
Abstract
Animal studies increasingly indicate that the gut microbiota composition and function can be involved in the pathophysiology and progression of Alzheimer’s disease (AD) at multiple levels. However, few studies have investigated this putative gut-brain axis in human beings, and none of them considered diet as a determinant of intestinal microbiota composition. Epidemiological studies highlight that a high intake of fruit and vegetables, such as that typical of the Mediterranean diet, can modulate AD progression. Thus, nutritional interventions are being increasingly studied as a possible non-pharmacological strategy to slow down the progression of AD. In particular, polyphenols and fibers represent the nutritional compounds with the higher potential of counterbalancing the pathophysiological mechanisms of dementia due to their antioxidant, anti-inflammatory, and anti-apoptotic properties. These actions are mediated by the gut microbiota, that can transform polyphenols and fibers into biologically active compounds including, among others, phenyl-γ-valerolactones, urolithins, butyrate, and other short-chain fatty acids. In this review, the complex mechanisms linking nutrition, gut microbiota composition, and pathophysiology of cognitive decline in AD are discussed, with a particular focus on the role of polyphenols and fibers. The gaps between pre-clinical and clinical studies are particularly emphasized, as well as the urgent need for studies comprehensively evaluating the link between nutrition, microbiome, and clinical aspects of AD.
INTRODUCTION: ALZHEIMER’S DISEASE AND DIET
A large body of evidence linking dietary patterns with cognitive function in aging has emerged from population-based studies in recent years [1, 2]. Namely, in older community-dwellers higher adherence to Mediterranean-style diet has been associated with better cognitive performance [3, 4], lower frequency of subjective cognitive complaints [5], and lower prevalence and incidence of mild cognitive impairment (MCI) and dementia [6, 7]. Host-related genetic factors, ethnicity, and gender can modulate these associations, but do not substantially modify the assumption that a healthy dietary pattern has a positive influence on the pathophysiology of Alzheimer’s disease (AD) and other types of dementia [8, 9].
The Mediterranean diet is a dietary model which has been widely studied for the pivotal role in the maintenance of the health status and the prevention of several disease risk factors [10], among which neurological disorders [11]. The daily and abundant consumption of plant-based food groups, such as fruit, vegetables, cereals, and pulses, leads to the intake of several nutrients that exhibit biologically activities on homocysteine metabolism (B-group vitamins), oxidative stress (vitamin C, vitamin E), neural inflammation, degeneration, and plasticity (vitamin D, n-3 fatty acids) [12, 13]. Such foods are also a great source of non-nutrient bioactive compounds, like polyphenols, with antioxidant, anti-inflammatory, and putative neuroprotective properties [14–16]. Not only the Mediterranean diet, but also the adherence to other dietary patterns in which the intake of such nutrients and bioactives is particularly emphasized and with reduced intakes of rapidly digested carbohydrates, saturated fatty acids, and salt, like the Mediterranean-DASH for Neurodegenerative Delay (MIND) diet or the NU-AGE diet, is associated with better cognitive performance and slower age-related cognitive decline [17, 18].
The ketogenic diet, with very low carbohydrate and high lipid content, has also gained attention for its potential neuroprotective action, especially in the prodromal phases of AD [19, 20]. Despite the promising findings of in vitro and animal studies, mainly on the effects of ketones on the mitochondria and their cellular energy-related functions, there still is a lack of clinical studies, as well as a low quality of evidence from the few running ones, to recommend these diets for the prevention of dementia [21, 22].
Conversely, the presence of malnutrition in older age, with insufficient energy and micronutrient intake, is a well-established risk factor for the onset and progression of MCI and dementia [23, 24]. Diets including an excessive intake of animal proteins, saturated fatty acids, and rapidly digested carbohydrates, are also associated with higher risk of MCI and dementia, and with increased cerebral amyloid-β deposition [25, 26]. Interestingly, recent evidence from animal models also links excessive salt intake with progression of dementia through promotion of tau phosphorylation, a central mechanism of neurodegeneration [27].
Despite the evidence from epidemiological and preclinical studies, the effective role of nutrition in the prevention and treatment of neurodegenerative disorders is far from being fully understood. Nutritional interventions for primary or secondary prevention of AD in older subjects have shown only limited benefits, and, to date, diet has only a secondary role in the clinical management of dementia [12, 29]. Apart from methodological and ethical issues making the design of specific trials in this field very challenging, there are also other elements that must be considered, among which the impact of food intake on the whole organism, by starting from the digestive tract where nutrients are metabolized and absorbed.
In fact, diet represents one of the main factors contributing to shape the intestinal microbiota composition and function. Conversely, the microbiota of each individual interacts with food compounds in a complex way, influencing the whole physiology of the host. The microbiota can also influence the pathophysiology of dementia through multiple mechanisms [30, 31], and for these reasons the relationship between diet and dementia should be absolutely unraveled by also considering the key role of the gut microbiota and its metabolic activity.
Unfortunately, the interplay between diet, microbiota composition, and cognitive function has not been comprehensively studied to date. The current state of knowledge allows to hypothesize that dietary patterns can modulate the pathophysiology of AD through mediation of the gut microbiota, and that the consumption of certain types of foods can positively modulate cognitive function only in the presence of specified microbiota composition. However, these assumptions are supported only by indirect evidence, mainly coming from preclinical and animal studies. The aim of this review is to discuss the most recent advances in this field, with a particular focus on how dietary polyphenols and fibers can influence the pathophysiology of AD through mediation by the intestinal microbiota.
THE GUT-BRAIN AXIS AND ALZHEIMER’S DISEASE: PATHOPHYSIOLOGICAL MECHANISMS
The term “gut-brain axis” is commonly used to describe a bidirectional channel of communication between the gastrointestinal tract and the central nervous system (CNS). More recently, this term has been extended to “microbiota-gut-brain axis”, in light of the accumulating evidence obtained from germ-free animals as well as probiotics, antibiotics, and infection studies on the impact of the intestinal microbiota on this gut-brain interaction [32].
Several neural, immune, endocrine, and metabolic pathways of communication between the gut microbiota and the CNS have been proposed. Undoubtedly, one of the most important is mediated by the vagus nerve, which conveys information between the gastrointestinal tract and the CNS in both afferent and efferent direction [33]. Relatedly, several microbial-derived intermediates, including short-chain fatty acids (SCFAs) and tryptophan metabolites [34, 35], interact with enteroendocrine cells, enterochromaffin cells, and the mucosal immune system, and might propagate bottom-up signaling via vagal and/or spinal afferents [36–38]. In addition, some of these microbial-derived molecules cross the intestinal barrier, entering portal and then systemic circulation, and might even reach brain sites directly [36–38]. The microbiota can also produce or contribute to the production of several neuroactive molecules including gamma-aminobutyric acid, serotonin, norepinephrine, and dopamine [39–42]. Yet, it is unclear whether these neurotransmitters reach relevant receptors in the brain or achieve sufficient levels to induce CNS responses.
In the past decade, advances in sequencing technology, metabolomics, and neurophysiology have allowed a deeper investigation of the cross-talk between the gut and the CNS via the microbiota both in health and disease states [43, 44]. An involvement of gut microbiota in the pathogenesis of neurodegenerative diseases, including AD, has thus been hypothesized [45, 46]. The association between gut microbiota and AD might be related to the central role of inflammation in the development and progression of this neurodegenerative disorder [47]. Indeed, the gut microbiota is a source of a large amount of amyloids—though different in their primary structure from the amyloids in the CNS –lipopolysaccharides, and other toxins that may contribute to systemic inflammation and disruption of physiological barriers [48]. Bacteria and/or their products can then migrate from the gastrointestinal tract to the CNS, especially in older subjects, where gut mucosa permeability tends to increase. In the CNS, bacterial products might promote neuronal amyloid aggregation and neuro-inflammation, ultimately leading to neural injury and degeneration [44, 50]. For example, rats exposed to bacterial amyloids displayed increased neuronal alpha-synuclein deposition in both the gut and the brain, enhanced microgliosis and astrogliosis, and increased central levels of proinflammatory cytokines compared to rats exposed to bacteria without the ability to produce amyloids [51].
Animal models have also been useful for gaining a better understanding of the relationship between microbiota dysbiosis and AD-like pathology. For example, in a series of elegant studies using amyloid precursor protein (APP) transgenic mice, a well-established preclinical model of AD, analysis of bacterial 16S rRNA from fecal samples revealed significant changes in gut microbial composition of transgenic mice compared to wild-type counterparts [52]. Remarkably, germ-free APP transgenic mice exhibited a less severe cerebral amyloid-β pathology compared with control mice with intestinal microbiota, suggesting that the absence of microbiota may retard the progression of AD-like pathology [52]. Intriguingly, colonization of germ-free APP transgenic mice increased cerebral amyloid-β to a greater extent when the microbiota originated from conventionally-raised APP transgenic mice compared with wild-type mice [52].
Supporting these findings, another study in APP transgenic mice revealed a shift in microbial composition compared to wild-type mice, as shown by the higher abundance of Helicobacteraceae and Desulfovibrionaceae at the family level and Odoribacter and Helicobacter at the genus level, and the lower abundance of Prevotella [53]. Notably, AD-related histological (i.e., amyloid plaque burden) and behavioral (i.e., impaired spatial learning and memory) features were found to be correlated with the specific microbiome state of animal models [53]. A subsequent study investigated age-related changes in the microbiota of APP transgenic mice and found that AD pathology shifted gut microbiota composition during aging towards an inflammation related bacterial profile (i.e., Proteobacteria and Erysipelotrichaceae), suggesting that these changes could contribute to disease progression and severity [54].
From a theoretical point of view, an abnormal microbiota composition characterized by a high abundance of proinflammatory bacteria and low abundance of anti-inflammatory bacteria may generate a systemic inflammatory response through a defective (i.e., “leaky”) gut barrier. This inflammatory response may, in turn, impair the blood-brain barrier, promote neuro-inflammation, and contribute to the pathogenesis of AD [49, 50]. The fecal transplantation from transgenic mice with dementia to cognitively healthy mice induced a worsening of cognitive abilities [55], suggesting a correlation between the gut microbiota composition and cognitive dysfunction. Similarly, studies based on Drosophila models of dementia [56] and mouse models of stroke [57, 58] showed an association between gut microbiota dysbiosis, cognitive functions, and clinical course of the disease.
Supporting this view, animal studies have also shown that substances that are able to modify microbiota composition, such as antibiotic agents or probiotics, can modulate inflammatory responses and positively or negatively affect the progression of the disease. For example, long-term antibiotic treatment induced perturbation in gut microbial diversity and altered peripherally circulating cytokine/chemokine composition in APP transgenic mice [59]. This was associated with a reduction in amyloid plaque deposition and elevated levels of soluble amyloid-β, reduced plaque-localized gliosis, and altered micro-glial morphology [59]. In other experimental studies, antibiotic-induced gut dybiosis was instead associated with increased systemic inflammation and poorer cognitive performance in mice [60, 61].
Likewise, gut microbiota manipulation induced by administration of probiotics belonging to Bifidobacterium spp. [62–65] or Lactobacillus spp. [62, 67] in transgenic mouse models of AD resulted in improved cognitive performance and reduced markers of inflammation and neuropathology. For example, in one of these studies four-week treatment with a probiotic formulation (i.e., SLAB51, a mixture of lactic acid bacteria and bifidobacteria) in the early stage of the disease led to an increase in the abundance of anti-inflammatory Bifidobacterium spp. and a reduction in the abundance of proinflammatory Campylobacterales [62]. These changes in microbiota composition were associated with a reduction in the levels of proinflammatory cytokines and a less severe cognitive decline, due to a reduction in brain damage and reduced accumulation of amyloid-β aggregates [62]. Moreover, this probiotic regimen caused a reduction of oxidative stress, another important element involved in the pathogenesis of AD [62].
Few studies also demonstrated that the manipulation of the gut microbiota of mouse models of AD through administration of functional foods, like fructooligosaccharides, flavonoids, or tea saponins resulted in improved cognitive functions [67–70].
Despite this body of evidence, it must be remembered that the results of animal studies may not be immediately transferrable to human beings. Moreover, microbiota-gut-brain communications are bidirectional and the possibility that any changes observed in the microbiota are secondary should be always considered. For example, the activity of the hypothalamic–pituitary–adrenocortical axis and the sympathetic nervous system may be increased in AD, thereby influencing top-down signaling to the gastrointestinal tract [71, 72]. Therefore, activation of these neuroendocrine stress response systems may contribute to the change of microbiota profile, both directly via host-enteric microbiota signaling and indirectly via changes in the intestinal milieu [73].
THE GUT MICROBIOTA IN ALZHEIMER’S DISEASE: HUMAN STUDIES
In the last decade, several studies have investigated the correlations between the fecal microbiome and presence of acute or chronic illness [74–76]. Notably, several microbial taxa of the intestinal microbiota play a crucial role in maintaining host homeostasis by modulating immunological, nutritional, metabolic, and physiological functionalities, from infancy to senescence [77–80]. The alteration of this symbiotic homeostasis may be involved in the onset and progression of diseases, not involving only the gastrointestinal system [77–80]. This concept also applies to CNS pathology [81]. Remarkably, the human intestinal microbiota can influence cognitive function, and intestinal bacteria may be involved in the pathophysiology of neurodegenerative disorders, including multiple sclerosis, Parkinson’s disease, and of course AD [81].
Several studies, mainly conducted with metagenomics sequencing of fecal samples, have explored the possible role of gut microbiota in the pathogenesis of dementia, particularly of AD, highlighting alteration in the intestinal microbial composition of patients in comparison with controls [82–89]. The findings of these studies are summarized in Table 1.
Overview of the findings of the main human studies investigating gut microbiota composition in AD
AD, Alzheimer’s disease; MCI, Mild Cognitive Impairment; US, United States.
As highlighted by Vogt and colleagues [83], one of the main differences in the gut microbiota composition, determined through the use of 16S rRNA gene profiling approach, between AD patients and controls is represented by reduced bacterial richness and diversity in subjects with dementia. Moreover, a distinct bacterial profile seems to characterize the gut microbiota of AD patients, who display high abundance of species belonging to Bacteroidetes phylum, such as Bacteroides spp. and Alistipes spp., and reduced representation of members of the Bifidobacterium genus and taxa belonging to Actinobacteria, and Firmicutes phyla [83, 89].
Remarkably, overrepresentation of Bacteroides fragilis in the microbiota has been associated with the development of AD, probably due to its production of lipopolysaccharide, which could contribute to systemic inflammation [90]. Two studies from China also show increased representation of bifidobacteria in fecal samples of patients with AD, suggesting that this genus may represent a microbial marker of AD [88, 89]. However, bifidobacteria were underrepresented in the fecal microbiota of patients with AD in the Vogt study [83], and the genus Bifidobacterium is generally associated with longevity and healthy aging [91].
In one study, the gut microbiota of AD patients was also characterized by low abundance of butyrate-producing taxa, such as members of the Butyrivibrio, Eubacterium, Roseburia, and Faecalibacterium genera [86]. Furthermore, deep metabolomic investigations based on microbiome data of AD patients compared to individuals without dementia highlighted a decrease in butyrate-coding genes in AD disease [86]. These findings might suggest that lower proportions of butyrate-producing species would contribute to the onset of the disease.
Moreover, AD patients displayed a tendency towards increase in relative abundance of bacterial species related to human diseases or able to act as opportunistic pathogens, like Klebsiella pneumoniae, Bacteroides fragilis, and Eggerthella lenta. These species could be characteristic of a pro-inflammatory microbiota, which might contribute to the pathophysiology of AD.
Oddly, Emery et al. explored the bacterial load of frozen and fixed postmortem tissue from AD and control temporal cortex [93]. This study reported that AD brains tend to have higher bacterial load than controls, reinforcing the notion of a possible correlation between human gut microbiota and the development of AD [93].
Two recent systematic reviews and meta-analysis have considered the human studies where cognitive function was measured as one of the clinical endpoints after administration of probiotics or prebiotics to subjects suffering from dementia or MCI [94, 95]. The large majority of these studies were of poor methodological quality, with small sample size, absence of randomization of treatments and, sometimes, absence of a control group receiving standard of care treatment [94, 95]. Only five studies, with an overall number of 297 participants, had a randomized controlled design and sufficient methodological quality [96–100]. These studies showed improvement in cognitive function of participants after probiotic administration, mainly consisting in blends of bifidobacteria and lactobacilli, supporting the existence of a gut-brain axis also in human beings with AD [94–100].
However, the clinical significance of these findings, and their impact on the future management of AD is still unknown. More importantly, none of these studies conducted on human beings considered diet as a confounding variable or as a possible modulator of gut microbiota composition.
POLYPHENOLS AND GUT MICROBIOTA IN ALZHEIMER’S DISEASE
Overview of polyphenols and their gut metabolites
The consumption of foods rich in polyphenols has been associated with numerous beneficial effects on the risk and development different neurodegenerative diseases [101]. Among polyphenols, positive findings have been highlighted for chlorogenic acids (mostly present in coffee, leafy vegetables, and berries), curcumin (mainly present in turmeric), and resveratrol (a stilbene present in grapes and wine) [102], but the most robust evidence is related to flavonoids, the most abundant subclass of polyphenols [103].
Two recent surveys conducted on US adult cohorts “The Nurses’ Health Study”, “The Health Professionals Follow-up Study” [104] and “Framingham Offspring Cohort” [105] confirmed that the long-term intake of flavonoids significantly decreased the risks of developing AD and related dementias and subjective cognitive decline markers. Among flavonoid subclasses, the most positive findings on cognitive health have been found for flavones, flavonols, and anthocyanins [102, 105]. In another survey, based on 921 older subjects from the US, the authors extrapolated intake data for single phenolic compounds, observing that high dietary flavonol intake, and particularly of kaempferol and isorhamnetin, may be protective against the development of AD [106].
Despite the in vitro experimental findings of the last decades confirm these molecular mechanisms, one should not forget the complexity of human physiology and, in particular, of the digestive system, and the now proven extensive catabolism of polyphenols in the lower gastro-intestinal tract has induced scientists to re-think what the real biological effectors might be. In fact, once ingested, the polyphenols present in planta pass through the oral cavity and stomach almost unmetabolized. Once in the small intestine, only a small amount is absorbed and able to reach the liver and, subsequently, the systemic circulation [107]. The most relevant fraction of ingested polyphenols reaches the colon and undergoes substantial modifications of the parent chemical structures through the action of bacterial enzymes. Thus, most of the biological effects of (poly)phenols are mediated by the intestinal microbiota. The generated bacterial metabolites are then absorbed and may undergo hepatic phase I and, most prominently, II metabolisms [108].
The result of such biochemical transformation is the release in the bloodstream of smaller phenolic compounds, including benzene diols and triols, benzaldehydes, benzoic acids, phenylacetic acids, phenylpropanoic acids, cinnamic acids, hippuric acids, and many other compounds, which might sometimes be very specifically linked to their parent phenolic structure [109]. For this reason, in the most recent years, the studies focusing on the role of (poly)phenols on health outcomes are considering, as real biological effectors, the colonic metabolites rather than the compounds that are naturally present in the food source. A recent review by Carregosa and collaborators [109] summarized in a very exhaustive manner the role of the several metabolites, categorized for the single deriving subclasses of flavonoids, on several markers of neurodegenerative diseases, among which AD, tested in very different model organisms, from fungi to humans.
Flavan-3-ols
Flavan-3-ols are among the most representative contributors of the flavonoid intake in several European countries [110–112]. A recent review from our group comprehensively described the possible colonic metabolites following flavan-3-ol ingestion, showing that phenyl-γ-valerolactones, phenylvaleric acids, and their glucuronide, sulfate, and methylated forms are the most abundant in humans [113]. In a study using different in vivo animal models, Angelino et al. [114] also demonstrated that one of these metabolites, namely 5-(hydroxyphenyl)-γ-valerolactone-sulfate (3′,4′ isomer), was effectively detected in brain tissues of the different animals used. Concerning the biological activity, recently our group evaluated the role of such metabolites in counteracting the formation of amyloid-β oligomers (AβO) in yeast, mammalian cell, and mouse models, at physiological concentrations [115]. Results showed that 5-(4′-hydroxyphenyl)-γ-valerolactone was able to counteract the toxicity induced by the formation of oligomers, but not fibrils [115]. The mouse model, characterized by individuals with AβO-induced memory impairment, showed a relieve of the brain tissue morphology and an improvement of the memory functions when the animals were co-exposed to the metabolite [115]. The same compound, as well as 5-(3′,4′-hydroxyphenyl)-γ-valerolactone and 5-(3′-hydroxyphenyl)-γ-valerolactone-4′-sulfate, tested up to 5μM, was effective in modulating cellular proteolysis via proteasome inhibition and consequent autophagy upregulation, together with the inhibition of cathepsin B activity [116]. This led to the decrease of the amount of intra- and extracellular Aβ1 - 42 peptides.
Two recent studies aimed at investigating the amelioration from the AβO toxicity by using extracts rich in flavan-3-ols in cell culture models and confirmed the presence of flavan-3-ol metabolites in murine models [116, 117]. Another study evaluated the role of a lotus extract (Nelumbo nucifera) in the reduction of cellular deformation and apoptosis rate in cells pre-incubated with Aβ25 - 35 by affecting different cascades involving brain-derived neurotrophic factor (BDNF), phosphorylation of cAMP-responsive element-binding (CREB), protein kinase B (also known as AKT), and the extracellular signal-regulated kinase (ERK) [116]. Authors evaluated the bioavailability of the extract by feeding rats with a 200 mg/kg bole dose of extract for two days, and confirmed the presence of small phenolic acids, mainly caffeic, vanillic, and p-hydroxyphenylacetic acids (no conjugated metabolites were measured as β-glucuronidase/sulfatase enzymes were used before analysis) [116]. Similarly, a study from the US demonstrated that different concentrations of a pine bark extract, rich in proanthocyanidins and (epi)catechin, was able to block the formation of aggregates of AβO and tau protein [117]. Authors studied the bioavailability of the extract by feeding bole doses of 200 mg/kg pine bark extract to rats for then days, identifying in plasma and brain tissues the methylated and glucuronidated conjugates of catechin and epicatechin [117].
Concerning human studies, Desideri et al. supplemented with increasing concentrations of cocoa flavan-3-ols, up to 990 mg/day for 8 weeks, older subjects with cognitive impairment, and showed a significant amelioration of the cognitive functions of participants consuming high doses of flavan-3-ols [118]. Moreover, Brickman et al. used the functional magnetic resonance imaging technique to demonstrate that healthy active older subjects consuming cocoa flavan-3-ols (900 mg/day) had an improvement of the cerebral blood flow in the body of the hippocampal circuit, primarily to the dentate gyrus and subiculum, driver of the age-related cognitive decline [119].
Different projects are now running to improve our knowledge of the role of (poly)phenol-rich foods on cognitive parameters. Among these, the “Cognitive Ageing, Nutrition and Neurogenesis (CANN)” trial is evaluating the effects of the supplementation of long-chain ω-3 polyunsaturated fatty acids plus 500/day of cocoa flavan-3-ols, for 12 months, on cognitive markers in older subjects from the UK and Australia with mild cognitive or subjective memory impairments [120]. The ”Valerolactones and healthy Ageing: LInking Dietary factors, nutrient biomarkers, metabolic status and inflammation with cognition in older adults (VALID)” project aims to demonstrate the validity of the phenyl-γ-valerolactones as biomarker of flavan-3-ol intake and to investigate whether the consumption of foods and beverages rich in flavan-3-ols—and so the presence of specific colonic metabolites–may have a role in the preservation of the cognitive health in the Irish TUDA elderly cohort participants [121].
Ellagitannins
Pomegranate (Punica granatum L.) is a rich source of ellagitannins, a class of condensed tannins which are very poorly absorbed and, instead, reach the colon where colonic bacteria metabolize them to form a group of compounds named urolithins [108]. Pomegranate has been supplemented in different animal studies to evaluate putative biological effects on AD and more general cognitive markers. A Brazilian study revealed that the supplementation of pomegranate peel extract (800 mg/kg/die) for 35 days to mice infused with AβO was able to decrease amyloid plaque density and the expression of several brain inflammatory features, in addition to improving animal behavior, compared to control mice [122]. While in the latter study there is no focus on the colonic metabolites, Kujawska et al. identified quantifiable amounts of urolithin A in brain tissue of rats treated with rotenone to induce cognitive impairment and after the administration of pomegranate juice (500 mg/kg/die) for 35 days [123]. The authors demonstrated that the pomegranate juice supplementation, and putatively the presence of the microbial metabolite urolithin A, was able to decrease the expression of different oxidative, neuroinflammatory, and apoptotic markers [123]. The effect of single urolithins has been investigated both in vitro and through animal studies. DaSilva and collaborators evaluated the modulation of inflammation by urolithin A, B, and their methylated forms, up to 10μM, in single LPS-stimulated microglia cells or in co-culture with neuroblastoma cells [124]. Results showed that all the compounds were able to reduce some neuroinflammatory mediators, like nitric oxide, interleukins, prostaglandins, and pro-apoptotic factors, compared to control. Finally, Gong and colleagues supplemented transgenic AD mouse models with 300 mg/kg urolithin A for 14 days and found an attenuation of neuron death through the inhibition of AβO deposition and a trigger of neurogenesis via inhibition of several markers of neuroinflammation and apoptosis [125].
Anthocyanins
Anthocyanins are the main contributors to the red/purple color of fruit and vegetables and include compounds such pelargonidin, cyanidin, delphinidin, peonidin, petunidin, and malvidin and their glycoside conjugates. Their main colonic metabolites are small phenolic acids, as protocatechuic, gallic, vanillic, syringic, and hydroxybenzoic acids [108]. A recent and exhaustive review pointed out that the main metabolites showing in vitro neuroprotective effects were protocatechuic, gallic, and vanillic acids, limited for other phenolic acids [126]. The most important target of these phenolic metabolites, mostly protocatechuic and vanillic acids, were the antioxidant activities at intracellular level, so counteracting the depletion of glutathione and catalase activities due to peroxide stimulation or improving cell viability after nitro-radical increase in the cell [127, 128]. Not only antioxidant, i.e., superoxide dismutase or catalase, but also inflammation enzymes, i.e., cyclooxygenases, were also inhibited by gallic acid treatment of neuronal cells, despite in high concentrations, with consequent decrease of cytokine production [129].
Concerning immortalized cells, several studies pointed out that gallic, vanillic, and hydroxybenzoic acids are able to block the oligomerization of Aβ, destabilize pre-fibril and fibril formation, by structural interaction, by metal ion chelation, or by decreasing neuro-inflammatory and apoptotic mediators [130–132]. These findings have been also confirmed in numerous animal models, with an improving of the endogenous antioxidant enzyme activities, as well as the reduction of the presence of several pro-inflammatory cytokines, such as TNF-α, IL-1β, IL-6, and IL-8, at brain level, by vanillic, gallic, and protocatechuic acids [126].
Flavanones
Flavanones are a flavonoid subclass mainly found in citrus fruits, and the most recurrent aglycones are hesperetin, naringenin, eriodictyol, isosakuranetin, and taxifolin, even if the major amounts of them are present as glycoside conjugated [133]. Several studies confirmed a low bioavailability of such compounds, with the identification and quantification of several colonic metabolites in biological fluids, such as phenylpropionic, phenylacetic, benzoic, and hydroxycarboxylic acids, and benzenetriol and benzoylglycine derivatives [134]. By considering blood oranges, mandarins, lime and lemon juice, the most representative compound in terms of amount in such fruits is hesperidin, which is the rutinoside of the hesperitin [133]. Hesperidin has been the object of several in vitro and in vivo animal studies to investigate its role as a neuroprotective compound. Concerning AD, the main results have been reported for the role of hesperidin in counteracting the oxidative stress at neuronal level, with improvement of the antioxidant endogenous mechanisms, such as counteracting glutathione depletion, and decrease of malondialdehyde and H2O2 accumulation, other than inhibiting some protein kinase phosphorylation leading to oxidative damage, in animal studies [135, 136]. Hesperidin at 100 mg/kg orally administered per 60 days was also found to be effective in the attenuation of the expression of amyloid-β precursors and oligomers in rats after aluminum chloride injection to mimic the damage caused by AD [137]. Several other molecular patterns involved in the boosting of neuroinflammation have been reduced in a mouse model affected by cognitive impairment when 100 and 200 mg/kg hesperidin has been supplemented for 15 days [138]. Anti-inflammatory and fibril anti-aggregative effects have also been observed in a recent study, where human neuroblastoma cells were incubated with up to 50μM liquiritigenin, mainly found in licorice [139]. To date, no human studies have considered the putative biological effects of the flavanones or their colonic metabolites on pathophysiological and clinical markers of AD, and this represents a promising area for future research.
Isoflavones
Among the so called “phytoestrogens”, isoflavones are a class of compounds mainly found in soy and its derivatives, genistein, daidzein, glycitein, formononetin, and their glycosides being the most representative compounds [108]. Studies on urine samples revealed a scarce bioavailability of such compounds, which are mainly metabolized by colonic bacteria in smaller compounds, such as equol, O-desmethylangolesin, and its 6’-hydroxylated form, which leads to 2-(4′-Hydroxyphenyl)propionic acid, and their conjugates [108]. There is solid evidence of neuroprotective effects of soy and soy-derived foods on several cognitive outcomes in animals and humans [140, 141]. Additionally, AD has been targeted in for studies focusing on isoflavones and, as previously detailed, the most relevant mechanisms highlighted in cell and animal models has been the counteraction of the amyloid-β deposition, oxidative damage, neuroinflammation, and apoptosis processes [142].
However, very few studies considered the colonic metabolites as real biological effectors towards the above-mentioned effects on the nervous system. In a randomized controlled trial, the administration of 100 mg/day of soy isoflavones to 65 older patients suffering from AD did not result in any significant improvement of cognitive function or dementia biomarkers after six months [143]. Researchers correctly argued that not all the individuals are equol-producers, mainly due to the gut microbiota composition and functionality, and so the missing effects of the isoflavone supplementation might be biased by the lack of consideration of this fact in the study design [143, 144]. Actually, when equol was administered to fifteen women with AD, the oxidative and inflammatory markers failed to be effectively modulated, but, clearly, further studies are needed to better understand this interesting aspect [145].
Flavonols
Flavonols are among the most ubiquitous polyphenols in fruit and vegetables, and quercetin, kaempferol, isorhamnetin, and myricetin are the principal representative aglycones, mainly conjugated in several different combinations with one or more sugar molecules [108]. Until recently, the major focus in biological fluids have been aglycones conjugated with glucuronide and/or sulfate moieties, mainly formed during the II detoxification at the upper gastro-intestinal and liver level. However, the simple fermentation of some compounds, such as quercetin, with fecal samples of healthy donors, revealed a deep rearrangement of its chemical structure, leading to the release of smaller phenolic acids, as phenylpropanoic, phenylacetic, and hydroxybenzoic acids [146]. A huge load of literature has been published regarding flavonols, and particularly quercetin, involvement in the processes leading to AD and, generally, cognitive disorders [147–148]. Results confirm a role of such compounds in the amelioration of oxidative status at neuronal level, inhibition of inflammatory and apoptotic key pathways, as well as amyloid-β oligomerization and deposition [147–149]. Also, for this class of compounds, colonic metabolites have not been deeply studied for their putative effects on cognitive decline markers.
Flavones
Regarding flavones, a flavonoid subclass widely present in vegetable-derived foods and beverages, the most studied compounds are apigenin and luteolin, which are usually present in foods as glycosides, which are barely absorbed along the small and large intestine and, above all, colonic bacteria seem to be unable to cleave the C-linked sugar moiety [108]. However, some data on protective effects of the in planta compounds towards AD-related markers, in cell and animal studies, have been reported for apigenin [150] and luteolin [151].
Gut microbiota involvement in polyphenol metabolism
As previously stated, the catabolism of phenolic compounds in the lower gastrointestinal tract is strictly dependent on the presence of certain bacteria strains, able to vastly modify the parent structures giving origin to smaller phenolic acids that are, in turn, absorbed quite efficiently. In a recent review, Cortés-Martín and collaborators summarized that, despite the many microbiological studies present in the literature, most of the microbial species responsible for specific metabolite production are still unidentified [152].
The hydrolysis of the sugar, in the case of the glycosylated polyphenols, is carried out by different species of Lactobacillus, Bifidobacterium, and Enterococcus, characterized by the activities of enzymes like glucosidases and rhamnosidases, able to cleave the O-sugar moiety (e.g., Lactobacillus plantarum or Bifidobacterium pseudocatenulatum) and, with a lower affinity, C-linkages (e.g., Enterococcus spp., Eubacterium cellulosolvens, and Lactococcus spp.) [152].
The elucidation of which species are able to cleave flavonoid rings is still open. Peiroten et al. summarized the main findings related to the metabolism of isoflavones by several different bacteria, identifying several species belonging to Clostridium and Eubacterium genera as able to transform daidzein into O-desmethylangolesin, and Bifidobacterium, Eggerthella, Lactococcus, and Slackia species as able to produce equol from daidzein [153]. A study from Germany identified two species, namely Eggerthella lenta and Flavonifractor plautii, in human fecal suspension, able to convert (–)-epicatechin and (+)-catechin into phenylpropanoic and phenylvaleric acids as well as to phenyl-γ-valerolactones [154]. Spanish researchers identified different species belonging to the Eggherthellaceae family (Gordonibacter urolithinfaciens and Gordonibacter pamelaeae) as able to convert ellagic acid into urolithins A and B and isourolithin A [155, 156].
The inter-individual variability role in the polyphenol gut microbial metabolism
Epidemiological studies have associated the intake of fruit, vegetables, and beverages rich in polyphenols with multiple beneficial effects on the human health. However, when intervention studies looked at the real biological effectors and at cause-effect relationships, a huge variability in terms of production/not production and, sometimes, concentrations of polyphenol gut microbial metabolites in fluids and tissues was observed. The inter-individual variability in the gut microbial capacity to produce peculiar profiles of phenolic metabolites seems to be the most important factor to explain the bioavailability of phenolic compounds and, possibly, the biological response to their intake [157]. The term “metaboytpe”—metabolic phenotype—has been introduced to indicate this characteristic excretion of metabolites after intake of specific dietary compounds [158]. Specifically, the new frontiers in this field will be the identification of cluster of individuals showing a metabotype associated with a well profiled gut ecosystem and, in turn, to a specific health status or improvement of health in response to food intake.
The definition of a metabotype should primarily include qualitative data, such as absence or presence of specific metabolites, but could also derive from quantitative aspects, i.e., relative ratios of metabolites [158]. The first pioneering studies in this field have been conducted on soy isoflavones, where the excretion of the urinary metabolites equol and O-desmethylangolesin have been observed only in certain individuals after soy consumption [159]. Equol producers have been shown to be characterized by more favorable hormonal profiles and by a lower breast cancer risk [160].
The research group led by Tomás-Barberán has shown that not only producers and not producers of urolithins, metabolites of dietary ellagitannins, are linked to specific enterotypes, but that producers can be further clusterized in metabotypes that are then associated to different presence of dysbiotic symptoms and also different cardiometabolic risk profiles [161]. Similarly, our group is working on the elucidation of the possible metabotypes following the ingestion of flavan-3-ol rich foods, with the preliminary findings indicating the presence of distinct groups of individuals excreting trihydroxyphenyl-γ-valerolactones, dihydroxyphenyl-γ-valerolactones, and hydroxyphenylpropionic acids [162, 163].
DIETARY FIBERS, GUT MICROBIOTA, AND SHORT-CHAIN FATTY ACIDS IN ALZHEIMER’S DISEASE
Dietary fibers include plant-based carbohydrates (non-starch polysaccharides, such as cellulose, resistant oligosaccharides, such as inulin, and resistant starch) that cannot be metabolized by human digestive enzymes [164]. Some gut bacteria are, however, able to metabolize such dietary compounds, generating SCFAs, i.e., acetate, propionate, and butyrate, as endproducts [165].
Most bacteria able to synthetize SCFAs belong to the Ruminococcaceae and Lachnospiraceae families [166, 167]. Few bacterial taxa, including Roseburia spp. and Eubacterium rectale, are able to complete the degradation of fibers to SCFAs alone [166, 167]. More frequently, there are cross-feeding interactions between taxa belonging to the same ecological niche: fibers are degraded to intermediate products by one group of bacteria, and then these products are utilized by SCFA producers [166, 167]. Alternatively, fiber degradation by one strain is essential for stimulating the growth and metabolic activity of a SCFA-producing strain [166]. The most known of these cross-feeding interactions regards Bifidobacterium spp. and Faecalibacterium prausnitzii, whose butyrate-producing capacity relies on the supply of acetate by bifidobacteria [168, 169].
SCFA synthesis by the gut microbiota is strongly influenced by the amount of fibers introduced with diet, that is able to shape the microbiota composition and enhance the SCFA metabolic pathways [170]. A recent population-based study has shown that the consumption of large amounts of fiber-rich plants is associated with increased representation of SCFA-producing genera in fecal microbiota [171]. A similar effect has also been demonstrated in an intervention study specifically focused on older subjects [172]. However, these effects do not necessarily correspond to an increase in SCFA production, due to the complex cross-feeding interactions among bacteria and variable expression of bacterial metabolic pathways [173].
Around 90% of SCFAs produced in the gut lumen are absorbed by colonocytes or enter circulation, while the remaining 10% is excreted in feces [166]. SCFAs exert a wide range of physiological functions in the intestine and in human metabolism, which have been extensively reviewed elsewhere [166, 174] and are summarized in Table 2.
Proposed physiological functions of short-chain fatty acids produced by gut microbiota in human beings
The studies investigating fecal microbiota composition in patients suffering from AD generally show a depletion SCFA-producing bacteria in comparison with controls (Table 3), especially taxa able to synthetize butyrate belonging to Lachnospiraceae or Ruminococcaceae [82–87]. However, butyrate producing bacteria can be isolated and cultivated from fecal samples of patients with AD [175] and a recent study failed to detect a significant difference in the relative abundance of the main SCFA producers between patients with AD and healthy controls (Table 3) [88]. Interestingly, a recent analysis of the gut microbiota composition and fecal metabolomics profile of 21 Chinese patients with AD and 44 controls with normal cognitive function revealed a paradoxical increase of representation of some SCFA-producing taxa, such as Ruminococcaceae and Faecalibacterium prausnitzii, in fecal samples from AD patients [89]. In the light of the existing literature, such findings suggest that the relationship between gut microbiota composition and cognitive dysfunction may be strongly influenced by external factors that were not considered as possible confounders, including dietary patterns and fiber intake. Geographical location of study participants may also represent an important issue: recent data indicate that study location represents the main factor explaining inter-individual variability in microbiome studies [176, 177].
Overview of the findings of studies that have investigated the potential of SCFA synthesis of the intestinal microbiota in patients with AD or other forms of dementia
AD, Alzheimer’s disease; MCI, Mild Cognitive Impairment; US, United States.
The gut mycobiome (i.e., ensemble of fungal populations symbiotically living with the host in the gut lumen) could also interact with the intestinal microbiome in patients with AD. A recent study has shown that reduced representation of SCFA producers, including Ruminococcus, Lachnospira, and Roseburia, is associated with a distinct intestinal mycobiome composition in patients with AD [178].
Overall, this evidence supports the assumption that patients with AD could have reduced SCFA synthesis in their gut microbiota. However, the des-criptive, cross-sectional design of these studies, and the absence of direct measurement of SCFA levels in feces or other biological samples does not allow to make any inference on the causal relationship between SCFA depletion and dementia. Furthermore, none of these studies considered nutritional intake of fibers as covariate.
In experimental models of AD, SCFAs exhibit a neuroprotective effect at multiple levels [179]. More specifically, they contribute to modulate the microglial cell function, reducing phagocytic activity, secretion of cytotoxins and synthesis of pro-inflammatory cytokines [179, 180]. In experimental conditions, SCFAs also inhibit amyloid-β aggregations, interfering with amyloid plaque formation and enlargement [181]. Interestingly, the brain amyloid load, measured with standardized uptake value ratio versus cerebellum through florbetapir amyloid-positron emission tomography (PET), was negatively correlated with blood butyrate levels in a group of 89 older subjects with different cognitive function, ranging from normal to overt dementia [182].
The clinical translation of these findings remains, however, uncertain. To date, studies specifically investigating the effects of increasing fiber intake on cognitive function and risk of dementia are lacking. Most studies were focused on nutritional patterns and frequency of consumption of fruit and vegetables, indicating that there could be an inverse relationship between fiber intake and cognitive performance, especially in those who already suffer from dementia or subjective cognitive complaints [24, 183–185]. Two studies also showed an inverse correlation between a dietary pattern characterized by elevated fiber intake and imaging biomarkers of AD (i.e., 18F-fluorodeoxyglucose uptake on brain PET in one study [186] and white matter hyperintensities module on brain magnetic resonance in another [187]). However, a large study conducted on 162 older subjects did not find any significant correlation between fiber intake and brain amyloid-β burden measured with amyloid-PET [188]. Furthermore, none of these studies considered investigation of SCFAs in their experimental design.
From a clinical perspective, the possibility of influencing AD pathophysiology through modulation of the intestinal microbiota towards an enhanced production of SCFAs is intriguing. However, very few clinical trials with probiotics or prebiotics have considered cognition as main endpoint [189]. Three recent systematic reviews and meta-analyses have shown contradicting results, supporting the plausibility of favorable effects of probiotics or prebiotics on cognition with a low level of evidence [94, 191]. None of the studied interventions were, however, specifically targeted at increasing SCFA production in the gut, and most studies were limited by small sample sizes.
Overall, the current evidence supports the concept that increasing dietary fiber intake is associated with favorable effects in the pathophysiology of AD through mediation of the gut microbiota. However, the clinical relevance of this mechanism and its therapeutic potential remains unclear.
CONCLUSIONS AND PERSPECTIVES
The current state of knowledge allows to hypothesize that the intestinal microbiome stands at the crossroads between nutrition and pathophysiology of AD, and that many of the putative beneficial effects of healthy dietary patterns on cognition could be mediated by the gut microbiota (Fig. 1). However, there is still a big gap between preclinical and clinical studies in this field. In fact, the influence of nutrition and gut microbiota on the pathophysiology and clinical course of AD has been studied mainly considering each player in a separate way, without integrating nutritional and microbiological investigations. The evidence suggesting favorable effects of gut microbiota-derived metabolites of polyphenols or fibers on markers of AD mostly comes from in vitro studies, and its translation in clinical practice is still difficult. As a result, there is a substantial lack of knowledge on how microbiome-centered nutritional interventions can modify cognitive function from a clinical perspective.
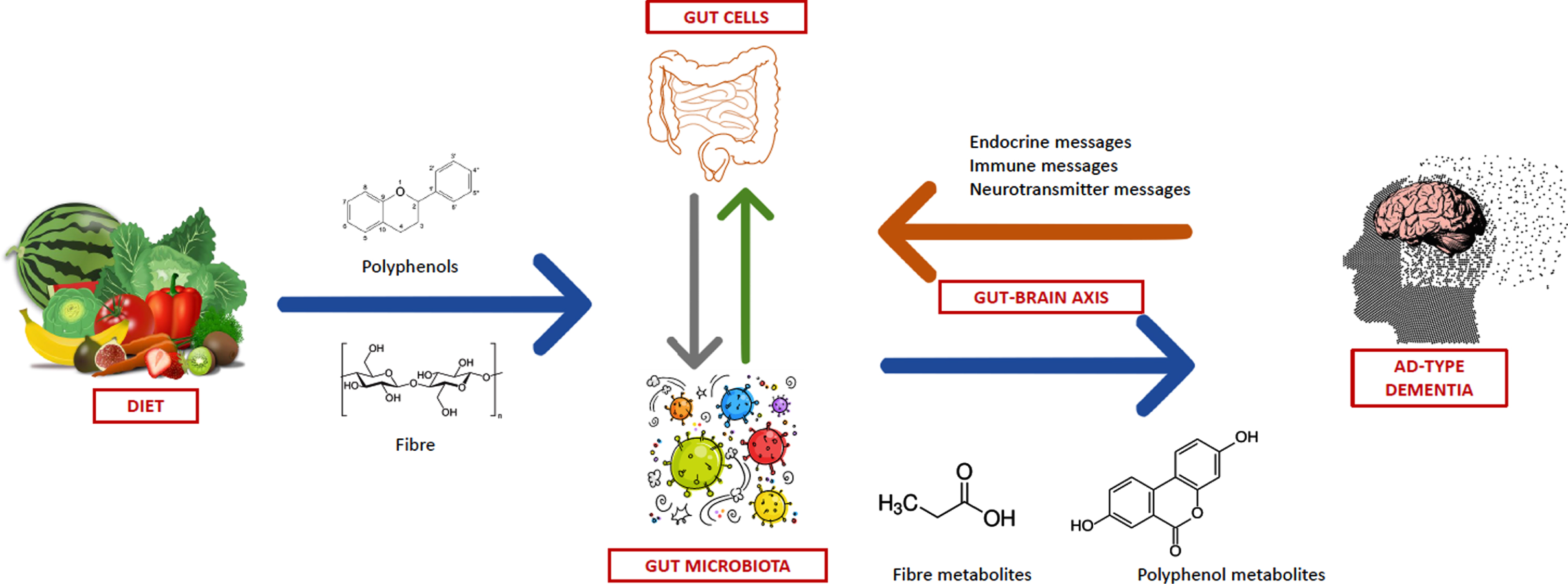
Overview of the possible link between dietary fiber and polyphenol intake, gut microbiota and modulation of the pathophysiology of Alzheimer’s disease.
However, the gut microbiota could become a reasonable target for anti-dementia interventions in the foreseeable future, as suggested by preclinical studies [80, 179]. Nutritional strategies implying an increase in the intake of polyphenols and fibers have a great potential of being effective in slowing down cognitive decline in older individuals, but future studies should also consider the influence of microbiota composition in this putative association. For example, the cognitive benefits of an increase in polyphenol intake could depend on specific metabotypes of the gut microbiota and the benefits, in terms of SCFA production, of increasing fiber intake could be influenced by the pre-existing network of strains with capacity of synthetizing butyrate [192, 193].
Future studies should also consider the co-occurrence of cognitive decline and physical frailty in older individuals [194]. Interestingly, gut microbiota dysbiosis is increasingly indicated as one of the contributors to the pathophysiology of physical frailty and sarcopenia [195, 196], concurring to define the health trajectory of older individuals [197]. Effective anti-aging microbiome-centered nutritional interventions should thus consider the link between microbiota and both cognitive and physical domains, translating the evidence coming from microbiological and animal studies into clinical practice.