
Abstract
Background:
Post-stroke cognitive impairment (PSCI) is an important factor causing disabilities after acute ischemic stroke (AIS). Emerging evidence suggested that gut microbiota play an important role in cognitive impairment.
Objective:
This study aimed to explore the association between PSCI and gut microbiota.
Method:
65 patients with newly diagnostic AIS finished the fecal collection on admission and cognitive assessment 3 months later in the clinic. Fecal samples were subjected to 16SrRNA gene sequencing and gas chromatography-mass spectrometry analysis. Additionally, we enrolled new 18 AIS patients, whose treatment was supplemented by probiotics, to assess the potential of microbial treatment in PSCI.
Results:
PSCI patients were characterized by the significantly decreased alpha-diversity, disturbed microbial composition, and corresponding metabolites compared with non-PSCI patients. Increased Fusobacterium and deficiency of microbial metabolized short-chain fatty acids (SCFAs) were significantly associated with PSCI. A model based on gut microbiota and SCFAs could predict 3 months or longer PSCI early and accurately after stroke onset. While traditional probiotic administration had little effect on PSCI, it could ameliorate patients’ mood, including depression and anxiety in the 3 months after stroke.
Conclusion:
Our study revealed the association between PSCI and gut microbiota and its corresponding metabolites for the first time, suggesting the potential for applying microbiota and its corresponding metabolites to early clinical diagnosis and treatment of PSCI.
INTRODUCTION
Stroke carries the second leading cause of death and third leading cause of disability adjustment life years lost all over the world, about 80% of which are ischemic stroke [1, 2]. Post-stroke cognitive impairment (PSCI) is fairly common in acute ischemic stroke (AIS) patients and probably develops into dementia [3]. Within a year of stroke, the incidence of PSCI ranges from 7.4% to 41.3% [4]. However, it often takes 3 months or longer post-stroke to diagnose PSCI by the present standard criteria, such as the Montreal Cognitive Assessment scale (MoCA), the Diagnostic and Statistical Manual of Mental Disorders IV, and the Mini-Mental State Examination (MMSE). Besides, only a few drugs used in Alzheimer’s disease are effective for PSCI patients, such as cholinesterase inhibitors and memantine. How to distinguish PSCI early and find newly valid treatment for PSCI is gaining a substantial amount of attention [3, 5]. With the development of metagenomics and the establishment of germ-free models, researchers have discovered that many neurological diseases are closely associated with gut microbiota, which possess many more genes than the human genome [6, 7]. The “microbiota-gut-brain” axis plays an important role in the occurrence and development of stroke and its corresponding post-stroke cognitive impairment [8–10]. Rodent studies have confirmed that disturbed gut microbiota could aggravate cognitive impairment, which still needs detailed mechanism studies and human studies to validate [11–15]. Moreover, the effect of microbial intervention on cognitive function, such as probiotics, prebiotics, and synbiotics is mixed [16–20], suggesting there is still lack of persuasive cognitive studies to expound the role of gut microbiota in cognitive impairment.
Thereby, we focused on post-stroke cognitive impairment, excluding cognitive impairment caused by other diseases in this study. We tried to find the association between PSCI and gut microbiota, its corresponding metabolites, and reveal the effect of probiotics on PSCI. Moreover, we applied gut microbiota and corresponding metabolites to the early diagnosis of PSCI.
METHODS
Enrollment of participants in this study
We performed a longitudinal research study including 83 patients between June 2018 and June 2019 in Shanghai Tenth People’s Hospital, China. First, we enrolled 65 patients with newly diagnosed AIS, as confirmed by magnetic resonance imaging, who finished the fecal collection on admission and cognitive assessment 3 months later in the clinic. After we found a significant association between PSCI and gut microbiota, we enrolled 18 new AIS patients, whose treatment was supplemented by probiotics to assess the effect of probiotic administration on PSCI. All 83 enrolled patients did not have severe hearing, visual, or reading deficits and cognitive impairment before stroke, that were recorded in medical documents or reflected by patients and families. Patients were excluded due to 1) illiterate, 2) cancer, 3) exposure to antibiotics, probiotics, prebiotics, or synbiotics within 1 month before admission, 4) severe depression or anxiety, 5) cerebral hemorrhage confirmed by imaging, 6) severe intestinal diseases, such as inflammatory bowel disease. The probiotics, took by 4*10∧6CFU/d * 90d, were composed of Bifidobacterium longum, Bifidobacterium lactis, Lactobacillus acidophilus, Lactobacillus rhamnosus, Lactobacillus paracasei, and Lactobacillus fermentan. Demographic and clinical data were gathered from the electronic medical records system. All the patients were provided with informed consents. This study has been approved by the ethical review committee of the Shanghai Tenth People’s Hospital and registered in clinical trials (NCT03812445).
Fecal sample and clinical data collection
To avoid the effect of drugs and hospitalization on gut microbiota, fecal samples were collected within 36 hours of admission. Fresh fecal samples were immediately given to doctors or nurses and then gathered into the sterile tubes. These samples were stored at –80°C prior to microbial analysis. Demographic and clinical data were collected during hospitalization, including age, sex, body mass index (BMI), high density lipoprotein cholesterol (HDL-C), low density lipoprotein cholesterol (LDL-C), triglyceride, and total cholesterol. Additionally, we collected possible cognition-related indicators reported in emerging studies, including fasting glucose, homocysteine, and thyroid function: free triiodothyronine (FT3), free tetraiodothyronine (FT4), total triiodothyronine (TT3), total tetraiodothyronine (TT4), and thyroid-stimulating hormone (TSH). In addition, we used the National Institute of Health stroke scale (NIHSS) to assess the severity of stroke.
After 3 months of AIS onset, all enrolled patients were evaluated with the Beijing version of the MoCA, MMSE, Hamilton Depression Scale (HAMD), and Hamilton Anxiety Scale (HAMA) in a stable condition by senior neurologists in the outpatient department. The Beijing version of the MoCA has been widely used in China to assess cognition of stroke patients. MoCA scores range from 0 to 30. If the score was below 26, the patient was considered to have PSCI. Considering the influence of education, we added one point to the score of patients who received less than 12 years of education, following the instruction of MoCA [21].
16S High-throughput sequencing and targeted metabolomics
DNA was extracted from fecal samples using the QlAamp DNA Stool Mini kit (Qiagen). The DNA concentrations and quality were examined by a Nanodrop 2000 uv-vis spectrophotometer (Thermo Scientific, Wilmington, MA, USA) and 1% agarose gel electrophoresis. The V3–V4 hypervariable regions of the bacteria 16S rRNA gene were amplified with forward primer 314F (5′- CCTACGGGRSGCAGCAG -3′) and reverse primer 806R (5′-GGACTACVVGGGTATCTAATC -3′) on a thermocycler PCR system. PCR products were extracted from a 2% agarose gel electrophoresis and recycled by AxyPrep DNA Gel Recovery Kit (Axygen Biosciences, Union City, CA, USA), and then examined by a Nanodrop 2000 uv-vis spectrophotometer (Thermo Scientific, Wilmington, MA, USA). Purified amplicons were pooled in equimolar solutions and underwent paired-end sequencing using an IlluminaMiSeq PE250 (Illumina, San Diego, CA, USA). Targeted SCFAs metabolome was tested by 7890A-5975C gas chromatography-mass spectrometer (GC-MS) (Agilent Technologies, Palo Alto, CA, USA) [22].
Bioinformatic and statistical analysis
After quality control of the original data, we filed the raw data, identified the optimized sequences, and then used Usearch (version7.1, http://drive5.com/uparse/) to assign the raw sequences to operational taxonomic units (OTUs) based on OTUs having ≥97% similarity. Each OTU is considered to represent each taxonomic level, i.e., kingdom, phylum, class, order, family, and genus. The OTUs cluster analysis was performed following chimeric filtration, including alpha diversity, beta diversity, analysis of biological difference and relevance as well as prediction of functional composition. Alpha diversity was used to describe the abundance and diversity of gut microbiota, including Observed_species index, Chao1 index, Shannon index, and Simpson index analysis. Beta diversity was utilized for further analysis of species diversity of gut microbiota from fecal samples, including analysis of similarity (Anosim) and Principal Coordinates Analysis (PCoA). Anosim was applied to estimate whether the difference between groups was greater than that within groups, and if it made sense to divide AIS patients into the both groups. Analysis of biological difference and relevance were consisted of linear discriminant analysis coupled with effect size (LEfSe), Spearman analysis of gut microbiota, and MoCA. Subsequently, we used Phylogenetic investigation of communities by reconstruction of unobserved states (PICRUSt) to predict the microbiota-associated functional pathway based on Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway.
All quantitative data were analyzed using GraphPad prism 8.0 and SPSS 22.0 software. Clinical data were expressed as mean±standard deviation. We used the Student’s t, Pearson’s Chi-square, Fisher’s exact and Mann-Whitney U tests to analyze the continuous and categorical variables, respectively.
RESULTS
General descriptions of enrolled patients
The demographic and clinical data of 65 AIS patients mentioned above are listed in Table 1. As shown, there were no difference found in sex, age, and BMI. Moreover, we found no difference in some clinical indicators that may influence cognition of AIS patients, including fasting glucose, homocysteine, thyroid function, and severity of AIS (assessed by NIHSS), suggesting none of confounding clinical parameters influenced group discrimination. For further study, we compared the sub item of MoCA between the both groups (shown in Table 2), demonstrating that PSCI patients had significantly lower scores, especially in visuospatial/executive function, language, abstraction, and delayed recall compared with non-PSCI patients.
Demographic and clinical characteristics of PSCI subjects versus non-PSCI subjects
Data were expressed as mean±standard deviation. BMI, body mass index; TC, total cholesterol; TG, triglyceride; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; FT3, free triiodothyronine; FT4, free tetraiodothyronine; TT3, total triiodothyronine; TT4, total tetraiodothyronine; TSH, thyroid-stimulating hormone; NIHSS, National Institutes of Health stroke scale.
Sub item of MoCA scores of patients with or without PSCI
*p < 0.01, **p < 0.001.
Lower diversity and disturbed composition of gut microbiota in PSCI subjects
From all fecal samples of enrolled patients, a total number of 3,196,723 clean reads were obtained, with an average count of 49,180 clean reads per sample, which were clustered into 696 OTUs based on ≥97% similarity. The Venn diagram suggested that 533 OTUs were shared by both groups while 113 and 48 OTUs were unique to non-PSCI and PSCI subjects, respectively (Supplementary Figure 1). Alpha diversity was utilized to assess difference in the richness (Chao1 and observed species indices) and diversity (Shannon and Simpson indices) of gut microbiota. Compared with non-PSCI subjects, PSCI subjects had significantly lower Shannon index (p = 0.027), observed species (p < 0.01), and Chao1 index (p < 0.01), whereas no significant difference was observed in Simpson index (p = 0.17) between the both groups (Fig. 1A).
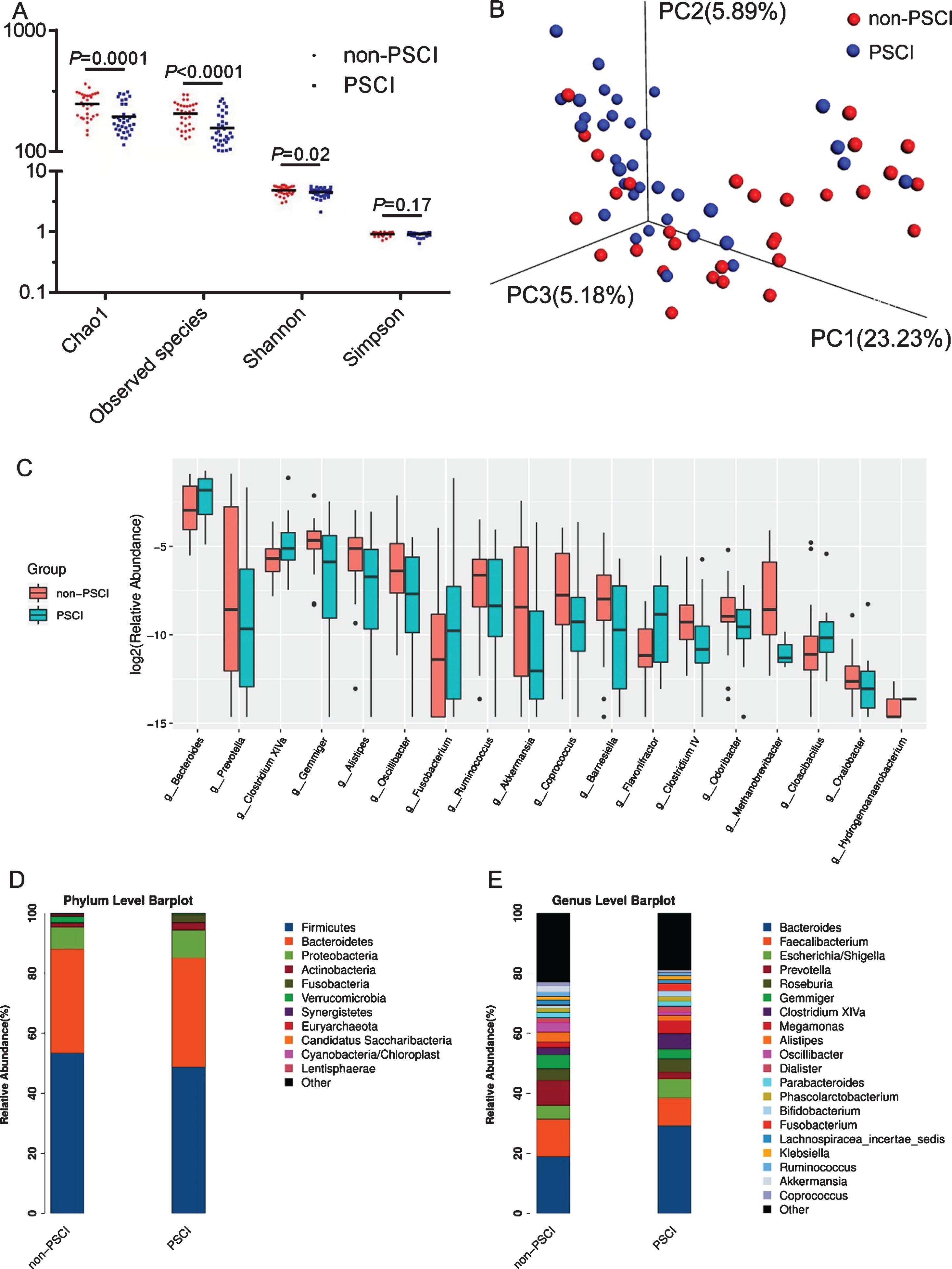
Diversity and structure of gut microbiota in AIS patients with or without PSCI. A) Significantly distinct alpha diversity between the both groups. B) Unweighted PCoA was used to illustrate bacterial 3D clustering. C) The different relative abundance of genera identified by Wilcoxon rank-sum test. Component proportion of bacteria in phylum (D) and genus (E) level between both groups was showed.
Moreover, the three-dimensional principal coordinate analysis based on unweighted UniFrac distance demonstrated that significantly different composition of gut microbiota was found between the both groups (Fig. 1B). For further verification, unweighted UniFrac Anosim was performed (R = 0.228, p = 0.001) (Supplementary Figure 2), confirming the distinct bacterial structure between the two groups.
Significantly different abundance of microbiota in PSCI versus non-PSCI subjects
Linear discriminant analysis effect size analysis and Wilcoxon rank-sum test were utilized to identify the significantly different bacterial taxa between the both groups. At the level of phylum, Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria were the main phyla in the both groups (53.37% versus 48.75%; 34.67% versus 36.31%; 7.24% versus 9.29%, and 1.24% versus 4.27%, respectively). There were 4 taxa found to be distinct between the both groups, including Fusobacteria, Synergistetes, Verrucomicrobia, and Euryarchaeota (all p < 0.05). The relative abundance of Fusobacteria was significantly increased in PSCI group, whereas that of the remaining 3 taxa were decreased in PSCI group (Fig. 1D). At the genus level, 20 taxa were identified for discriminating PSCI subjects from non-PSCI subjects (LDA score>2, p < 0.05), including enriched abundance of Fusobacterium, Bacteroides, Clostridium XlVa, Gemella, and Flavonifractor, and decreased abundance of the remaining 15 genera (Fig. 2). Generally, the 20 distinct genera mainly belonged to the phyla Firmicutes (9/20, 45%) and Bacteroidetes (5/20, 25%). Notably, the relative abundance of Fusobacterium, which increased approximately 10-fold in PSCI relative to that in non-PSCI (0.26% versus 2.51%, p < 0.01), demonstrated the strongest relation with PSCI, whereas the relative abundance of SCFAs-producing genera decreased significantly in PSCI group compared to that in non-PSCI group, such as Oscillibacter, Ruminococcus, Gemmiger, and Coprococcus (Fig. 1C, E).
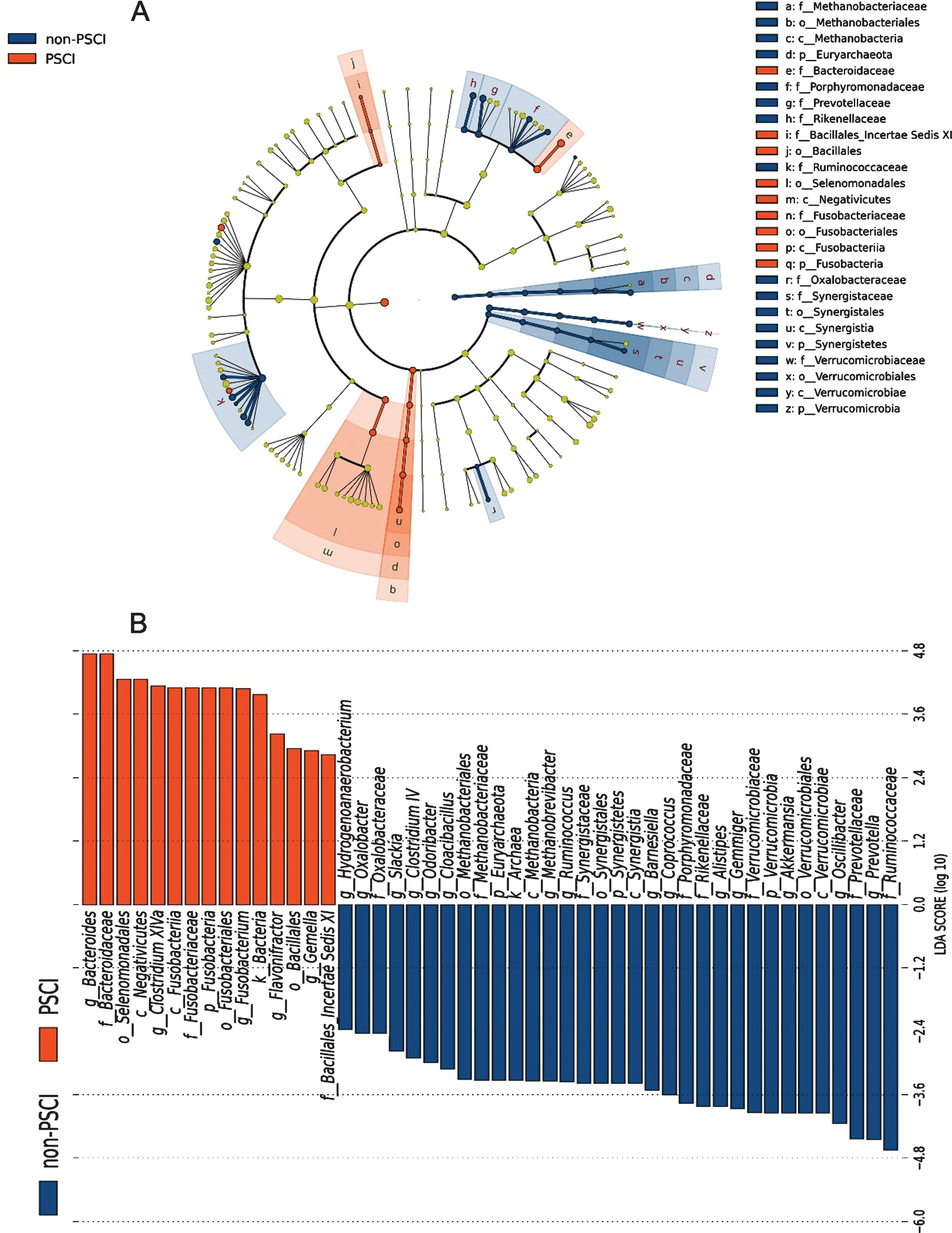
Linear discriminant analysis integrated with effect size (LEfSe). Cladogram (A) and histogram (B) illustrated the phylogenetic distribution of gut microbiota related to both groups.
Altered short-chain fatty acids in PSCI subjects
Based on LEfSe, we found that numerous genera, which were capable to produce SCFAs by fermentation of dietary fiber, decreased significantly in PSCI subjects. Emerging evidence has confirmed SCFAs play an important role in brain health, such as affection and cognition [23]. In this study, 7 dominant kinds of SCFAs were tested by targeted metabolomics, including acetic acid, propionic acid, isobutyric acid, butanoic acid, isovaleric acid, valeric acid, and hexanoic acid. Notably, Mann-Whitney U test identified the significantly decreased content of SCFAs in PSCI group, especially hexanoic acid (Fig. 3A–G). Correlation analysis showed that SCFAs were positively associated with SCFAs-producing bacteria, suggesting lack of SCFAs is probably due to decreased abundance of SCFAs-producing bacteria in AIS patients (Fig. 3H).
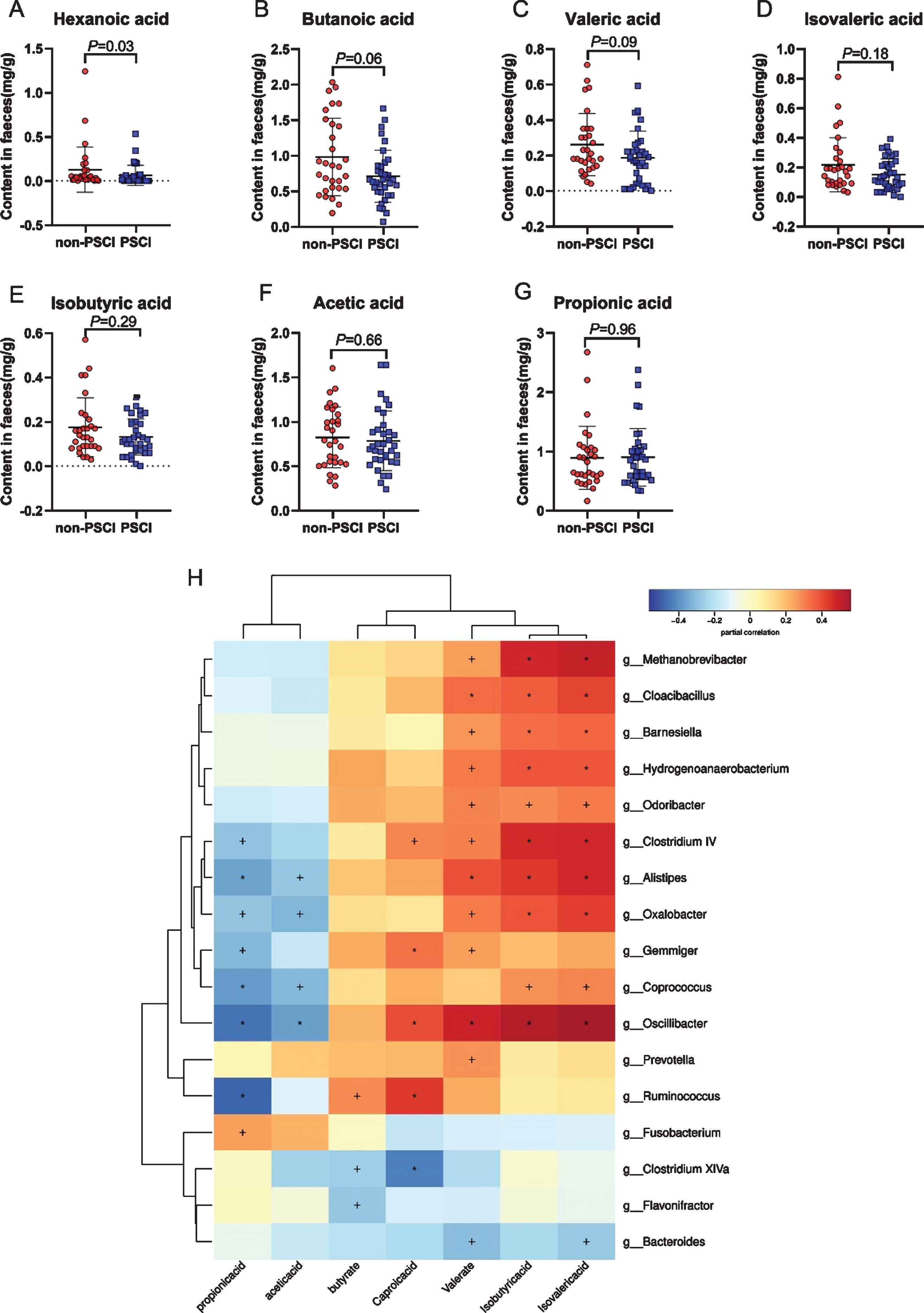
Comparison of SCFAs between AIS patients with or without PSCI. 7 main kinds of SCFAs were denoted in the form of boxplot (A–G). H, the heat map illustrated the correlation analysis of SCFAs and gut microbiota. +p < 0.05; *p < 0.01.
Gut microbiota and SCFAs-based model discriminated PSCI
To assess the potential capability of gut microbiota and SCFAs as biomarkers for early diagnosis of PSCI, receiver operating characteristic curve (ROC) was performed. Utilizing different 20 genera found in LEfSe generated an area under curve (AUC) of 0.836. Further, we found the predictive performance could be improved significantly with an AUC of 0.934 by the most distinct 10 genera combined with hexanoic acid, which primarily belonged to Fusobacteria, Firmicutes, and Bacteroidetes (Fig. 4).
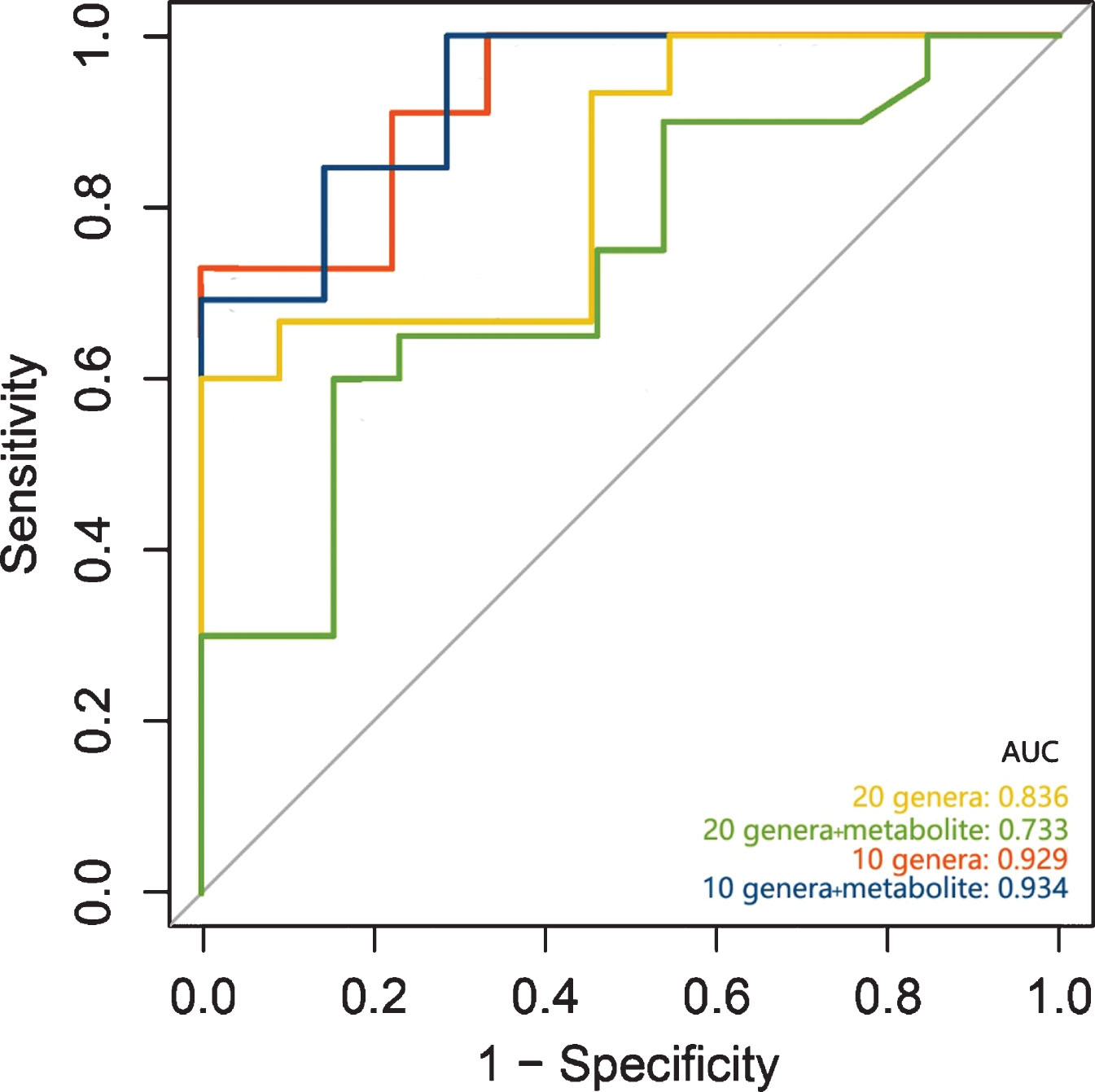
Predictive models based on microbiota and metabolites for early diagnosis of PSCI.
16S functional prediction and correlation analysis
Based on the Kyoto Encyclopedia of Genes and Genomes databases, we used PICRUSt to predict the function of gut microbiota. The significantly changed functions of gut microbiota were the upregulation of carbohydrate metabolism and immune system diseases in PSCI group. Remarkably, we identified the overexpression of utilization of glucose in L3 category in PSCI group, including fructose and mannose metabolism, galactose metabolism, and pentose phosphate pathway. In addition, amino acid metabolism and other iron coupled transporters pathway were also found to be significantly abundance in PSCI group (Supplementary Figure 3). The microbiota-microbiota correlation analysis of all AIS patients demonstrated that Fusobacterium, Flavonifractor, Bacteroides, and Clostridium XlVa were positively correlated with each other, whereas they were negatively associated with some SCFAs-producing or common bacteria, such as: Gemmiger, Barnesiella, Coprococcus, and Ruminococcus (Supplementary Figure 4). To determine the association between gut microbiota and clinical parameters of AIS patients, we performed correlation analysis of MoCA, clinical variables, and gut microbiota. Notably, Fusobacterium had the strongest negative correlation with MoCA score, whereas SCFAs-producing bacteria were positively related to MoCA score. Additionally, Fusobacterium was positively related to TG, which might be a potential pathway that Fusobacterium influences PSCI. Additionally, homocysteine, a cognition-related clinical variable, was positively related to Alistipes (Fig. 5).
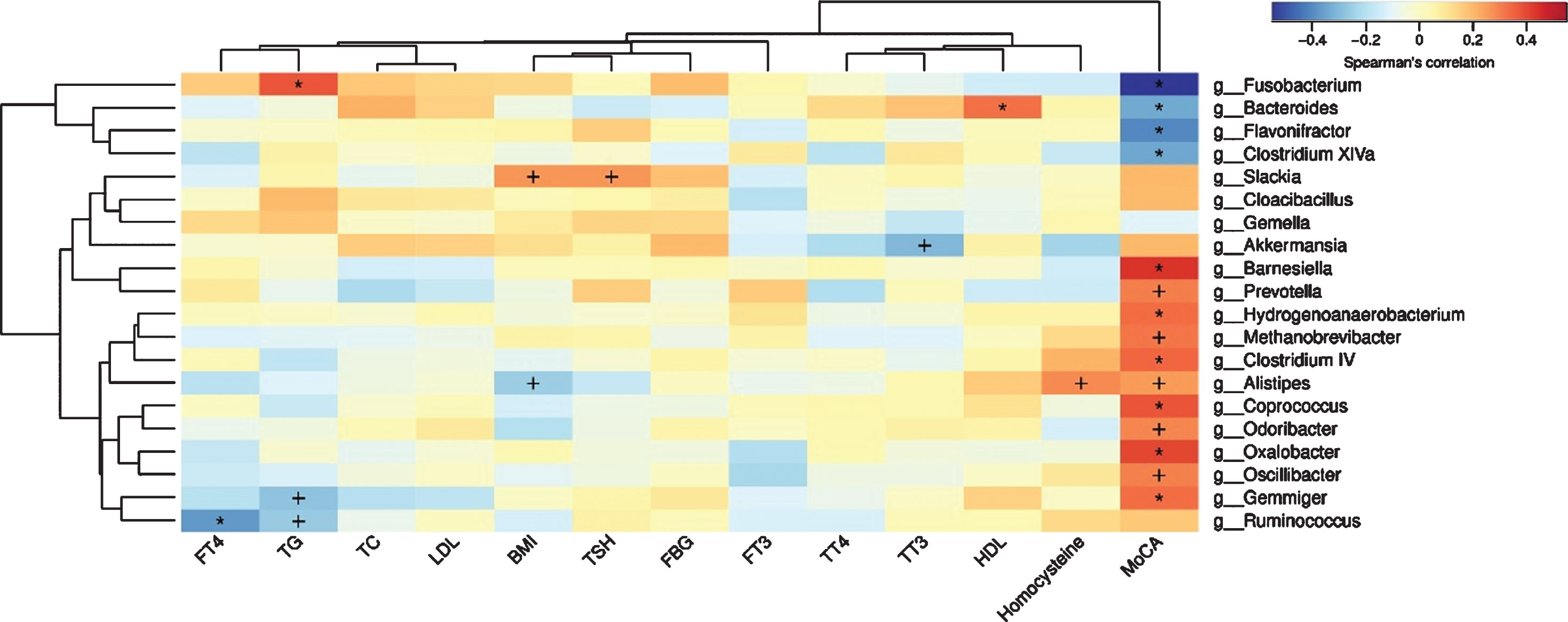
Spearman’s correlation analysis of gut microbiota, MoCA as well as clinical variables. +p < 0.05; *p < 0.01.
The effect of probiotics intervention on PSCI
Next, we recruited 18 newly diagnosed AIS patients, whose treatment was supplemented by probiotics. The assessment 3 months later showed that compared to the 65 subjects without probiotic treatment mentioned above, the 18 probiotics-treated subjects had better mood, including depression and anxiety (assessed by HAMD and HAMA), whereas no statistical difference was found in the proportion of PSCI and average scores between both groups (assessed by MoCA and MMSE) (Table 3).
MoCA scores in probiotics-treated subjects versus controls
DISCUSSION
After the proposal of the “brain-gut-microbiota axis”, microbiota-associated studies have paid great attention to the alteration of composition, function, and metabolome of gut microbiota in diseases of the central nervous system. Accumulating evidence has confirmed that gut microbiota and its metabolome were associated with cognitive function [24]. Therefore, utilizing gut microbiota and its metabolites as new screening tools for early recognition of PSCI is promising. We revealed for the first time the hallmark of gut microbiota and its metabolites SCFAs in AIS patients with PSCI in this study, following 16S sequencing and targeted metabolome analysis. We also established a gut microbiota and SCFAs-based model that had the power for early diagnosis of PSCI. Moreover, we found that traditional probiotics intervention could improve AIS patients’ mood, but it had no effect on AIS patients’ cognition.
It was exhibited that PSCI subjects had significantly decreased diversity of gut microbiota relative to subjects without PSCI, suggesting a high microbial diversity might reflect the relatively healthy status after acute ischemic stroke. Similar decrease of microbial diversity was also observed in other cognitive impairment-related diseases from humans and rodents, including Alzheimer’s disease and streptozotocin-induced diabetes [25–27].
Consistent with the decreased diversity, we found the microbial composition of PSCI subjects was significantly different from that of non-PSCI subjects. Based on LEfSe and correlation analysis, it was found that Fusobacterium had the strongest correlation with PSCI. Fusobacterium is a gram-negative non-sporeforming pathogen, which could induce the dysbiosis of intestinal microenvironment, generate proinflammatory environment in vivo, and cause immune evasion of some pathogenic bacteria [28, 29]. Besides, our former studies discovered that Fusobacterium nucleatum enhanced intestinal tumor cell invasion and adhesion by activating Toll-like receptor and upregulating expression of microRNA-21 [30]. Hence, we speculated that Fusobacterium could aggravate cognitive impairment by promoting the inflammatory response of the neural system and activating specific receptors after stroke, which still needs future study to verify. Remarkably, emerging evidence demonstrated that Fusobacterium was enriched in some neural diseases, including autism spectrum disorder [31], mild cognitive impairment [32], and myasthenia gravis [33], hinting that Fusobacterium and its metabolites might be able to injure the neural system via direct or indirect involvement.
Moreover, the bacterial composition of PSCI subjects were also characterized by the lack of SCFAs-producing bacteria, such as Oscillibacter, Ruminococcus, Gemmiger, Coprococcus, and Barnesiella. SCFAs, the major metabolites from bacterial fermentation of dietary fiber, played an important role in the nervous system [34]. It was confirmed that SCFAs could cross the blood-brain barrier to the brain and their receptors were expressed in brain tissue and sympathetic ganglia [35]. Thereby, we tested the content of SCFAs from AIS patients’ feces to explore the relationship between SCFAs and complete cognitive function. Consistent with the decreased SCFAs-producing bacteria, the content of main SCFAs in feces of PSCI subjects decreased significantly compared with that of non-PSCI subjects. Not only limited to PSCI, the decreased SCFAs were also found in Alzheimer’s disease and Parkinson’s disease [36, 37]. SCFAs play a critical role in microbiota-gut-brain axis by interacting with free fatty acid receptor, inhibiting histone deacetylases, and affecting microglia cell [23, 39]. It has also been confirmed that SCFAs administration could ameliorate cognitive function in rodent researches and inhibit Aβ aggregation in vitro. Taken together, the significantly increased Fusobacterium and decreased SCFAs induced severe immune disorders, which might be capable in explaining the occurrence and development of PSCI.
The probiotic intervention improved mood after stroke, including anxiety and depression, but it had no statistically significant effect on cognition. The probiotics used in this study were composed of Lactobacillus and Bifidobacterium, which were capable to produce SCFAs. Lactobacillus could produce SCFAs by fermentation of carbohydrates and by the phosphoketolase route in the hetero fermenting conditions [42]. Bifidobacterium uses the fermentation pathway to produce mainly acetate and formate under carbohydrates limitation, and acetate and lactate when carbohydrates are in excess [43]. Supplementation of Lactobacillus and Bifidobacterium increased the concentration of SCFAs in SHIME (for Simulator of Human Microbial Ecosystem) reactor [44]. Based on the research mentioned above, we tried to improve microbial structure and alleviate the lack of SCFAs in PSCI patients utilizing Lactobacillus and Bifidobacterium. The useful intervention for improving PSCI might need high-dose supplement of SCFAs or SCFAs-producing probiotics rather than traditional-dose Lactobacillus and Bifidobacterium. Additionally, fecal bacteria transplantation or supplement of new-type probiotics, such as Akkermansia, which decreased significantly in PSCI subjects might have potential for ameliorating PSCI.
Nonetheless, there were still some shortcomings. First, due to the simplicity of the research design, we only excluded patients with cognitive impairment before stroke and performed cognitive assessment 3 months after stroke. In future research, we will further explore the association between PSCI and microbiota, as well as its metabolized SCFAs, containing detailed and long-term follow-up of cognitive function. Secondly, we failed to collect fecal samples 3 months after stroke that should have been utilized to assess microbial structure and measure SCFAs concentration after taking probiotics.
The gut microbial signature and corresponding metabolites, as well as the diagnostic model associated with PSCI, still need a larger sample size of clinical studies to validate. We used 16S rRNA rather than metagenomic sequencing, which limited us to the finding of specific bacteria related to PSCI in the species level.
In brief, we found that PSCI was characterized by the anomalous gut microbial diversity, composition, and corresponding metabolites. PSCI subjects could be distinguished from non-PSCI subjects accurately, utilizing specific bacteria and metabolites. Traditional probiotics administration had beneficial effect on the control of depression and anxiety after stroke.
Footnotes
ACKNOWLEDGMENTS
All authors are thankful for the support of all enrolled patients. This study is supported by the National Natural Science Foundation of China (81230057, 81372615, 81472262 and 81200264) and Clinical Research Project of Xiuyisheng Probiotics and Chronic Disease.