
Abstract
The gut microbiota is made up of trillions of microbial cells including bacteria, viruses, fungi, and other microbial bodies and is greatly involved in the maintenance of proper health of the host body. In particular, the gut microbiota has been shown to not only be involved in brain development but also in the modulation of behavior, neuropsychiatric disorders, and neurodegenerative diseases including Alzheimer’s disease. The precise mechanism by which the gut microbiota can affect the development of Alzheimer’s disease is unknown, but the gut microbiota is thought to communicate with the brain directly via the vagus nerve or indirectly through signaling molecules such as cytokines, neuroendocrine hormones, bacterial components, neuroactive molecules, or microbial metabolites such as short-chain fatty acids. In particular, interventions such as probiotic supplementation, fecal microbiota transfer, and supplementation with microbial metabolites have been used not only to study the effects that the gut microbiota has on behavior and cognitive function, but also as potential therapeutics for Alzheimer’s disease. A few of these interventions, such as probiotics, are promising candidates for the improvement of cognition in Alzheimer ’s disease and are the focus of this review.
INTRODUCTION
Alzheimer’s disease (AD) is a multifactorial, progressive, neurodegenerative disease and the most common form of dementia. Dementia is a general term for the mental decline that effects a person’s ability to make decisions, think clearly, impairs memory, and causes changes in mood; AD is distinguished from other forms of dementia by the accumulation of extracellular amyloid-β (Aβ) fibers and intracellular neurofibrillary tangles (NFTs) which are thought to contribute to neuronal death and eventual symptom development. The strongest risk factor is advanced age as most cases are diagnosed in people over the age of 65; however, it is also known that genetic factors, i.e., apolipoprotein E (APOE), and environmental factors, i.e., air pollution exposure, contribute to disease [1–3]. Currently, the mechanism by which Aβ and NFT accumulation initially occurs is unknown. The current prevailing hypothesis is that lifelong exposures in a genetically susceptible person contribute to this phenomenon.
Recent evidence has shed light on a new potential contributor to AD development and progression: microbes (reviewed in [4, 5]). Spirochetes and the oral pathogen Porphyromonas ginigvalis have been found to be associated with Aβ plaques in postmortem brains of AD patients and AD animal models [6–10]. In vitro and in vivo evidence have both demonstrated that Aβ can act as an antimicrobial peptide, which has led to the hypothesis that Aβ accumulates in the brain in an effort to clear microbes that have penetrated the blood-brain barrier and entered the brain [11, 12]. The Gram negative bacterial membrane protein lipopolysaccharide (LPS) has also been found to be more prevalent in AD patient brains as compared to non-AD brains and can also trigger Aβ accumulation [13–15]. In examining other organ systems with bacterial alterations, AD patients have an altered gastrointestinal (GI) microbiota, termed dysbiosis, as compared to non-AD patients [16–18]. Transgenic animal models of AD also experience GI dysbiosis and AD mouse models that lack a gut microbiota have reduced amyloid plaques [19–21]. Furthermore, AD mouse models treated with antibiotics have reduced AD characteristics such as amyloid plaques and cognitive dysfunction related to memory [20, 22–24]. These studies strengthen the proposed relationship between the gut microbiota and the brain, specifically in regards to AD. Additionally, bacterial and amyloid proteins are thought to cross-seed resulting in heterogeneous amyloid formations. By acting as prions and self-seeding, infectious amyloids essentially spread their infectivity by eliciting additional amyloid conformations among other molecules. The eventual aggregation of these misfolded proteins results in neurodegeneration. Curli proteins from various bacterial species are an example of a microbial-derived protein with the ability to cross-seed amyloid formation [25]. Current understanding of prion cross-seeding, microbial amyloids, and their role in AD specifically is limited, but this is an area of growing interest due to the possibility of gut microbial molecules to perpetrate amyloid formation [26]. In this review, we will highlight the current state of the science regarding resident microbes and cognition in AD patients.
COGNITION IN AD
Cognitive health is a spectrum, and although some individuals maintain cognitive acuity with advanced age, cognitive decline is considered a common sign of aging [27]. Mild cognitive impairment (MCI) is a prodrome stage of AD, falling between normal cognitive function based on age and education-level and severe impairment due to AD dementia. Diagnostic criteria for MCI include subjective cognitive complaint, objective cognitive impairment, preserved independence in functional abilities, and no dementia. Although MCI does not always progress to AD, the cognitive difficulties commonly experienced by those suffering from MCI provide a preview of what is to come with an AD diagnosis [28]. The Mini-Mental State Examination (MMSE) is commonly implemented to assess cognitive function by testing orientation, attention, memory, language, and visual-spatial skills [29]. Typically, cognitive testing is used in conjunction with patient and family interviews and laboratory and brain-imaging tests to make an AD diagnosis [29]. Similar methods will be discussed throughout this review as varied degrees of cognitive function are explored.
Cerebral inflammation develops in AD patients years before disease onset is clinically apparent. During this asymptomatic time, Aβ plaques and activated microglial cells accumulate and contribute to the initial inflammation [30]. At first, neurological accumulation of Aβ proteins and NFTs are asymptomatic, but they later become critical hallmarks of AD brains. Aβ and NFTs in the frontal cortex and hippocampal regions of the brain decrease active neurons, resulting in a loss of function, leading to the cognitive impairment and memory deficits observed in AD patients. It is known that aberrant Aβ and NFTs are the underlying causes of AD, but why these proteins accumulate has yet to be established.
MICROBIOTA-GUT-BRAIN AXIS
There is a wealth of literature characterizing the bidirectional communication of the gut and the brain through various mechanisms [31]. The main communication pathways between the gut and the brain include: direct communication via the vagus nerve and indirect communication through the secretion of cytokines and neuroendocrine hormones to activate the immune and endocrine system [31]. Microbes that reside in the gut can also play a major role in the communication between the gut and brain, leading to the newly refined microbiota-gut-brain axis. These bacteria are able to produce substances that directly act on the enteric nervous system and vagus nerve, as well as substances that can contribute to indirect communication via activation of the immune system. Signaling molecules that are derived from or influenced by the gut microbiota include but are not limited to bacterial structural components, short-chain fatty acids (SCFAs), and neuroactive molecules. It is critical that we work to understand these signaling molecules and their relationship to cognition and AD in the presence of gut dysbiosis.
Gut dysbiosis refers to alterations of the resident bacterial populations of the gut, which can result from dietary changes, stress, medications, and alcohol or chemical consumption. Unbalanced bacterial populations may resolve themselves; however, persistent stressors can cause chronic dysbiosis, which is implicated in the development of inflammatory bowel disease, autism spectrum disorders, obesity, diabetes, cancer, and AD representing its vast pathophysiological outcomes [32–34]. The main consequences of gut dysbiosis are the production of noxious substances (e.g., lipopolysaccharide and trimethylamine) leading to an inflammatory response and an altered secretion of signaling molecules such as SCFAs [33, 36]. Uncontrolled inflammation may induce aberrant immune activation, tissue destruction, and eventual loss of function. It can also cause malfunction of cell barriers. By increasing the permeability of the gastrointestinal epithelial layers, inflammation can cause dysregulation of local epithelial layers and allow microbial byproducts to escape the gut and enter the bloodstream. Furthermore, altered levels of signaling molecules such as SCFAs are problematic as they play important roles in the host [37]. SCFAs can induce gluconeogenesis, reduce inflammation, and directly interact with vagal afferents [38–40]. Additionally, circulating levels of SCFAs can cross the blood-brain barrier (BBB), with measurable amounts in the human brain [41, 42].
As gut dysbiosis can cause increased gut per-meability and potentially increase the BBB permeability via altered SCFA secretion, systemic inflammation, and secretion of noxious compounds, this could lead to the development of cognitive impairment [31]. Systemic inflammation has been associated with reduced cognitive function specifically with short term memory and verbal learning and memory [43]. Researchers studying the relationship between brain amyloidosis and gut microbiota taxa found cognitively impaired patients with amyloidosis expressed greater levels of pro-inflammatory cytokines in blood compared to cognitively impaired patients without amyloidosis and healthy controls. Cognitively impaired patients with amyloidosis expe-rienced varied taxa abundances compared to those without amyloidosis and controls; differences were determined via stool sampling and included lower abundance of Eubacterium rectale and higher abundances of Escherichia and Shigella. The enhanced pro-inflammatory and reduced anti-inflammatory taxon abundances associated with cognitively impa-ired patients suffering from amyloidosis supports the association between the gut-microbiota, inflammation, and brain amyloidosis [44]. Interestingly, a study of gut microbiota alterations experienced by AD versus MCI patients found no genera differences between AD and MCI patients [45]. However, 11 genera from feces and 11 from blood differed between AD patients and normal controls. These findings suggest that patients suffering from AD and MCI share similar gut microbiota profiles, indicating gut dysbiosis, may originate years before AD onset [45]. This research has the potential to have diagnostic implications, as differences in microbiota profiles could provide biomarker targets for earlier and noninvasive detection of AD [45]. Systemic inflammation can also lead to an activation of the brain-resident macrophages also known as microglia cells, which have been associated with the development of AD. Microglial cells in AD are thought to play a role in synapse loss, worsening tau pathology, and release of pro-inflammatory cytokines which can further damage neurons [46]. This can lead to reduced or altered cognitive function. Furthermore, as SCFAs have been associated with increased brain derived neurotrophic factor (BDNF) expression, a critical gene for neurogenesis and synaptic plasticity, and can also act to decrease inflammation, altered SCFA levels caused by gut dysbiosis could exacerbate the damage caused by systemic inflammation in the brain [38, 48].
The various communication pathways between the gut microbiota and CNS demonstrate the complexities of their interrelationship, while also offering promising avenues for further investigation. Potential therapeutics to improve cognitive function in AD by targeting the gut microbiota have been studied using the following methods: germ-free mice, fecal microbiota transfer, humanized gnotobiotic mice, and probiotic supplementation.
GERM-FREE MICE
In order to understand the effects of the gut microbiota on the brain, several studies utilize mice free of bacteria, known as germ free (GF) mice [49]. GF mice and specific pathogen free mice (SPF), mice free of specific pathogens also sometimes referred to as conventional mice, allow researchers to determine the influence of the gut microbiota on behavior and the brain. GF mice display lower anxiety-like behavior measured by more time spent in light versus dark boxes and more time spent in the open arms of the elevated plus mazes compared to SPF mice [50, 51]. Similarly, in open field tests, GF mice spent more time than SPF mice in the center of the field instead of the periphery, further indicating reduced anxiety-like behavior among GF animals [52]. Interestingly, the introduction of microbiota from SPF mice thirty days before mating normalized some behaviors of the progeny as compared to SPF mice; however, this was not the case in adult GF mice introduced to microbiota from SPF mice [50]. Depending on the mouse strain, the differences in anxiety-like behaviors between GF and SPF mice do not always hold true. For example, C57BL/6J mice that are SPF spent more time in the center of the open field test than GF at 4 weeks of age, but this was not the case at 12 weeks. Furthermore, in C57BL/6J mice there were no differences detected in the time spent in the open and closed arms of the elevated plus maze between GF and SPF mice at 4 or 12 weeks [53]. In regards to memory, the C57BL/6J SPF mice demonstrated greater spatial and learning memory than GF mice as measured by a Morris water maze (MWM) test and had better contextual memory as measured by fear conditioning [53]. The behavioral differences between GF and SPF mice are also accompanied by neurochemical and structural differences in the brain. For example, BDNF, which is neuroprotective and promotes synaptic growth, is reduced in the amygdala and hippocampus of GF mice as compared to SPF mice [48, 55]. GF mice also display decreased brain volume and myelination as compared to SPF mice, indicating less brain maturation [53].
GF mice display higher BBB permeability as compared to SPF mice [56]. However, colonization of the gut of GF mice with a single strain of bacteria, Clostridium tyrobutyricum, whose main metabolite is butyrate, or Bacteroides thetaiotaomicron, whose main metabolites are acetate and propionate, improved BBB permeability. When treated specifically with the SCFA sodium butyrate, the GF mice displayed decreased BBB permeability and increased protein levels of occludin-5 in the frontal cortex and hippocampus [56].
The use of GF mice to study the relationship between the gut microbiota and the brain is popular, though this method comes with challenges, as maintaining GF mice is a labor-intensive and costly process requiring specialized equipment and well-trained personnel [57]. Therefore, many researchers focus on probiotic interventions or fecal microbiota transfer (FMT) to better understand the gut microbiota-brain interaction and potential therapeutics for neuropsychiatric and neurodegenerative disorders, including AD.
FECAL MICROBIOTA TRANSFER AND HUMANIZED GNOTOBIOTIC MICE
FMT is the transfer of fecal bacteria from one donor to another. In animal models, the FMT recipient is typically a GF mouse. This allows researchers to determine the effect that the gut microbiota can have on the host. In humans, FMT is frequently used to treat Clostridium difficile (CDI), which causes diarrhea and intestinal inflammation, as FMT is thought to help re-establish a healthy gut microbiota [58]. FMT is receiving further consideration as a treatment option for diseases such as irritable bowel syndrome, autism, psychiatric disorders, and Parkinson’s disease, though reports of success vary [59–61]. Regarding Parkinson’s disease specifically, a recent animal study demonstrated that a FMT from control mice to an induced Parkinson’s disease mouse model improved bradykinesia and reflexes in the FMT-recipient mice. Conversely, control mice treated with a FMT from the Parkinson’s disease mouse model displayed reduced movement and reflexes accompanied by lower levels of dopamine and serotonin in the striatum [61]. In the context of AD, a recent case study showed that an elderly man with AD had improved cognitive function when treated with a FMT for CDI at 2 and 6 months post treatment. [62]. While this is a single case study, it displays great promise for the future treatment of AD with FMT.
Another relevant animal model is the humanized gnotobiotic mouse model which involves a FMT from humans to GF mice. Post-transplant, the mice incorporate the human microbiota into their systems, providing a powerful tool for researchers to study the human gut microbiota and various diet and therapeutic interventions [63]. The humanized gnotobiotic mouse model has been studied since the 1980s, but additional research must be conducted, as behavioral and cognitive function assessments with this animal model have only recently commenced [63, 64]. Specifically, in 2016, FMT recipient GF Kunming mice from patients with major depressive disorder displayed increased depressive-like behaviors as measured with a forced swim test as compared to FMT recipient GF Kunming from healthy control volunteers [65]. More recently, FMT recipient C57BL/6J mice from patients with schizophrenia displayed impaired memory and recognition memory as measured by a Barnes maze and novel object recognition test. However, these were not GF mice but were treated with antibiotics, which depleted most of their gut microbiota [66].
PROBIOTICS
Probiotics are defined as “live microorganisms that confer a health benefit to the host when consumed in adequate amounts” [67]. Probiotic intervention involves the administration of live mono or mixed bacteria cultures to humans or animals, which can alter the host gut microbiota and/or produce substances that benefit the host. Several bacterial genera have been used as probiotics, including, but not limited to, Lactobacillus, Bacteroides, Bifidobacterium, Bacillus, and Streptococcus [68]. Such genera have been used extensively to treat various gastrointestinal conditions, such as diarrhea, constipation, and irritable bowel syndrome, supporting the notion that probiotic consumption is associated with improved immune health, bowel disease prevention, and anti-hypertensive effects [69]. Additional studies have found treatment of fibromyalgia patients with multispecies probiotics targeting gut microbial imbalances improved cognitive deficits (i.e., attention) associated with the syndrome [70]. The beneficial effects of probiotics are well established, and although their mechanism of action remains unknown, they provide a potential strategy to restore gut microbial communities and diminish inflammation induced by disease and environmental exposures [71].
One of the first in vivo studies to measure the effect of probiotics on behavior involved treating the drinking water of Sprague Dawley rats with a single strain of Bifidobacteria infantis for 14 days. The probiotic treatment did not elicit antidepressant effects, as no changes were found in forced swim test swimming behaviors. However, probiotic treatment caused an increase in blood plasma tryptophan levels, a precursor to serotonin (5-HT). Still, this increase in tryptophan failed to translate into an increase in 5-HT or any other monoamine measured in the brain (e.g., noradrenaline and dopamine) [72]. Alternatively, a 12-week clinical study with a randomized, double-blinded, and placebo-controlled intervention administering one probiotic strain, Lactobacillus plantarum, found reductions in stress and anxiety as measured by a total Depression, Anxiety, and Stress Scale (DASS-42) questionnaire in the probiotic group as compared to the placebo (maltodextrin) control. This change was accompanied by marginal differences in cortisol levels and significant reductions in pro-inflammatory cytokines. Furthermore, this study found that the probiotic group had heightened memory and cognition as measured by the CogState battery test (e.g., increased speed for social emotional cognition and verbal learning and memory) as compared to the placebo group. There were some sex-specific differences as women consuming the probiotic demonstrated marginal improvements in speed of the identification tasks, while men experienced marginal improvements in overall correct input. However, both women and men experienced enhanced cognition at some capacity [73].
In regards to multispecies probiotic intervention in animals, one of the first studies to measure the effects of probiotics on memory was in a diabetic rat model in 2013 [74]. In this study, the probiotic was dissolved in water and consisted of three strains of bacteria: Lactobacillus acidophilus, Bifidobacterium lactis, and Lactobacillus fermentum. The diabetic rats lacking probiotic supplementation took the longest time to find the platform in the MWM and spent the shortest amount of time in the correct quadrant during the retrieval test of the MWM as compared to the non-diabetic rats. However, the diabetic rats supplemented with the probiotic experienced improved spatial memory almost to the point of non-diabetic rat memory capabilities [74]. A clinical study using a multispecies probiotic (Lactobacillus casei W56, Lactobacillus acidophilus W22, Lactobacillus paracasei W20, Bifidobacterium lactis W51, Lactobacillus salivarius W24, Lactococcus lactis W19, Bifidobacterium lactis W52, Lactobacillus plantarum W62, and Bifidobacterium bifidum W23), divided healthy participants into a probiotic, placebo, and no intervention group. Following the intervention, the probiotic group had improved self-reported behavioral measures of positive affect and cognitive reactivity. The probiotic group had altered behavior scores for depression and anxiety questionnaires by increasing positive affect and dampening vulnerability to depression (hopelessness and risk aversion measures). In addition, the probiotic group was found to have improved memory performance, altered brain activation patterns, and reduced decision changes for unpleasant stimuli in the emotional decision making task as compared to the placebo and no intervention control groups. Response accuracy was also found to be increased in the probiotic intervention. Lastly, during the emotional decision-making task, the researchers found significant differences in brain activation patterns between the three groups. Specifically, different activation patterns in the precuneus, mid cingulum, middle temporal gyrus, inferior parietal lobule, and paracentral lobule were detected. There were also significant increases in brain activity in the left anterior cingulum in the probiotic versus nonintervention group. These findings indicate that probiotics play a role in decision making processing and memory performance [75]. Furthermore, multispecies probiotic supplementation of depressive patients over a four-week intervention resulted in reduced IL-6 gene expression, supporting the role of probiotics in reducing inflammation among depressed individuals and potentially among other diseased groups as well [76].
PREBIOTICS AND BEYOND
Prebiotics are non-digestible dietary substances that can alter the composition of the gut microbiota and be fermented by its constituents to produce SCFAs. Prebiotics are typically comprised of carbohydrates, though they can also include flavanols [77]. Several studies have tested prebiotics as a possible therapeutic for gastrointestinal disease, though they have additional potential to improve cognition. In a clinical study, a prebiotic dietary fiber supplement, comprised mainly of polydextrose (PDX) was compared to maltodextrin (placebo). A group of women (n = 18) were administered the prebiotic or placebo over a four-week intervention period. This double-blind, randomized, crossover study revealed improved cognitive flexibility and sustained attention (via CANTAB battery test) in the PDX-treated group compared to controls, representing a moderate improvement in cognitive function. Still, the effects of PDX were limited, and further research on the impact of dietary fiber and prebiotics on gut microbial composition and cognitive function is necessary to better understand the role prebiotics as a supplement and possible therapeutic going forward [78].
Like prebiotics, antibiotics have been a proposed method of improving cognition by inhibiting microglial activation and modulating memory systems. Berens et al. [79] implemented minocycline in a clinical trial of 20 healthy male participants, but contrary to expectations, found reduced overall spatial memory performance. Additional research is needed to determine the potential, if any, for antibiotics to modulate cognitive functions, specifically in the memory space [80]. In essence, the gut-brain-axis is implicated in various disease phenotypes and cognitive outcomes.
MICROBIAL INVOLVEMENT IN COGNITION AND AD
Disruption of the gut microbiota is associated with AD development, and pathogenic microbe elicited neuroinflammation is a proposed etiological link between gut dysbiosis and AD progression. As previously mentioned, APOE genotypes are differentially associated with late-onset AD risk. A study conducted by Tran et al. [80] investigated the relationship between APOE genotypes and the composition of the human gut microbiota [80]. Fifty-six healthy participants ages 56 to 78 were selected according to APOE genotype to determine if there was a unique microbial signature associated with the APOE2/E3, APOE3/E4, or APOE4/E4 genotypes. No significant differences in alpha or beta microbial diversity were found between genotypes; however, differences in relative abundances of bacterial taxa were established, and the APOE genotype was associated with specific gut microbial profiles in human and APOE-transgenic mice. Relative abundance of the phylum Firmicutes and order Clostridiales was heightened in participants with the APOE2/E3 genotype compared to APOE3/E4 or APOE4/E4. At the family level, abundance of Ruminococcaceae, a family of fermentative anaerobes linked to fiber break-down and SCFA production was higher in APOE2/E3 than in APOE3/E3, APOE3/E4, or APOE4/E4 and 3 genera within the family including Clostridium cluster IV, Clostridium cluster XIVa, and Gemmiger, were statistically significant and differentially abundant based on APOE genotypes. This study established an association between Prevotellaceae and Ruminococcaceae and various butyrate-producing genera with APOE genotypes [80]. The differences in taxa among APOE genotypes supports the association between genotypes and gut microbiota composition. Further investigation is necessary to ascertain whether the altered microbiota profile is responsible for exacerbated AD pathogenesis according to APOE genotype.
A recent study provides insight into the relationship between the at-risk APOE genotype (E4 allele) and cognitive function. Rodriguez et al. [81] investigated the effects of the at-risk APOE genotype (E4 allele) and mental work demands on cognitive decline in cognitively healthy adults older than 75 years from the Ageing, Cognition and Dementia in Primary Care Patients multi-centered population-based longitudinal cohort study in Germany. The Mini-Mental State Examination was used to determine cognitive functioning. Information pertaining to participants’ occupational history was also gathered to determine mental demands at work and the relationship between demand level, APOE genotype, and cognitive decline. The researchers found heightened work demands were associated with better cognitive functioning at baseline and slower cognitive decline overtime. In terms of the APOE4 allele, those carrying this allele experienced an accelerated difference in cognitive decline at follow-up. Furthermore, medium versus low work demands were associated with slower cognitive decline among APOE4 carriers. This study better characterized the relationship cognitive decline and the influence of a high-risk genotype for late-onset AD [81].
Another research team utilized fecal samples to characterize the bacterial taxonomic composition of patients with AD diagnoses [16]. Using bacterial 16S rRNA genetic sequencing of DNA isolated from fecal samples, researchers characterized the gut microbial communities of participants with and without clinical diagnoses of dementia due to AD. Here, AD patients were found to possess reduced microbial diversity and maintained compositionally distinct microbiomes compared to age and sex-matched healthy individuals. Phylum through genus wide differences in AD participants’ gut microbiome bacterial abundance were established, including reduced Firmicutes, heightened Bacteroidetes, and reduced Bifidobacterium (Table 1) [16]. AD patients experienced significantly reduced microbiome richness compared to control with respect to ACE and Chao1. Furthermore, Shannon Index and Faith’s PD were significantly decreased in AD participants. In terms of beta diversity, Bray-Curtis dissimilarity and UniFrac analysis demonstrated compositional dissimilarity in the microbiomes of AD versus control groups. Fecal sample analysis revealed reduced abundance at the phylum level of Firmicutes and Actinobacteria, which exhibit anti-inflammatory properties and reduce intestinal permeability, while AD participants experienced increased abundance of Bacteroidetes compared to healthy controls (Table 1) [16]. Families within Firmicutes (Ruminococcaceae, Turicibacteraceae, Peptostreptococcaceae, Clostridiaceae, and Mogibacteriaceae) and the genera SMB53 (family Clostridiaceae), Dialister, Clostridium, Turicibacter, and cc115 (family Erysipelotrichaceae) were less abundant, while the family Gemellaceae and genera Blautia, Phascolarctobacterium, and Gemella were increasingly abundant in AD participants (Table 1). Within Bacteroidetes, Bacteroidaceae and Rikenellaceae at the family level, and Bacteroides and Alistipes at the genus level were more abundant in AD participants (Table 1).
Differences in bacteria taxa and metabolites among patients with MCI or AD and controls (p < 0.05)
*MCI patient versus healthy controls.
The heightened levels of Bacteriodetes found among AD patients aligns with hypotheses related to LPS abundance, inflammation, and AD. Gram negative Bacteriodetes maintain an outer membrane made of LPS; thus, increased Bacteriodetes correlates with greater LPS abundance (Table 1) [82]. Increased presence of LPS in elderly individuals has been associated with heightened gut permeability, microbial translocation, inflammation, and amyloid pathology as LPS can trigger systemic inflammation and release of proinflammatory cytokines post-translocation from the gut to systemic circulation [16, 83]. Additional support for the interplay between gut microbial composition and AD pathology is supported by the negative correlation found between genera abundant in AD and cerebrospinal fluid (CSF) Aβ42/Aβ40 levels since reduced CSF Aβ42/40 is suggestive of heightened amyloid deposition in the brain. Furthermore, positive correlations between bacterial abundance and CSF p-tau and p-tau/Aβ42 were established in this study [16, 84].
PROBIOTIC AND DIETARY INTERVENTIONS IN AD
The use of probiotics in animal and clinical studies of AD have elicited mixed findings. Two different rodent models of AD with distinct probiotic supplementations, were found to improve the disease phenotype. One study utilized a triple transgenic mouse model of AD, B6;129-Psen1tm1Mpm Tg (APPSwe,tauP301L)1Lfa/J (named 3xTg-AD), and the other used a rat model to induce AD via intraperitoneal injection of D-Galactose [87, 88]. Both rodent models displayed reduced cognition as measured by the MWM test and novel object recognition test and presented amyloid plaque burden in the hippocampus [87, 88]. The 3xTg-AD mice were treated with SLAB51, a probiotic comprised of 9 different bacterial strains ((Streptococcus thermophilus, bifidobacteria (B. longum, B. breve, B. infantis), and lactobacilli (L. acidophilus, L plantarum, L. paracasei, L. delbrueckii subsp. bulgaricus, L. brevis)) and the D-galactose induced AD rats were treated with L. plantarum MTCC1325. In response to probiotic treatments, both rodent models experienced improved cognition and reduced plaque burden in the brain. More recently, researchers treated a third model of AD, AppNL–G–F with a probiotic cocktail of eight bacterial strains (L. plantarum, L. delbrueckii subsp. Bulgaricus, L. paracasei, L. acidophilus, B. breve, B. longum, B. infantis, and S. salivarius subspecies, thermophiles) commercially marketed as VSL#3. Probiotic-treated AppNL–G–F mice displayed reduced anxiety-like behaviors and increased c-Fos expression in the hippocampus, a common functional marker for neuronal activity [89, 90]. However, the probiotic cocktail failed to reduce plaque burden, astrogliosis, or microgliosis in the AppNL–G–F mice brains [89]. It should be noted that this failure to improve markers of AD in the brain was only evaluated in female AppNL–G–F and not male mice. A previous study with APPSWE/PS1L166P male and female mice treated with a cocktail of antibiotics found that only male mice had reduced Aβ plaques and changes in microglial phenotype [24]. These studies highlight the need for both male and female in AD research as the disease treatment may be sex-dependent.
A clinical study found results contrasting the findings from studies of AD rodent models supplemented with various probiotics. Leblhuber et al. [91] analyzed serum, fecal samples, and cognitive statuses via MMSE and clock Drawing test (CDT) of 18 patients with AD diagnoses (based on ICD-10, MRT, and routine lab testing) pre- and post-28-day multispecies probiotic supplementation of Lactobacillus casei W56, Lactococcus lactis W19, Lactobacillus acidophilus W22, Bifidobacterium lactis W52, Lactobacillus paracasei W20, Lactobacillus plantarum W62, Bifidobacterium lactis W51, Bifidobacterium bifidum W23, and Lactobacillus salivarius W24 [91]. There were no significant changes in cognitive parameters nor differences in blood serum levels of tryptophan, phenylalanine, and tyrosine after the 28-day probiotic intervention. Though increases were found in serum levels of kynurenine, a metabolite of tryptophan and key regulator of the immune system, in probiotic supplemented individuals [92]. Still, in patients diagnosed with MCI, there is evidence that a probiotic intervention may be of benefit. Patients with MCI experience a unique intermediate period between healthy cognitive function and an AD diagnosis. Addressing symptoms at this time may provide a means of preventing the escalation of cognitive deficits associated with AD. In a 12-week, double-blind, placebo-controlled, randomized clinical trial, an 800 mg/day Lactobaccillus plantarum C29-fermented soybean intervention improved cognition among the probiotic-treated group [93]. Both probiotic and placebo groups were at least 90% compliant in following daily dosage prescriptions. Post-test, patients diagnosed with MCI based on DSM-5 criteria and treated with the probiotic experienced enhanced composite scores in cognitive function (attention, working memory, verbal memory) based on results from computerized neurocognitive function tests. Specifically, the probiotic group demonstrated significant score increases in attention. Lastly, the probiotic group experienced significant increases in the Lactobacilli bacterial population and serum BDNF levels which is positively associated with enhanced combined cognitive function scores [93]. Akbari et al. also offers support for the use of probiotics in combating cognitive decline [94]. After a 12-week study, cognitive functionality, assessed via MMSE scores, was improved in 30 AD patients treated with a probiotic cocktail containing Lactobacillus acidophilus, Lactobacillus casei, Bifidobacterium bifidum, and Lactobacillus fermentum, compared to patients treated with control milk. MMSE scores improved by 27.90% ±8.07 among those treated with the probiotic cocktail, while scores declined by 5.03% ±3.00 among control participants (p < 0.001) [94]. This significant difference in cognitive function between treatment groups indicates probiotics may have a therapeutic application in ameliorating symptoms associated with AD [94]. A recent review by Arora et al. also highlights the use of probiotics as anti-inflammatory and antioxidant-containing agents and the potential limitations as a therapeutic [95].
Dietary intervention must be considered in addition to probiotics when discussing microbiome-based therapeutics as diet is proposed to influence the gut-brain axis via changes of the gut microbiota composition. The shared incidence between AD and illnesses that diet directly influences, including obesity, cardiovascular disease, type 2 diabetes, and metabolic syndrome point us to a connection between diet and AD pathology [96]. There is evidence to suggest that the use of certain nutrient combinations can combat the synaptic loss associated with Parkinson’s disease [97]. A similar line of thinking may be applied to AD. Rather than supplementing the diet with individual nutrients, a more comprehensive approach shows promise in combating AD with diet. For example, uridine, docosahexaenoic acid (DHA), and choline are phospholipid precursors responsible for the production and maintenance of neuronal membranes, and their systemic availability has been found to be reduced in patients suffering from AD [96, 97]. By increasing the abundance of these nutrients and cofactors associated with phospholipid synthesis (i.e., vitamins B, C, E, and selenium) via dietary intervention, circulating levels in blood and CSF may be increased, resulting in heightened phospholipid production and maintenance of neuronal membrane integrity. In addition to individual nutrients, dietary fat intake may modify the risk of developing AD by altering the body’s response to environmental neurotoxins, and preventative dietary interventions are thought to reduce colonic inflammation. Lastly, poly-unsaturated fatty acids like docosahexaenoic acid have anti-inflammatory effects and may reduce reactive oxygen species and oxidative stress associated with AD [97].
Healthy dietary patterns, including those primarily comprised of plant-based foods, probiotics, antioxidants, soybeans, nuts, and omega-3 polyunsaturated fatty acids and minimally containing saturated fats, animal-derived proteins, and refined sugars have been shown to reduce neurocognitive impairment risk and eventual onset and diagnosis of AD [96]. Diets comprised of such rich foods have been shown to elicit a positive impact on the body’s inflammatory and immune response and reduce insulin resistance and risk of neurocognitive impairments [96]. The Mediterranean diet, a cultural diet incorporating healthy dietary practices from countries surrounding the Mediterranean Sea (i.e., France, Greece, Italy, and Spain) and DASH (Dietary Approach to Stop Hypertension) have been researched for their vast applications in medicine, including their ability to reduce cardiovascular diseases and other risk factors [96, 98]. These diets are hypothesized to slow cognitive decline and have been found to elicit more benefits than single nutrient supplementation [96, 98]. In turn, MIND (Mediterranean-DASH diet Intervention for Neurodegenerative Delay) was developed as a hybrid of the Mediterranean and DASH diets. MIND focuses on brain protection by incorporating factors from both diets found to enhance cognitive performance and lower dementia and AD risk [96, 98]. Specifically, the MIND diet deviates from the others by emphasizing the value of berries over general fruit consumption and recommending one meal containing fish per week [98]. Further, the MIND diet highlights the importance of green leafy vegetables and plant-based foods and restricts consumption of animal products and those containing high levels of saturated fat [98]. Morris et al. conducted a study with participants from the Memory and Aging Project and found the MIND diet notably reduced cognitive decline over an average 4.7 years over the study duration. By scoring the dietary practices of each of the 960 participants on a scale, they determined that the difference in cognitive scores and function between the top and bottom tertiles was equivalent to being 7.5 years younger [98]. Additional studies support the substantial influence of the MIND diet on cognitive acuity; thus, the role of dietary practices in microbiota health, cognition, and AD must be studied further to understand the power of dietary intervention in mitigating AD [99–101].
FECAL MICROBIOTA TRANSFER AND HUMANIZED GNOTOBIOTIC MICE IN AD
The application of FMT as a therapeutic for AD patients remains unclear. However, a study of humanized gnotobiotic mice provides additional support for the use of FMT as a tool to study the gut-brain axis specifically in AD. In this study, FMT from a human AD donor to GF mice reduced cognitive function earlier than FMT from a healthy non-AD donor to GF mice. In fact, mice treated with FMT from human AD patients performed significantly worse on an object location test and object recognition test at 55 weeks of age as compared to mice with FMT from healthy non-AD donors [102]. There are limitations to the use of humanized gnotobiotic mice as the GI anatomy, physiology, and diet between mice and humans are different [103]. There is also low engraftment from human donor samples to GF recipient mice and the gut microbiota of humanized mice only partially resembles that of the human donor [104, 105].
SHORT CHAIN FATTY ACIDS IN AD
SCFAs are carboxylic acids 2–6 carbons long and are the metabolites produced by gut bacteria during the fermentation of fiber [37]. The main SCFAs produced from gut bacteria metabolism of carbohydrates are acetate, butyrate, and propionate [106]. In the context of AD, butyric and propionic acid have the ability to disrupt the formation of Aβ oligomers in vitro and AD mice have been shown to exhibit lower levels of total SCFA in the brain as compared to WT mice [107, 108]. Additionally, when a late disease stage mouse model of AD was treated with sodium butyrate, animals experienced enhanced associated memory, though plaque load in the brain remained the same. The improved associated memory in AD mice treated with sodium butyrate was accompanied by an increase in the expression of genes related to memory consolidation in the hippocampus [109]. In an alternate mouse model of AD, 5XFAD, animals were treated with sodium butyrate in their food at an early stage of disease progression (2 months) rather than a late stage (14-15 months). This 12-week intervention, resulted in a 40% reduction in brain Aβ levels and significantly improved associative learning and cognitive function [110]. Additionally, cognitively impaired mice (LPS-induced and APP/PS1 transgenic) treated with a specific bacterium, Agathobaculum butyriciproducens, which mainly produces butyric acid showed improved memory recognition and spatial memory as measured by a novel object test and a spontaneous Y-maze alternation test. The A. butyriciproducens-treated LPS and APP/PS1 mice also displayed reduced microglial activation and in the APP/PS1 treated mice they displayed reduced Aβ plaque deposition [111].
As exciting as the possibility is to use SCFAs as a therapeutic for AD, contradicting research exists. In a recently published study by Colombo [112], GF APP/PS1 mice presented with lower plaque burden and improved memory as compared to the re-conventionalized GF AD and SPF AD mice. Further evidence shows the SCFA treatment, consisting of sodium propionate, sodium butyrate, and sodium acetate, in the SPF and GF AD mice caused a larger plaque burden in the brain as compared to GF AD mice [112]. Additionally, treatment of a GF Parkinson’s disease mouse model with SCFAs increased the motor dysfunction of these mice and further increasing the controversy surrounding the benefit of SCFA use [113, 114]. However, these data should also be examined with appropriate limitations. As previously described, GF mice have altered brain development and are behaviorally and neurochemically different from conventional mice [50–52]. GF mice also have immature immune systems and in the context of the brain they have naïve microglia with decreased responses to viral infections [115]. Furthermore, GF mice have been shown to lack a working and non-spatial memory as measured by a T-Maze and novel object recognition test [116]. Therefore, care should be exercised in the examination of potential therapeutics with a GF mouse model.
CONCLUSIONS
Significant progress has been made in understanding the relationship between the gut microbiota and the brain; the potential to mitigate gut dysbiosis and its associated diseases exists via a variety of intervention strategies. Figure 1 highlights the current understanding of the factors influencing gut dysbiosis and the associated pathogenesis of cognitive decline and AD. Considerable research must still be conducted in order to understand how the gut microbiota may be leveraged to treat cognitive disorders, including AD.
Regarding the use of rodent models, we suggest: The use of more and varied rodent models of AD with probiotic, FMT, or SCFA treatment before symptomatic AD, during mild cognitive impairment period, and at the late stage of cognitive impairment. Special attention should be paid to the sex and the microbial state (GF versus SPF) of the animal models used, as the effects of any treatment could vary based on these factors. Various outcomes are expected, but this is a necessary strategy to reach a consensus and to establish which (if any) therapeutics are worth further study and at what stage of the disease. The use of different probiotics containing single or multiple strains and different concentrations. The use of single and multiple strains of bacteria in a probiotic supplementation would aid in identifying useful strains and whether any work synergistically to the benefit of the host. The use of humanized gnotobiotic mice with humanized gut microbiotas. To better understand the interactions between the gut microbiota and brain in the context of human health and disease while maintaining the flexibility of an animal model, it will be necessary to use humanized gnotobiotic mice. This animal model provides more clinical relevance and translation to human health.
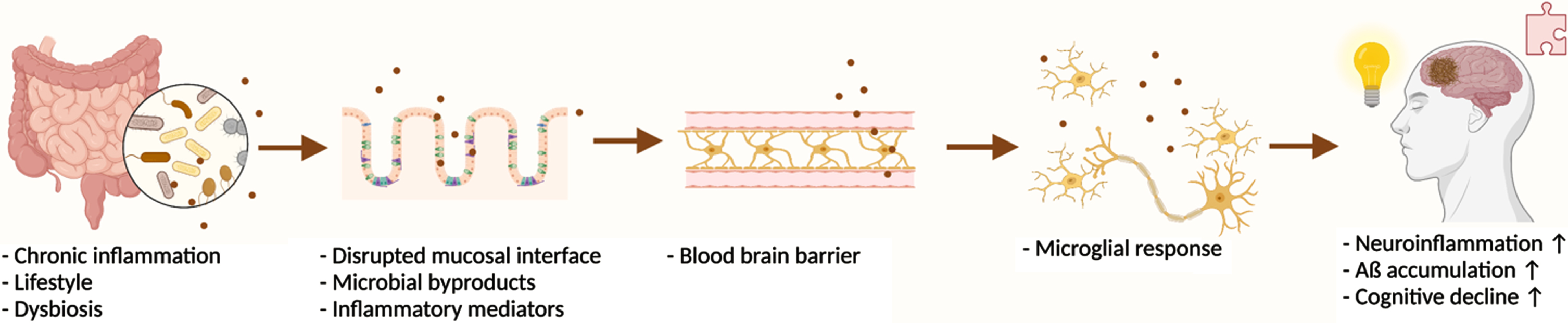
The proposed pathogenesis of cognitive decline and AD development via gut microbiota disturbances and inflammation. Created with BioRender.com.
For clinical studies, we recommend the increased abundance and variation of interventions such as different durations, probiotic compositions, and stages (pre-clinical, mild, late) of AD and potentially FMTs. This will allow researchers to better characterize strategies to mitigate the presence and progression of this widespread and devastating disease.