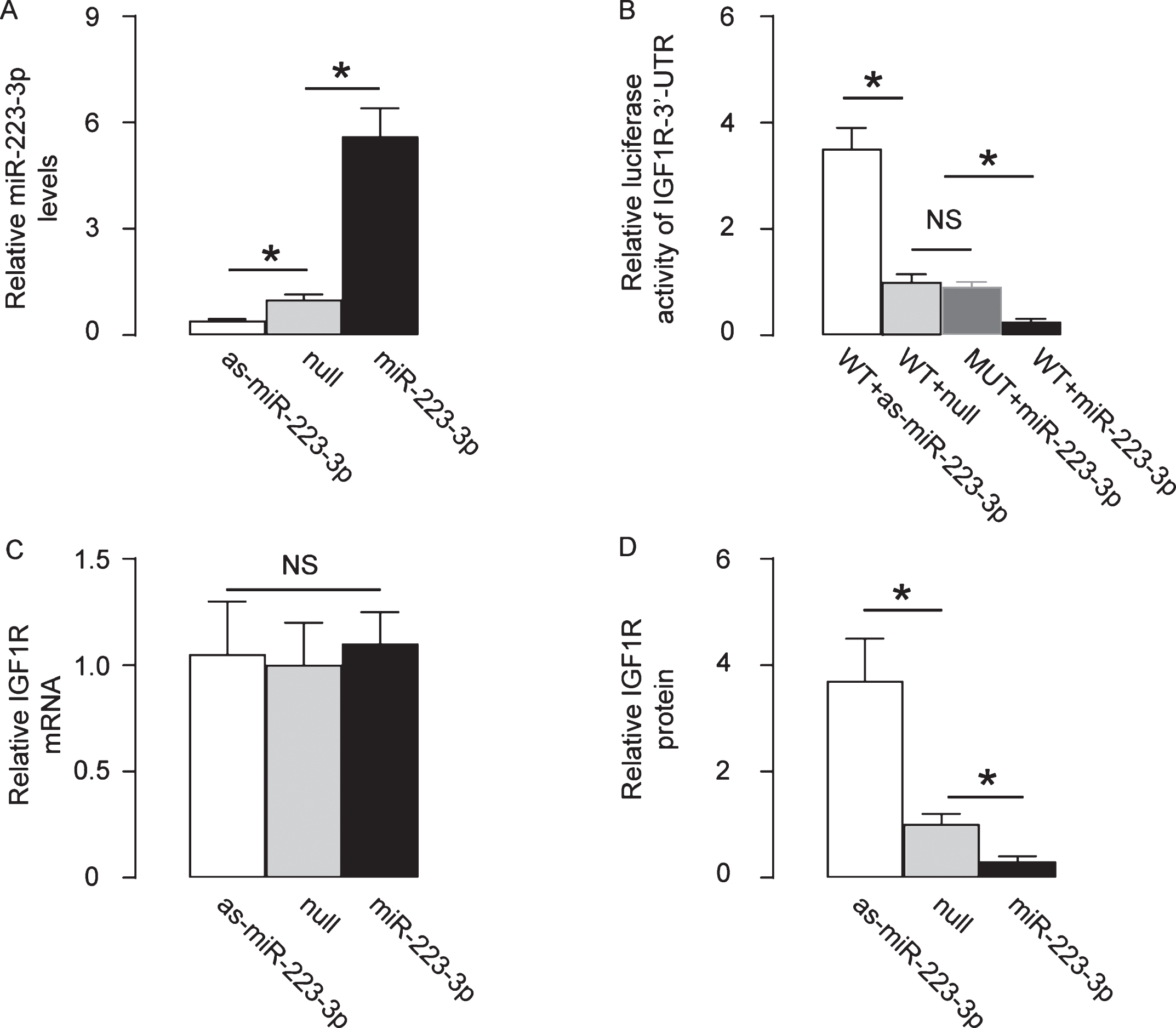
MiR-223-3p inhibits translation of IGF1R in neuronal cells. MiR-223-3p was either overexpressed or inhibited by plasmids carrying a miR-223-3p mimic or an as-miR-223-3p in HCN-2 cells. Transfection with a null sequence was used as a control. A) RT-qPCR for miR-223-3p levels. B) A wildtype 3’-UTR of IGF1R mRNA (WT-IGF1R 3’-UTR) and an 3’-UTR of IGF1R mRNA with a mutant at the miR-223-3p-binding site (MUT-IGF1R 3’-UTR) were generated to be used in a dual luciferase reporter assay together with miR-223-3p-modifying plasmids. RT-qPCR (C) and ELISA (D) for IGF1R levels in miR-223-3p-modified HCN-2 cells. *p < 0.05. NS, non-significant. N = 5.
Abolishment of miR-223-3p attenuates SEVO-induced neuronal cell apoptosis
Finally, we evaluated the effects of abolishment of miR-223-3p upregulation on neuronal cell apoptosis by SEVO in vitro. Null- or miR-223-3p- transfected HCN-2 cells were exposed to 5% SEVO and casp3 expression was examined after 48 h. Our data showed that abolishment of miR-223-3p significantly attenuated SEVO-induced neuronal cell apoptosis, shown by representative images (Fig. 5A), and by quantification (Fig. 5B). Moreover, abolishment of miR-223-3p significantly increased the total number of the cells after SEVO exposure (Fig. 5C). Finally, the IGF1R levels were determined in these cells, confirming the salvage of IGF1R (Fig. 5D).

Abolishment of miR-223-3p attenuates SEVO-induced neuronal cell apoptosis. The effects of abolishment of miR-223-3p upregulation on neuronal cell apoptosis by SEVO were examined in vitro. Null- or miR-223-3p- transfected HCN-2 cells were exposed to 5% SEVO and casp3 expression was examined after 48 h, shown by representative images (A), and by quantification (B). C) The quantification of total cell number. D) ELISA for IGF1R in treated HCN-2 cells. *p < 0.05. NS, non-significant. N = 5.
DISCUSSION
Insulin and insulin receptor play critical roles in glycose metabolism. This signaling pathway initiates from binding of insulin receptor by insulin, which induces a rapid auto-phosphorylation of insulin receptor, followed by activation of downstream signaling pathways, such as PI3K-AKT and MAPK to regulate a variety of biological events. Insulin receptor also exists in the hippocampus [18], and its compromised expression has been shown in the brains of patients with Alzheimer’s and Parkinson diseases [19].
SEVO exposure has been shown to induce neurotoxicity through IGF-1 signaling [20–22]. Interestingly, SEVO has been shown to regulate IGF-1 expression by miRNAs in liver [23], and in brain [24, 25]. However, the direct regulation of IGF1R by SEVO was not shown before. Here, by in vivo and in vitro studies, we showed that SEVO induced apoptotic death of neuronal cells, through post-transcriptional inhibition of IGF1R by miR-223-3p. The apoptosis was determined by analysis on casp3 expression and changes in cell number. Cell viability and mass maintenance may result from summation of apoptosis, and pro-survival and/or anti-survival, and proliferation of the cells in this model. Future studies may address the detailed mechanisms of cell death upon SEVO exposure. Interestingly, although both IGF-1 and IGF1R were both suppressed in the brain from the SEVO-treated rats, it seemed that the source of the changes in IGF-1 levels was non-neurons. It may be interesting to examine cells other than neurons, e.g., microglial, to determine the molecular control of IGF-1 by SEVO.
The conclusion of changes in post-transcriptional control of IGF1R was resulting from the fact that IGF1R protein was significantly reduced by SEVO, but IGF1R mRNA was unaffected. Although protein post-translational modification and degradation may also contribute to this result, our approach in a loss-of-function experiment showed that the as-miRNA-mediated abolishment of miR-223-3p expression may be more important for the differential expression mRNA and protein of IGF1R, since it nearly completely abolished the effects on cell apoptosis. The limitation of the current study is that we are not able to determine the overall effects of miR-223-3p knockdown in contradicting the detrimental effects of SEVO on rat brain, since miR-223-3p has target genes other than IGF1R. In the future, a systematic analysis on miR-223-3p target genes may provide more evidence for a miR-223-3p-based treatment against SEVO-exposure.
DISCLOSURE STATEMENT
Authors’ disclosures available online (https://www.j-alz.com/manuscript-disclosures/19-0596r1).