
Abstract
Alzheimer’s disease (AD) is a progressive neurodegenerative disease with limited therapeutic strategies. NB-02 is a novel botanical drug that has shown promise as a protective and therapeutic treatment for AD in an APP/PS1 preclinical mouse model. In this paper, we investigate the underlying mechanisms by which NB-02 provides these therapeutic advantages using in vitro neuron-astrocyte co-cultures. Pretreatment with NB-02 prevented pathological calcium elevations in neurons and astrocytes after application of toxic soluble amyloid-β (Aβ) oligomers. NB-02 also prevented cell death associated with the addition of soluble Aβ oligomers suggesting NB-02 is effective at protecting both neurons and astrocytes from Aβ-mediated damage.
INTRODUCTION
Alzheimer’s disease (AD) is a progressive neurodegenerative disease with limited treatment options [1]. AD is characterized by amyloid beta plaques, neurofibrillary tangles, neuronal loss, and synapse dysfunction [2, 3]. Recently approved disease-modifying therapies present considerable risks [4–6].
NB-02 is a botanical therapeutic being developed by NeuroBo Pharmaceuticals. This multimodal drug, formerly DA-9803, contains extracts of Morus alba L. and Poria cocos [7, 8]. In vivo studies suggest NB-02 both prevents and slows AD progression in APP/PS1 mice [7, 8]. NB-02 treatment prevented Aβ plaque deposition, reduced rates of neuronal calcium overload, and induced morphological changes in astrocytes and microglia in young APP/PS1 mice [7]. In older APP/PS1 mice, NB-02 reduced Aβ plaque deposition and restored neuronal calcium levels [8]. We sought to explore the mechanisms by which NB-02 mediates these preventative effects using primary cortical neuron-astrocyte co-cultures.
Soluble Aβ oligomers are emerging as a particularly toxic form of Aβ [9]. We found that NB-02 treatment prevented Aβ-mediated calcium overload in both neurons and astrocytes in culture after application of transgenic conditioned media (TgCM) enriched with small bioactive soluble Aβ oligomers [10, 11]. NB-02 also improved cell viability after TgCM application. Overall, these results extend our previous findings and recapitulate the protective effects of NB-02 against Aβ-mediated damage.
MATERIALS AND METHODS
Primary neuron-astrocyte co-cultures
All animal procedures were performed in accordance with the Massachusetts General Hospital animal care committee’s regulations. Primary neuron-astrocyte co-cultures were prepared from E13-16 CD1 mouse embryos (Charles River Laboratories) as described elsewhere [12]. The cortices were dissociated using a Papain Dissociation System (Worthington Biochemical Corporation) following the manufacturer’s protocol. Cells maintained in neurobasal medium with 2% B27 supplement, 2 mM Glutamax, 100 U/mL penicillin, and 100 g/mL streptomycin at 37°C with 5% CO2 for 10–18 days in vitro (DIV). The neuron-astrocyte co-cultures were mainly comprised of neurons (93.7%, Supplementary Figure 1B) with a small percentage of astrocytes and no identifiable microglia (Supplementary Figure 1A).
Preparation of transgenic conditioned media (TgCM) and wildtype conditioned media (WtM)
Primary neuronal cultures were prepared from E13–E15 Tg2576 transgenic or nontransgenic mice acquired from Taconic using the method described previously [10] and maintained in culture for 14DIV. The culture media enriched with Aβ oligomers bathing the cells was collected and considered TgCM if derived from Tg2576 cultures or WtM if derived from nontransgenic cultures. The levels of Aβ40 in the TgCM and WtM were calculated using an Aβ1–40 human/rat sandwich ELISA kit (Wako Pure Chemicals Industries, Japan #294-64701). The average concentration of Aβ40 was 10 nM in TgCM and 0.33 nM in WtM. The levels of Aβ42 in the TgCM and WtM were calculated using a human/rat Aβ42 ELISA kit, high sensitivity (Wako Pure Chemicals Industries, Japan #292-64501). The average concentrations of Aβ42 were 0.8 nM in TgCM and negligible amounts in WtM.
Multiphoton imaging of live cells
10–18 DIV cultures were incubated with 6μM indo-1/AM dye, a cell permeant ratiometric calcium sensor as previously described [10] and 2μM sulforhodamine 101 (SR101), a dye that labels astrocytes, for 45 min at 37°C. Cells were imaged on an inverted Zeiss LSM 510 multiphoton live imaging system, while maintained at 37°C in a humidified environmental chamber with 5% CO2:O2. Multiphoton excitation of Indo-1 was performed as previously described [10] and SR101 was imaged using 543 nm excitation with a 565–615 IR emission filter. During imaging, 4–10 fields per well were randomly acquired that included ∼300–400 neurons and ∼50–100 astrocytes in 5–6 wells per condition.
Image processing and analysis
Image analysis to measure the ratio of fluorescence intensity of bound calcium to unbound calcium was done as described elsewhere [10]. Images were pseudocolored according to calcium concentrations derived from the ratios. Calcium overload was defined as an Indo-1 ratio of two standard deviations above the mean Indo-1 ratio in the baseline condition.
Therapeutic treatment
Prior to baseline imaging, primary 10-18DIV co-cultures were pretreated with 300μg/mL NB-02 or Vehicle for 45 min. The Vehicle consisted of hydroxypropyl methyl cellulose (HPMC) dissolved in neurobasal medium with 2% B27 supplement, 2 mM Glutamax, 100 U/mL penicillin, and 100 g/mL streptomycin. NB-02 consisted of NB-02 mixed with the Vehicle solution. One batch of NB-02 was used for all experiments.
Cell viability assay
The LIVE/DEAD Viability/Cytotoxicity Kit (ThermoFisher #L3224) was used according to the manufacturer’s instructions. 10-14DIV co-cultures processed for the LIVE/DEAD assay were imaged on an inverted Olympus confocal microscope. Live cells were labeled with calcein-AM, with emission at 494 nm, while dead cells were labeled with ethidium homodimer-1, with emission at 528 nm. 4–10 fields per well were acquired, with ∼300 cells in 5–6 wells per condition. Live and dead cells were manually counted to obtain the cell viability (live cells/total cells).
Statistical analysis
Graphpad Prism and Microsoft Excel were used to perform statistical analyses. The Shapiro-Wilk test was used to determine normality. For statistical analysis of calcium overload, a mixed effect model was used. For analysis of cell viability, the Kruskal-Wallis, or one-way ANOVA was used to test for statistical significance. Afterwards, we used Dunn’s Multiple Comparison Test to determine which experimental conditions were significantly different from each other.
RESULTS
NB-02 prevents Aβ-mediated calcium overload in neurons
Primary neuron-astrocyte co-cultures were used to assess the effect of soluble Aβ oligomers on cytosolic calcium. At 10-18DIV, Indo-1/AM was added to measure intracellular calcium concentrations ([Ca2+]) in neurons and astrocytes, and SR101 to label astrocytes. Neuronal calcium was analyzed by identifying cell bodies that were positive for Indo-1 but not SR101. Cultures primarily consisted of neurons, 93.7% were Map2+ (Supplementary Figure 1A, B). Cells were treated with NB-02, Vehicle, or No Drug for 45 min and imaged at baseline (Fig. 1A). A small percentage of neurons exhibiting calcium overload was observed at baseline (4.0±0.1% Fig. 1B), consistent with past work [10]. 150μl out of a total of 300μl of neurobasal media was replaced in each well with either TgCM, WtM or Neurobasal Media for 1 h and followed by a second round of imaging (Fig. 1A). Only half of the media was changed to prevent disruption to the primary cells and to decrease TgCM toxicity. TgCM application significantly increased the percentage of neurons exhibiting calcium overload (17.5±1.8% p = 0.0011; Fig. 1C) [11, 12]. Critically, pretreatment with NB-02 prevented TgCM-mediated increases in neuronal calcium overload (4.1±0.4% Fig. 1F); however, pretreatment with vehicle failed to do so (11.8±0.9% p = 0.0161; Fig. 1I). As a control, WtM, which contains 0.33 nM non-toxic mouse Aβ40, was tested on untreated cultures. Neuronal calcium overload after WtM application was comparable to baseline (3.5±0.6% Fig. 1D). Pretreatment with NB-02 maintained low levels of calcium overload after WtM application (4.9±0.7% Fig. 1 G), similar to pretreatment with vehicle (2.0±0.4% Fig. 1J). NB-02, on its own, did not significantly change neuronal calcium overload rates (3.6±0.8% Fig. 1E). Vehicle treatment did not change intracellular calcium in the absence of conditioned media (4.1±1.2% Fig. 1 H). Thus, TgCM elicited neuronal calcium overload in neuron-astrocyte co-cultures (Fig. 1K), while neuronal calcium overload rates remained low after application of WtM (Fig. 1K). TgCM contains human and mouse Aβ oligomers while WtM contains only mouse Aβ oligomers, suggesting that these calcium elevations are a result of the presence of human Aβ in the TgCM. Importantly, pre-treatment with NB-02 exclusively prevented significant elevations in neuronal calcium when challenged with TgCM (Fig. 1K). Furthermore, there was not a significant effect of WtM and NB-02 when administered independently or together on neuronal calcium elevations (Fig. 1K). This work suggests that NB-02 reduces the number of neurons with Aβ oligomer-dependent elevations in calcium.
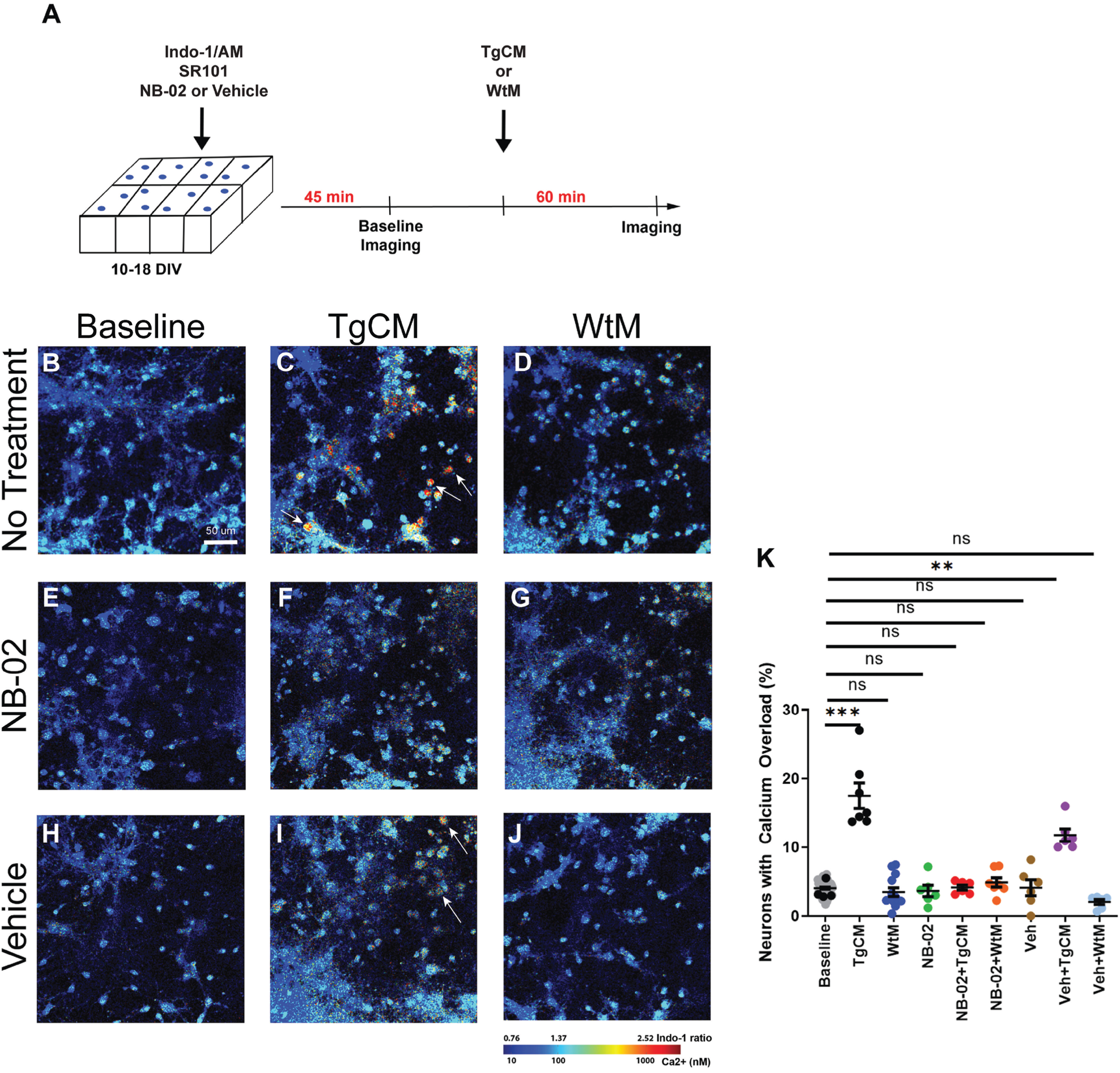
NB-02 prevents Aβ-mediated calcium overload in neurons. A) Experimental setup showing addition of NB-02 or Vehicle concurrently with SR101 and Indo-1 45 min before baseline imaging. TgCM or WtM was then added for 60 min before cells were re-imaged. B–J) Multiphoton microscopy images of neuron-astrocyte co-cultures at baseline (B), after addition of TgCM (C) or WtM (D). Representative multiphoton microscopy images of cells pretreated with NB-02 at baseline (E), following the addition of TgCM (F) or WtM (G). Representative multiphoton microscopy images of Vehicle-treated cells at baseline (H), after addition of TgCM (I) or WtM (J). K) The percentage of neurons that exhibited calcium overload was calculated for each condition (K). Arrows in (C) and (I) show neurons exhibiting calcium overload. Mean±SEM, N = 300 neurons/well in 5–6 wells. Mixed Effect Model used to test for statistical significance, *p < 0.05, **p < 0.01, ***p < 0.001; n.s, not significant. Scale bar: 50μm.
NB-02 prevents Aβ-mediated calcium overload in astrocytes
To explore NB-02’s non-neuronal effects, we examined how NB-02 affected calcium overload levels in astrocytes when similarly challenged by Aβ. Astrocytic cell bodies were identified from images by selecting cells that were positive for both Indo-1 (Fig. 2A) and SR101 (Fig. 2B). Cells exhibiting co-localization of Indo-1 and SR101 (Fig. 1C) were labeled as astrocytes and analyzed to determine the percentage of cells that exhibited calcium overload. Although the primary cultures were neuron-rich, a small percentage of astrocytes were identified (Fig. 1C, Supplementary Figure 1A). At baseline, only 4.5±0.1% of astrocytes exhibited calcium overload (Fig. 2D) [13]. A significant increase in calcium overload (13.4±1.0% p = 0.0150; Fig. 2E) was observed after 150μl out of the 300μl total neurobasal media was replaced with TgCM. Pretreatment with NB-02 prevented TgCM-mediated elevations in astrocytic calcium overload (3.7±0.8% Fig. 2H), while vehicle failed to do so (13.1±0.8%, p = 0.0217; Fig. 2K). As expected, WtM addition to cultures did not significantly increase calcium overload (4.0±1.0% Fig. 2F), neither did pretreatment with NB-02 (3.5±0.9% Fig. 2I) nor Vehicle (4.6±0.7% Fig. 2L). Addition of NB-02 alone did not result in significant changes of astrocytic calcium overload from baseline (2.9±1.1% Fig. 2G). The observed calcium overload following pretreatment with Vehicle was comparable to baseline (4.2±1.1% Fig. 2J). Thus, addition of TgCM elicited calcium elevations in astrocytes (Fig. 2M). Similar to NB-02’s protective effect on neurons, pretreatment of cultures with NB-02 exclusively prevented significant calcium elevations after the addition of TgCM (Fig. 2M).
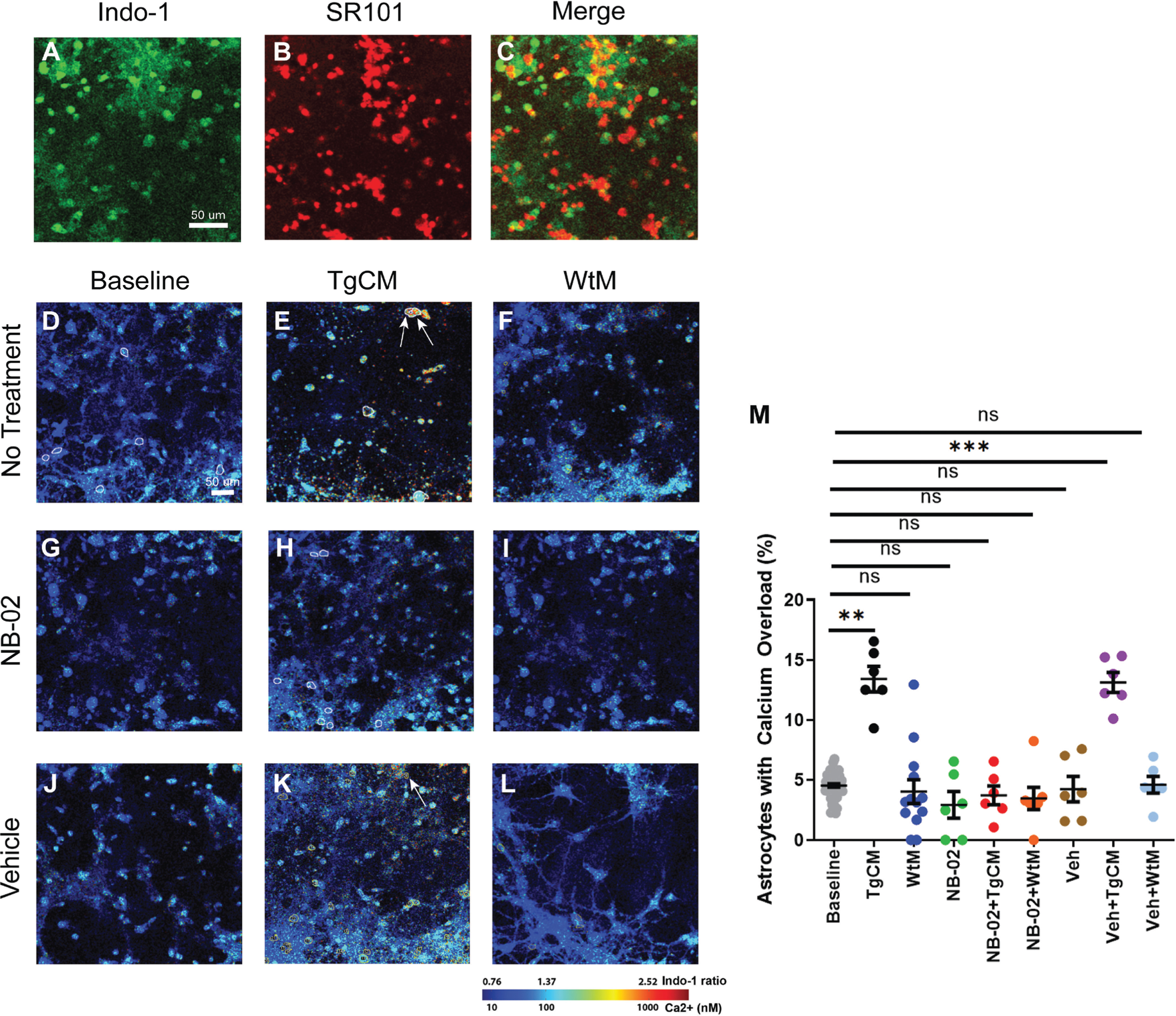
NB-02 prevents Aβ-mediated calcium overload in astrocytes. Multiphoton microscopy images of Indo-1 (A) and SR101 (B) and their co-localization (C) used to distinguish astrocytes and neurons. D–L) Multiphoton microscopy images of neuron-astrocyte co-cultures at baseline (D), after addition of TgCM (E) or WtM (F). Representative multiphoton microscopy images of cells pretreated with NB-02 at baseline (G), following the addition of TgCM (H) or WtM (I). Representative multiphoton microscopy images of Vehicle-treated cells at baseline (J), after addition of TgCM (K) or WtM (L). M) The percentage of astrocytes that exhibited calcium overload was calculated for each condition (M). Arrows in (E) and (K) show astrocytes exhibiting calcium overload. Mean±SEM, N = 300 astrocytes/well in 5–6 wells. Mixed Effect Model used to test for statistical significance, *p < 0.05, **p < 0.01, ***p < 0.001; n.s, not significant. Scale bar: 50μm.
NB-02 prevents Aβ-mediated cell death in neurons
We evaluated the effect of NB-02 on cell viability in the presence of human Aβ oligomers. Cell viability was 0.87±0.01 for cultures maintained in neurobasal media in absence of treatment (Fig. 3A). The cell viability was significantly decreased after the replacement of all 600μl neurobasal media with TgCM (0.09±0.07; p = 0.0003; Fig 3B). Pretreatment with NB-02 prevented the decrease in cell viability associated with the addition of TgCM, maintaining cell viability comparable to that of untreated co-cultures (0.68±0.02; Fig. 3E). However, pretreatment with Vehicle resulted in significant decreases in cell viability after application of TgCM (0.15±0.08; p = 0.0016.; Fig. 3H). When 600μl of WtM was added to co-cultures, cell viability did not significantly differ from that of untreated cells (0.77±0.02; Fig. 3C). Addition of WtM to NB-02-pretreated cultures maintained cell viability comparable to baseline (0.73±0.02; Fig. 3F). Pretreatment with vehicle also maintained cell viability after incubation with WtM (0.84±0.02; Fig. 3I). Neither NB-02 nor vehicle significantly altered cell viability compared to baseline in cultures lacking TgCM and WtM (0.84±0.02; Fig. 3D, 0.84±0.02; Fig. 3G). Therefore, the addition of TgCM, containing human oligomeric Aβ, significantly decreased cell viability (Fig. 3J), except when pre-treated by NB-02.
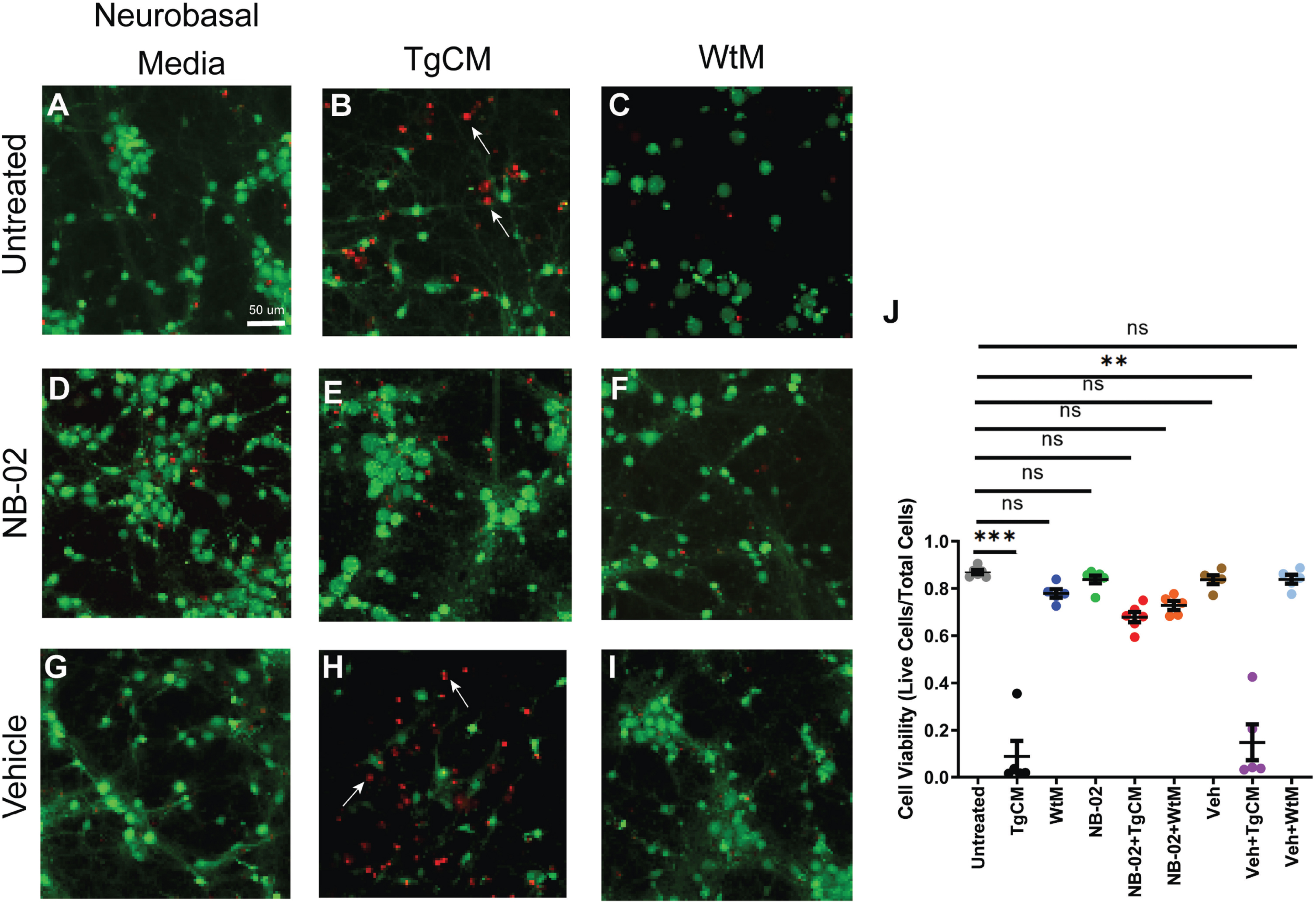
NB-02 prevents Aβ-mediated cell death in neurons. A–I) Confocal microscopy images acquired during the LIVE/DEAD assay showing live cells stained with calcein-AM (green) and dead cells stained with ethidium homodimer-1 (red) in neurobasal media in absence of treatment (A), after incubation with TgCM (B) or WtM (C). Representative images acquired after pretreatment with NB-02 in absence of (D), or presence of TgCM (E) or WtM (F). Representative images acquired after pretreatment with Vehicle in absence of (G), or presence of TgCM (H) or WtM (I). J) Cell viability across condition. Arrows in (B) and (H) indicate dead cells. N = 300 cells/well in 5–6 wells. Mean±SEM. Kruskal-Wallis used to test for statistical significance, **p < 0.01, ***p < 0.001; n.s, not significant. Scale bar: 50μm.
DISCUSSION
NB-02 has been shown to have both preventative and therapeutic effects in vivo in APP/PS1 mice [7, 8]. In this study, we find evidence that NB-02’s salubrious effects may be due to NB-02’s protection against oligomeric Aβ-mediated damage. Neurons and astrocytes normally exhibit pathological calcium elevations with the addition of human Aβ oligomers present in TgCM. However, pre-treatment with NB-02 prevented neuronal/astrocytic calcium overload as well as increased neuronal viability when challenged with human oligomeric Aβ.
The calcium hypothesis of AD argues that calcium dyshomeostasis leads to downstream biochemical cascades resulting in the progression of AD [14, 15]. We reproduce results from previous groups, Alberdi et al., 2010 and Sanz-Blasco et al., 2008, showing that human Aβ oligomers present in TgCM lead to cell death in culture [11, 16]. By maintaining cell viability and reducing rates of calcium overload in both neurons and astrocytes, NB-02’s therapeutic effects are evident. The activation of NMDA receptors has been shown responsible for calcium overload, possibly suggesting a method through which NB-02 protects neurons and astrocytes from Aβ-mediated damage [10]. Further investigation is necessary to understand whether NB-02 modulates NMDA receptors and precisely how NB-02’s protective effects are mediated.
AUTHOR CONTRIBUTIONS
Morgan R. Miller (Conceptualization; Data curation; Formal analysis; Methodology; Project administration; Writing – original draft; Writing – review & editing); Lavender Lariviere (Conceptualization; Data curation; Formal analysis; Methodology; Writing – original draft); Guillaume J Pagnier (Conceptualization; Methodology; Writing – original draft; Writing – review & editing); Sema Aygar (Data curation); Natalia Wieckiewicz (Data curation); Masato Maesako (Data curation); Brian J Bacskai (Conceptualization; Funding acquisition; Project administration; Resources); Ksenia V. Kastanenka (Conceptualization; Funding acquisition; Investigation; Methodology; Project administration; Resources; Supervision; Writing – original draft; Writing – review & editing).
Footnotes
ACKNOWLEDGMENTS
The authors have no acknowledgements to report.
FUNDING
The work was supported by NeuroBo Pharmaceuticals, the BrightFocus Foundation Grant A2020833 S and the National Institute of Health Grant R01AG066171.
CONFLICT OF INTEREST
This work was funded by NeuroBo Pharmaceuticals.
DATA AVAILABILITY
The data supporting the findings of this study are available on reasonable request from the corresponding author.