
Abstract
BACKGROUND:
Blueberry juice is a common drink for relieving fatigue, but its mechanisms of action are unclear.
OBJECTIVE:
This study aimed to validate changes in the dopaminergic system, especially changes in dopamine transporter (DAT) levels, in a fatigued state and assess whether such fatigue-related changes can be ameliorated by blueberry juice.
METHODS:
We investigated the expression of striatal DAT and tyrosine hydroxylase (TH) in mice subjected to a forced swimming fatigue task and subsequently administered blueberry juice (anthocyanin), caffeine, or Red Bull (caffeine + taurine) to assess DAT and TH changes in the nigrostriatal pathway related with fatigue as well as their relief.
RESULTS:
Striatal DAT and TH expression was downregulated by fatigue and upregulated by treatment with blueberry juice, caffeine, and Red Bull. In addition, DAT and TH expression exhibited positive correlations with fatigue-related motor performance.
CONCLUSIONS:
We provided evidence of compensatory responses of striatal DAT and TH in the fatigued state, and treatments to relieve fatigue might upregulate the expression of both striatal DAT and TH. Moreover, small amounts of blueberry juice exerted similar effects as large amounts, suggesting that anthocyanin has relatively strong anti-fatigue effects.
Keywords
Introduction
Fatigue is a familiar but subjective feeling of a lack of energy or tiredness that can be caused by several factors such as staying up late and job stress. As relieving fatigue is essential for enhancing quality of life, several interventions, including dietary therapy, are under investigation. However, the mechanisms of fatigue have been only partially elucidated. Reportedly, fatigue is a self-protective inhibition that aims to prevent defatigation [1], and it is associated with the limbic input in the basal ganglia related to dopaminergic neurons [2]. Several studies suggested that reduced dopamine (DA) levels [3, 4] or downregulation of DA production [5] triggers the onset of fatigue, whereas enhancement of cerebral DA content relieves fatigue. Another possible factor associated with fatigue is the dopamine transporter (DAT), which mediates the reuptake of DA from the extracellular space to presynaptic neurons. A prior study reported that animals that underwent treadmill training exhibited downregulation of DAT mRNA expression, whereas striatal DA levels were increased [6]. Foley et al. highlighted that the downregulation of DAT (induced by physical training) might prolong the elevation of DA levels in the nigrostriatal pathway by reducing DA reuptake, which delays fatigue [7]. However, in the fatigued state, no changes in DAT levels have been observed directly. In our opinion, DAT might be downregulated with reduced DA levels in the striatum, which is a compensatory response to fatigue.
Anthocyanin is a vital bioactive component of many fruits. A study using mulberry reported that anthocyanin possesses anti-fatigue effects [8]. However, the mechanisms involved in the anti-fatigue effects of blueberry remain unclear. We speculate that the dopaminergic system might play a role in the anti-fatigue effects of blueberry.
The objectives of this study were two-fold. First, the study aimed to verify the changes in the dopaminergic system, especially of DAT levels, in the fatigued state. Tyrosine hydroxylase (TH), the enzyme associated with DA synthesis, has been used as a marker of dopaminergic neurons. Although there is no direct evidence linking TH expression with fatigue, it has been reported that TH levels are highly associated with DA levels in the brain [9, 10], which are also closely related to fatigue [11]. Some reports indicated that phosphorylated TH is more active than TH [12], but later reports regarded TH as the gold standard marker for identifying DA neurons [13]; hence, we also selected TH as an index of the cerebral dopaminergic state in this study. Second, the study sought to elucidate whether such fatigue-related changes can be ameliorated by blueberry juice, which is readily available. We hope that the findings of this study provide evidence of changes of DAT levels in the simple fatigued state and offer a method to for relieving fatigue, namely drinking blueberry juice.
Materials and methods
Experimental design
In this study, we used 42 male Kunming mice (body weight, 20±2 g) obtained from the Experimental Animal Center of the Dalian Medical University (Dalian, China). All animals were divided into six groups (n = 7/group) as follows: Cont (intact control, untreated mice); F (fatigue); RB (Red Bull, 20 mL/kg; RedBull China Co. Ltd. Beijing China); C (caffeine, 5 mg/kg; Nanjing Jiancheng Bioengineering Institute, Nanjing, China); BL (low-concentration blueberry juice, 5 mL/kg); and BH (high-concentration blueberry juice, 20 mL/kg). All animals were treated according to the National Institute of Health Guidelines for the Care and Use of Laboratory Animals, and all experimental processes were approved and supervised by the Animal Care and Use Committee of Dalian Medical University (authorization No: 85–23). We housed all mice under controlled temperature (22±2°C), humidity (50% ±5%), and light conditions (lights on at 08:00 and off at 20:00), with food and water made available ad libitum.
In our experiments, we used a forced swimming fatigue mouse model to elucidate the anti-fatigue effects of blueberry juice. We also compared the anti-fatigue effects of Red Bull (a popular energy drink to relieve fatigue containing caffeine, taurine, and B vitamins) and caffeine (a nervous system stimulant with anti-fatigue effects) with those of blueberry juice at low and high concentrations. We used the same Red Bull formulation as a previous study [14], which included caffeine 80 mg, taurine 1000 mg, B group vitamins (B3, B5, B6, B12), glucose 27 g, and alpine spring water per 250 mL [14]. The concentration of Red Bull was decided by our preliminary experiments and a previous study [15] using Red Bull in mice. The caffeine solution was prepared, and the concentration (5 mg/kg) was selected according to a previous study [16] in consideration of the dose in humans (recommended maximum amount of 400 mg/day [17]). The concentration of blueberry juice was decided by our preliminary experiments considering the tolerance capacity of mice. In addition, we established the Cont and F groups for comparison. Mice were forced to swim (60 min/day) for 5 consecutive weeks. From the fourth week, swimming was performed following the intragastric administration of blueberry juice or other solutions. To confirm the fatigued state, animals were heavily anesthetized (chloral hydrate, 400 mg/kg), and blood samples were obtained to evaluate the concentrations of blood lactic acid (BLA), blood urea nitrogen (BUN), and glycogen (liver glycogen [LG] and muscle glycogen [MG]) as fatigue indices [18], which were measured according to the instructions provided with the biochemical kits (Liver/Muscle glycogen assay kit, Jiancheng Bioengineering Institute, Nanjing, China). Then, the brains of the mice were removed for immunohistochemical staining to investigate striatal TH and DAT expression. The experimental design is shown in Fig. 1.
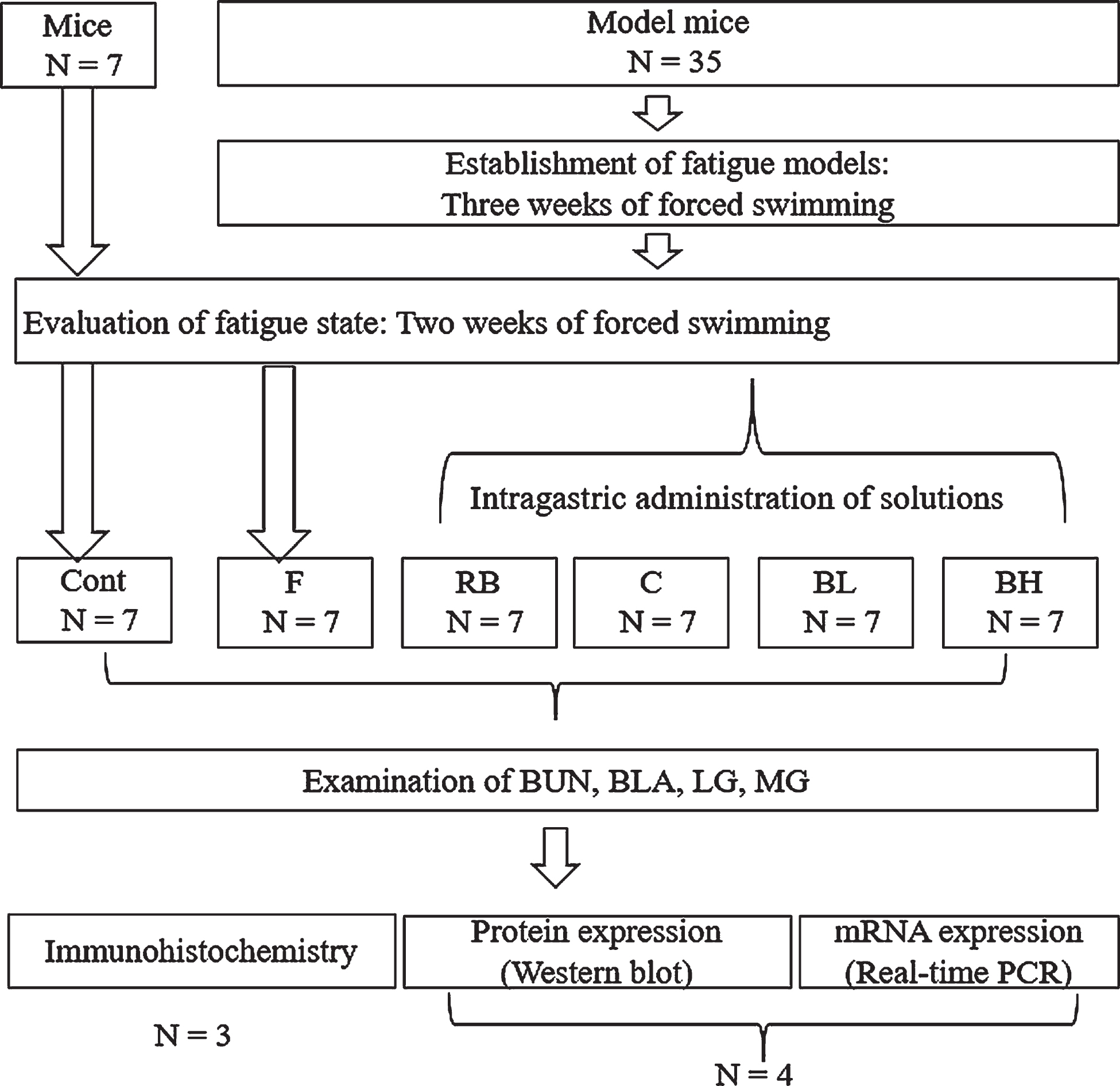
Experimental design of the present study. Cont = intact control group; F = fatigue group; RB = Red Bull group; C = caffeine group; BL = low-concentration blueberry juice group; BH = high-concentration blueberry juice group.
We treated fresh blueberries (Dalian Limited Company of Blueberry Development, Dalian, China) using freeze concentration technology [19, 20]. Briefly, 5 kg of frozen blueberry fruits were manually crushed and stored at – 20°C overnight. After freezing the solution, 50% of the initial volume was thawed and separated. The same procedure was repeated for the next cryoconcentration cycle until the final volume of blueberry juice reached 312 mL. Of note, all procedures were performed away from light.
Ingredient analysis of the samples
The ingredients of the samples of fresh blueberry juice and fruit were analyzed using a highly sensitive and specific liquid chromatography-tandem mass spectrometry method described in previous studies [21, 22]. Briefly, samples were analyzed using an Agilent 1100 (Agilent, Santa, CA, USA) HPLC system with a small molecule chip cube interface and Agilent 6500 Q-TOF MS/MS (Agilent, USA). A C18 (4.6×150 mm, 5μm, Kromasil, Sweden) column was used for detection. We used 0.1% formic acid (A) and acetonitrile (B) as the mobile phase. We held the gradient as follows: 0–5 min, 20% B; 5–10 min, 20–25% B; 20–25 min, 25–30% B; and 35–40 min, 30–100% B. We set the VCap at 3500 V and the fragmentor at 175 V. The capillary temperature was set at 350°C using N2 as the carrier gas (flow rate of 13 L/min). The MS scan rate was 1 scan per second. Each compound level was calculated using the semi-quantified peak area abundance identified with the characteristic compound parent fragment molecular weight and the fragmented MS/MS backbone molecular weight. We measured the compounds in samples using averaged ion counts.
The ingredients of the blueberry juice and fruit are shown in Table 1. Our data revealed that the main active ingredient was anthocyanin, and no caffeine or taurine was detected in the samples, in accordance with previous studies [23–25]. Hence, we evaluated the concentration of anthocyanin to confirm the bioactivity of blueberry juice. The data suggested good bioactivity for the juice and verified that the sample was appropriate for further experiments (Table 1) [26].
Major ingredients of the blueberry samples used in the present study measured using LC-MS/MS ingredient analysis
Major ingredients of the blueberry samples used in the present study measured using LC-MS/MS ingredient analysis
The data are presented as the mean±standard deviation.
The forced swimming test is usually used for testing depression-like behavior in rodents [27]. However, it is also an essential behavioral test for fatigue [5]. However, the index for fatigue is the “exhaust time” (time of swimming), or the duration of the entire swimming process, whereas that for depression is the “duration of immobilization” (time of NOT swimming) [27, 28].
As described previously [29], after the last intragastric administration, all animals were forced to swim individually with a load (∼5% of their body weight) in a 30 cm deep cylindrical container filled with water (23±2°C). They were forced to swim until they could not return to the water surface within 10 s. We performed the experiments at 3:00 pm for the 5 consecutive weeks. The first 3 weeks were used to establish the fatigue model, and the next 2 weeks were used to evaluate the fatigued state (Fig. 1). Each day, we allotted the animals one practice swim and then performed the examination twice with a 30 min interval. We recorded the time of the entire process as “the exhaust time” as an index of the forced swimming test.
Immunohistochemical staining
After the mice were heavily anesthetized (chloral hydrate, 400 mg/kg), their brains were removed and post-fixed in 4% paraformaldehyde, followed by cutting the coronal sections on a vibratome (Kedee KD-202, China) and examination under a light microscope (Olympus, Japan). Then, we selected sections from the striatum of each mouse. Striatal samples from three mice in each group were used to assess TH and DAT expression via immunohistochemical staining. Anti-TH (1:1000; SAB2103911; Sigma) and anti-DAT (1:200; D6944; Sigma) primary antibodies and Alexa Fluor 488-conjugated goat anti-rabbit secondary antibody (1:500; D111101; Sangon Biotech) were used for immunohistochemistry. Furthermore, all sections were examined using fluorescence microscopy (Olympus IX-71; Olympus).
Western blot analysis of TH and DAT protein expression in the striatum
We used a standard process for Western blotting to evaluate striatal TH and DAT protein expression. We homogenized a part of the striatum samples (4 mice/group) in radioimmunoprecipitation assay lysis buffer containing protease and phosphatase inhibitor cocktails followed by centrifugation. Then, we separated the protein extracts in a 12% SDS–PAGE gel and transferred them to a PVDF membrane. The membrane was then blocked with 5% non-fat dry milk in TBS containing 1% Tween-20 and 5% milk, followed by incubation with the primary antibody (Anti-Dopamine Transporter [catalog number ab5990], Anti-Tyrosine Hydroxylase [catalog number ab75875]), Dingguo, Beijing, China) and HRP-conjugated goat anti-rabbit secondary antibody (1:10000; D110058; Sangon Biotech, Shanghai, China). Finally, the immunoreactive bands in the membrane were visualized using ECL reagent.
Reverse transcription and real-time PCR analysis of TH and DAT mRNA expression
The remaining striatal tissue of four mice in each group was used to obtain mRNA. We performed real-time PCR using cDNA to measure TH and DAT mRNA expression relative to that of the housekeeping gene β-actin using the following primers: TH forward, 5′-CAA TGA TGG CAG AGG TGT GG-3′; TH reverse, 5′-CTC GTG GCA GCA GTC TCT GT-3′; DAT forward, 5′-GAA TGG AGT GCA GCT GAC CA-3′; DAT reverse, 5′-GAA GCC GAT GAC TGA GAG CA-3′; β-actin forward, 5′-CCA GAG AGC TCA CCA TTC ACC-3′; and β-actin reverse, 5′-CAT CTG CTG GAA GGT GGA CA-3′. These primers were designed using primer 5.0. In this study, 40 cycles were used for TH, DAT, and β-actin amplification. All samples were run in triplicate, and data were analyzed using the 2–ΔΔCt method [30].
Statistical analysis
All statistical analyses in this study were performed using SPSS software (V20.0.0; IBM, IL). We used two-way analysis of variance followed by Bonferroni’s post-hoc correction for multiple comparisons to determine differences among the groups. By contrast, the Student’s t-test was used to compare the difference between groups administered low and high concentrations of blueberry juice. All data are presented as the mean±standard deviation, and we considered P < 0.05 to indicate statistical significance.
Results
Figure 2 shows the effects of blueberry juice on the indices associated with fatigue. First, all indices in the fatigue group were worse than those in the control group, suggesting the successful establishment of a mouse fatigue model. We observed that all interventions significantly ameliorated the changes in indices induces by the forced swimming task. Remarkably, we did not observe any difference in effects between the low and high concentrations of blueberry juice, which suggested the high activity of blueberry juice.
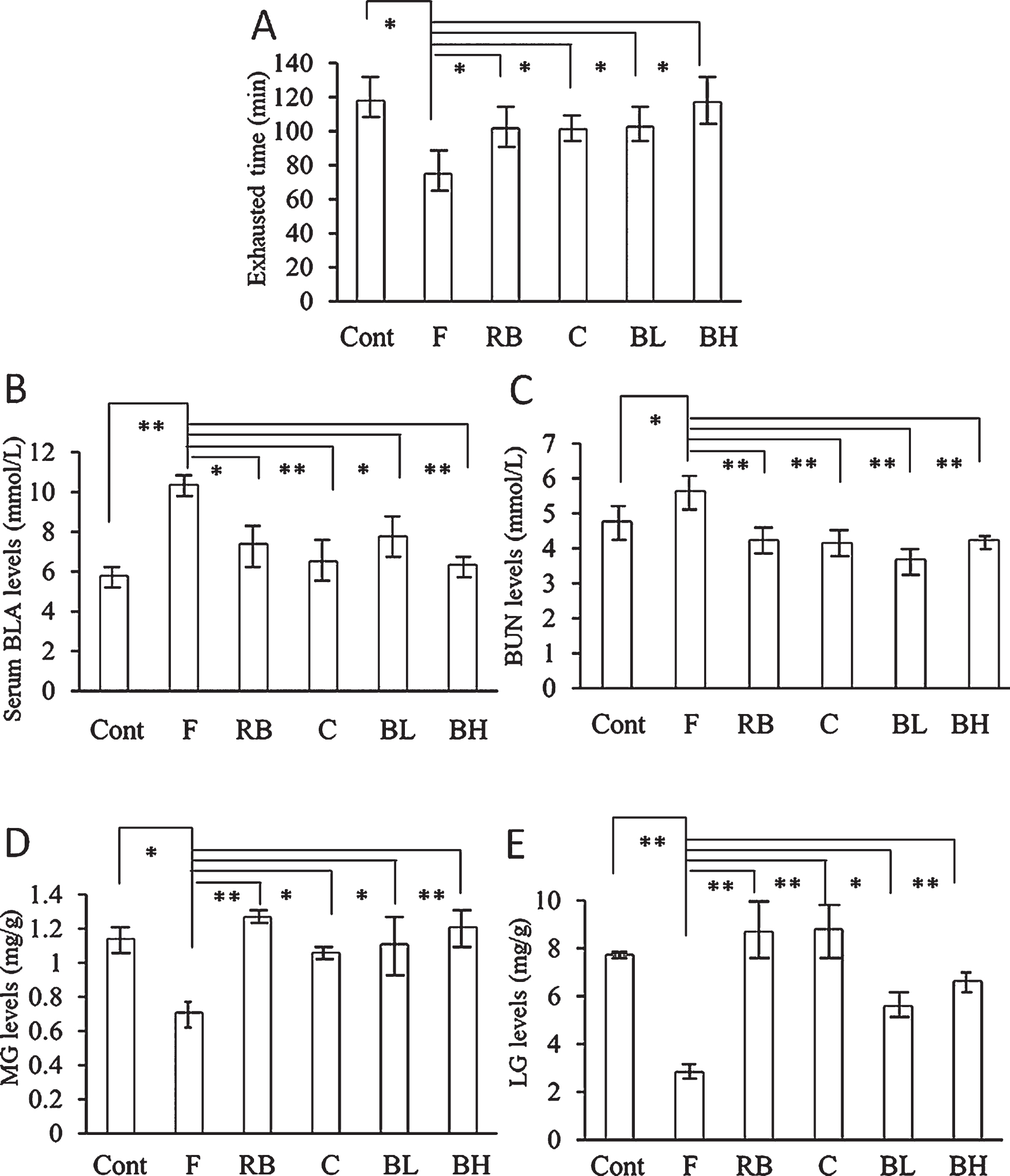
Effects of blueberry juice on fatigue-related indices. A, the fatigue group exhibited a significantly shorter exhaust time than the control group. Treatment with Red Bull, caffeine, and low-and high-concentration blueberry juice significantly enhanced the exhaust time. B, the fatigue group exhibited significantly higher blood lactic acid (BLA) levels than the control group. Treatment with Red Bull, caffeine, and low-and high-concentration blueberry juice significantly reduced BLA levels. C, the fatigue group exhibited significantly higher blood urea nitrogen (BUN) levels than the control group. Treatment with Red Bull, caffeine, and low-and high-concentration blueberry juice significantly reduced BUN levels. D, the fatigue group exhibited significantly lower muscle glycogen (MG) levels than the control group. Treatment with Red Bull, caffeine, and low-and high-concentration blueberry juice significantly enhanced MG levels. E, the fatigue group exhibited significantly lower liver glycogen (LG) levels than the control group. Treatment with Red Bull, caffeine, and low-and high-concentration blueberry juice significantly enhanced LG levels. *P < 0.05; **P < 0.01. Cont = intact control group; F = fatigue group; RB = Red Bull group; C = caffeine group; BL = low-concentration blueberry juice group; BH = high-concentration blueberry juice group.
Figure 3 shows the effect of blueberry juice on TH+ and DAT+ cells. A significant reduction in the numbers of striatal TH+ and DAT+ cells confirmed that the changes in TH and DAT expression were caused by fatigue. However, TH+ neurons counts were only significantly increased in the Red Bull group (Fig. 3A), and whereas DAT+ cell counts were increased only in the Red Bull and caffeine groups (Fig. 3B). In addition, TH+ and DAT+ cell counts tended to increase in the blueberry juice groups; however, the results were not statistically significant.
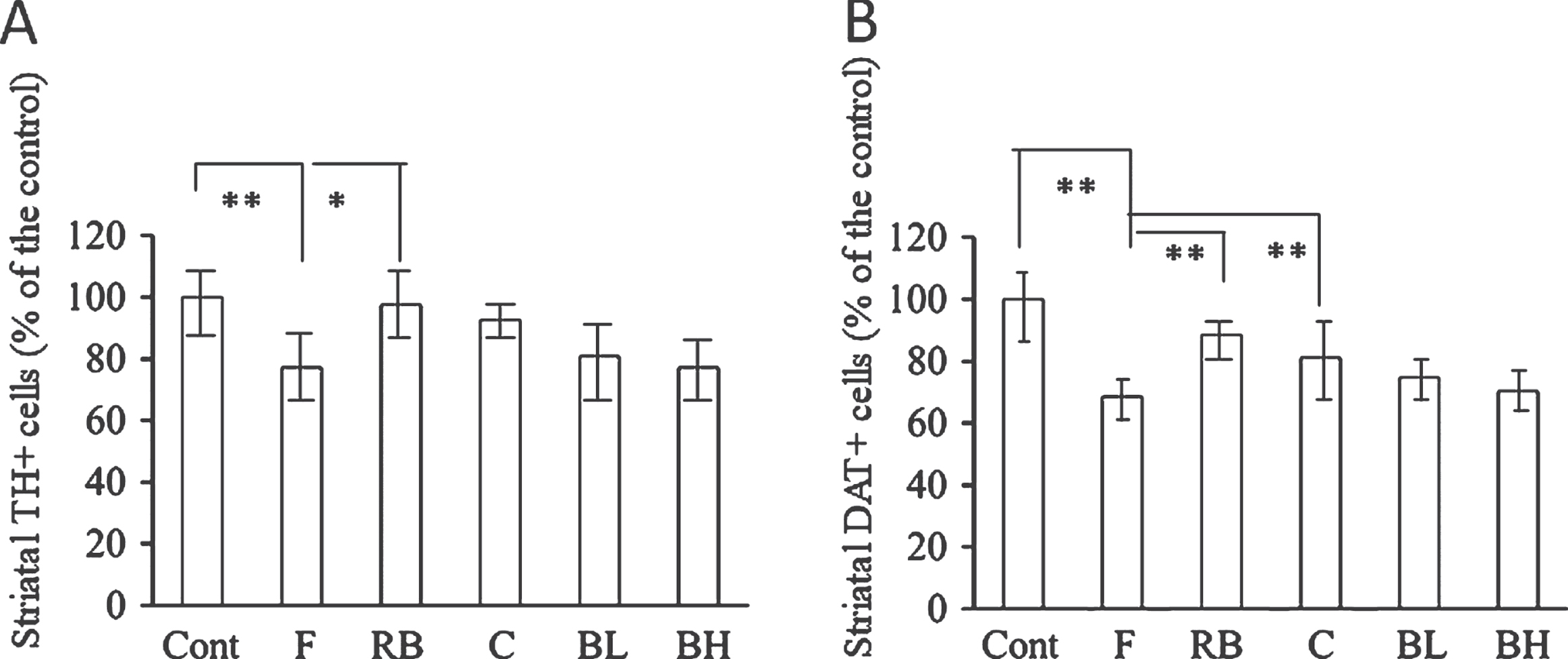
Quantitative analyses of immunohistochemical staining of tyrosine hydroxylase (TH) and dopamine transporter (DAT) in the striatum. A, fatigue processes significantly decreased TH+ cell counts in the striatum. Although all interventions tended to increase the number of TH+ cells, only the Red Bull group exhibited a significant increase. B, fatigue processes significantly decreased DAT+ cell count in the striatum. Although all interventions tended to increase the number of DAT+ cells, only the Red Bull and caffeine groups exhibited significant enhancements. *P < 0.05; **P < 0.01. Cont = intact control group; F = fatigue group; RB = Red Bull group; C = caffeine group; BL = low-concentration blueberry juice group; BH = high-concentration blueberry juice group.
Figure 4 shows that blueberry juice can reverse the downregulation of TH and DAT induced by fatigue. The results for protein (Fig. 4A-C) and mRNA expression (Fig. 4D-E) were analogous. Whereas fatigue significantly downregulated striatal TH and DAT expression, Red Bull, caffeine, and blueberry juice significantly upregulated their expression. We observed no significant difference in effect between the low and high concentrations of blueberry juice.
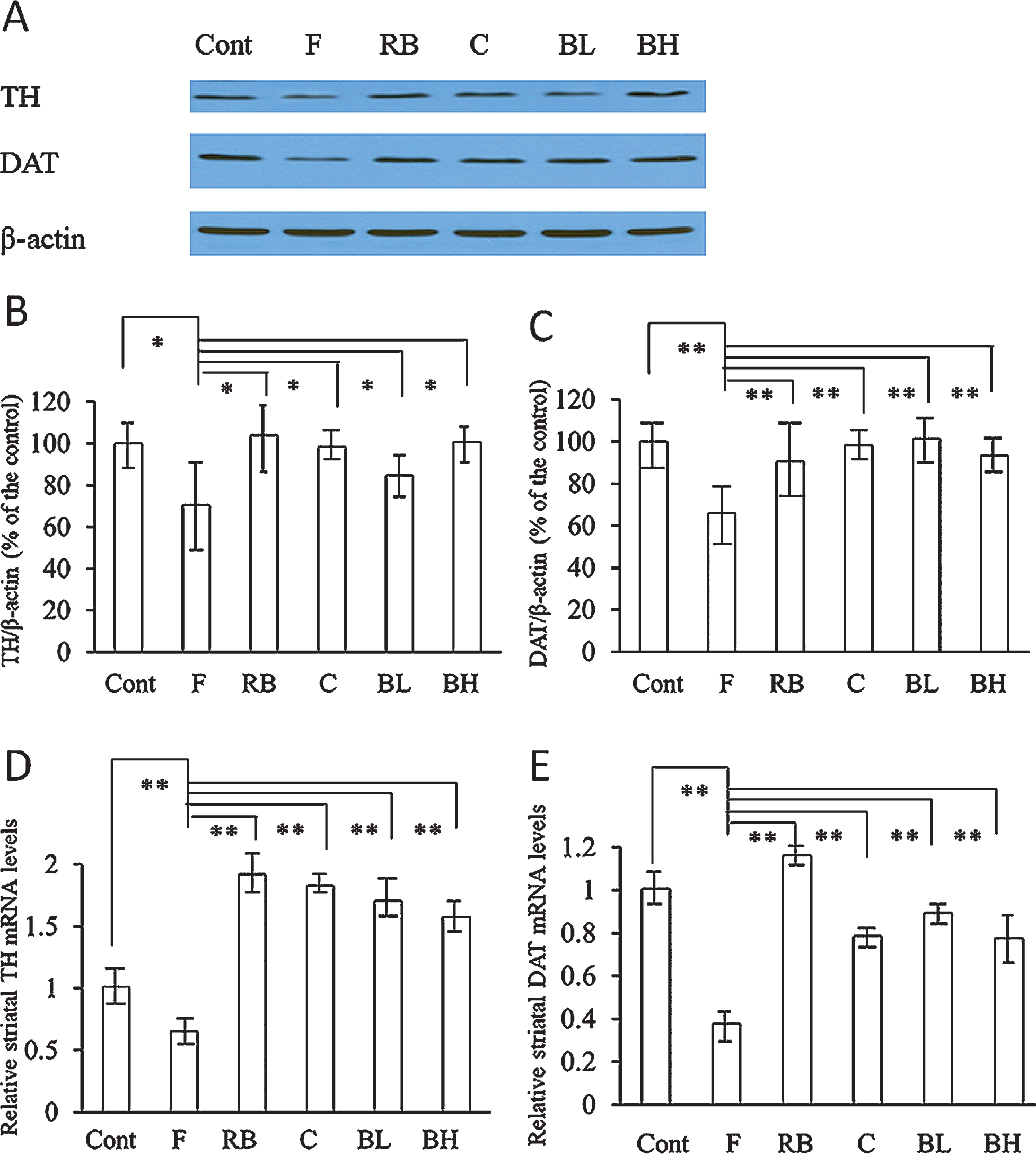
Tyrosine hydroxylase (TH) and dopamine transporter (DAT) expression in the striatum. A, Western blot analysis of TH and DAT expression in the striatum. Fatigue processes downregulated TH and DAT expression, whereas Red Bull, caffeine, and blueberry juice reversed these changes. B, quantitative data of TH protein expression detected via Western blotting. Fatigue processes significantly downregulated TH protein expression, and all interventions significantly upregulated TH protein expression. C, quantitative data of DAT protein expression detected via Western blotting. Fatigue processes significantly downregulated DAT protein expression, and all interventions significantly upregulated DAT protein expression. D, quantitative data of TH mRNA expression measured via RT-PCR. Fatigue processes significantly downregulated TH mRNA expression, and all interventions significantly upregulated TH mRNA expression. E, quantitative data of DAT mRNA expression measured via RT-PCR. Fatigue processes significantly downregulated DAT mRNA expression, and all interventions significantly upregulated DAT mRNA expression. *P < 0.05; **P < 0.01. Cont = intact control group; F = fatigue group; RB = Red Bull group; C = caffeine group; BL = low-concentration blueberry juice group; BH = high-concentration blueberry juice group.
Figure 5 shows the correlations of the exhaust time for swimming with TH and DAT expression. We determined that TH+ cell counts (Fig. 5A; r = 0.63, P < 0.01), TH protein expression (Fig. 5B; r = 0.79, P < 0.01), DAT+ cell counts (Fig. 5C; r = 0.64, P < 0.01), and DAT protein expression (Fig. 5D; r = 0.77, P < 0.01) were positively correlated with motor performance. In addition, a more extended correlated with more TH+ and DAT+ cells and stronger TH and DAT expression, whereas a shorter exhaust-time correlated with fewer TH+ and DAT+ cells and weaker TH and DAT expression.
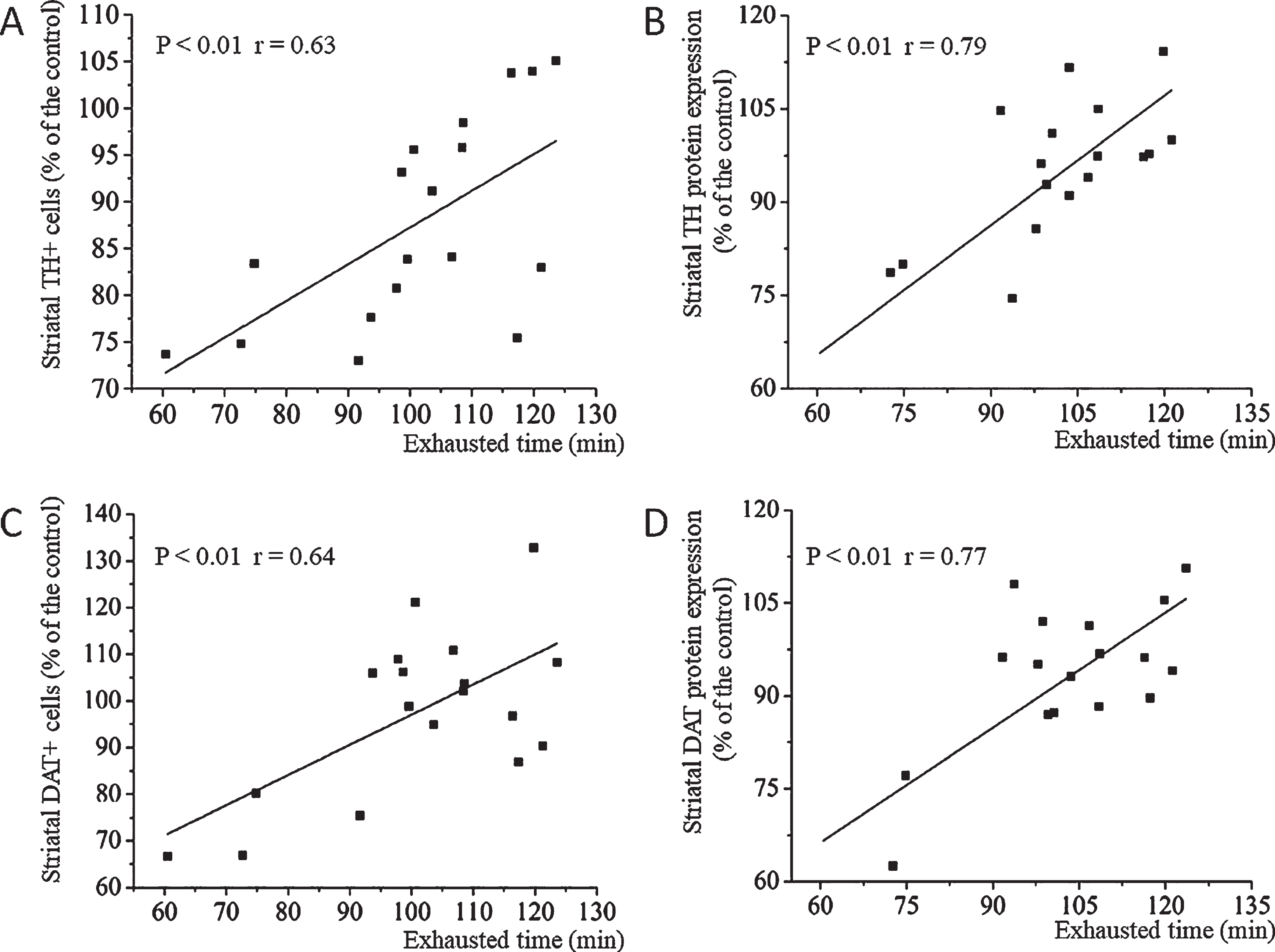
Correlations of the exhaust time for swimming with tyrosine hydroxylase (TH) and dopamine transporter (DAT) expression. A, correlations between striatal TH+ cell counts and the exhaust time for swimming. B, correlations between TH protein expression and the exhaust time for swimming. C, correlations between striatal DAT+ cell counts and the exhaust time for swimming. D, correlations between DAT protein expression and the exhaust time for swimming.
This study investigated changes in the dopaminergic system during striatum-related fatigue. The results confirmed the anti-fatigue effects of blueberry juice and demonstrated that the behavioral performance of mice was reduced in conjunction with TH and DAT expression as well as by forced swimming; however, these changes can be ameliorated using blueberry juice, Red Bull, and caffeine. To the best of our knowledge, this is the first study to provide direct evidence of the effects of changes of DAT expression on fatigue, which need further verification. In addition, this study established that blueberry juice has anti-fatigue effects independent of caffeine and taurine because the ingredient analysis in preliminary experiments of this study and previous studies [23–25] did not detect any caffeine and taurine in blueberry juice.
First, in this study, we verified the reliability of the experimental system. Our verification comprised motor performance and BLA, BUN, MG, and LG levels. We found that the motor performance was significantly reduced by fatigue but was recovered by Red Bull, caffeine, and blueberry juice consumption. In addition, BLA, BUN, MG, and LG levels were altered by fatigue. These data validated the successful establishment of our fatigue mouse model, as fatigued animals exhibited a poorer exhaust time, higher BLA and BUN levels, and lower MG and LG levels. We also confirmed the ameliorating effects of blueberry juice (anthocyanin), caffeine, and Red Bull (caffeine + taurine), confirming that this experimental system was reliable and appropriate for experimentation.
Regarding changes in the dopaminergic system, we assessed TH+ and DAT+ cell counts in the striatum and determined that fatigue processes significantly reduced their numbers. However, blueberry juice did not significantly enhance their numbers. Of note, TH+ cell counts were significantly increased only in the RB group, and DAT+ cell counts were ameliorated only in the RB and caffeine groups. By contrast, the protein and mRNA expression of both TH and DAT protein was significantly downregulated by fatigue processes and recovered by Red Bull, caffeine, and blueberry juice administration. These changes in TH+ cell counts accorded with previous findings [5]. This study revealed changes in DAT+ neuron counts and DAT mRNA and protein expression attributable to fatigue. However, this downregulation was ameliorated by caffeine, taurine, and anthocyanin. The positive correlation between behavioral performance and DAT expression strengthened the evidence that striatal DAT expression is downregulated in the fatigued state. However, the underlying mechanisms remain unknown. A previous study suggested that physical training might delay fatigue by downregulating DAT expression [7]; however, it did not provide direct evidence of changes in DAT content. We hypothesized that changes in DAT expression represent a compensatory response to fatigue. In this study, once fatigue processes were triggered, striatal TH expression was reduced. Consequently, DAT expression was compensatorily downregulated, resulting in reduced reuptake of DA to the presynapse and DA accumulation in the nigrostriatal pathway. Nevertheless, we did not measure DA or its metabolites or perform pharmacological analyses of DAT function in the present study, which are required to assess whether the changes in DAT and TH protein levels are biologically relevant. DA levels are more reliable for investigating fatigue. However, directly measuring striatal DA levels in vivo is technically difficult. We hence selected TH+ cells in the present study. Another important issue is the role of DA receptors in fatigue. Although many studies investigated the roles of DA receptors in mental fatigue, no study examined the relationship between physical fatigue and DA receptors. We will perform these experiments in our future investigation.
Furthermore, this study proved that blueberry juice is an excellent modality for reducing fatigue. We did not observe any difference between the low and high concentrations (5 and 20 mL/kg, respectively) of blueberry juice, indicating that anthocyanin exerts relatively significant anti-fatigue effects. In addition, the anti-fatigue effect of the low concentration of blueberry juice was non-inferior to those of caffeine and taurine. Hence, as blueberry juice is readily available, it is recommended as a supplemental drink to relieve fatigue.
One limitation of this study was that we did not count striatal DAT+ and TH+ cells, as upregulation of DAT and TH levels can result from enhancement of the number of DAT and TH+ neurons. We will directly assess changes in the numbers of DAT+ and TH+ cells in response to blueberry juice and Red Bull in our future studies.
Conclusions
Overall, this study deduced that striatal DAT and TH expression was downregulated by fatigue, and these changes were reversed by blueberry juice (anthocyanin), caffeine, and Red Bull (caffeine + taurine). In addition, this study provided direct evidence linking changes in DAT expression with fatigue. Hence, blueberry juice is an excellent supplemental drink for relieving fatigue that does not contain caffeine and taurine.
Funding
The authors report no funding.
Conflict of interest
No potential conflict of interest were disclosed in this study.
Authors’ contributions
TA and BJ designed the study; BJ, SZ, LA, JG, and TA performed the experiments; BJ, SZ and TA wrote the draft; BJ, SZ, LA, JG. and TA discussed and decided the last version of manuscript; TA and BJ supervised the study.
Footnotes
Acknowledgments
This work was supported by grants from the Japanese Society for the Promotion of Science (Grant-in-Aid for Young Scientists, Type B, No. 20791025 and Grant-in-Aid for Scientific Research C, General, No. 24592157,15k10358 and 18K08991).