
Abstract
BACKGROUND:
Melatonin (MT) is an endogenous indoleamine that regulates senescence progression and stress response in plants.
OBJECTIVE:
Here, we investigated the effect of MT on the shelf-life and postharvest quality of table grapes (Vitis labrusca L. cv. ‘Fengzao’).
METHODS:
After harvesting, ‘Fengzao’ grapes were immersed in MT solution at various concentrations (0 [as control], 0.05, 0.1, 0.5 and 1.0 mM for 2 h and stored at 24±1 for 15 days. Physiological indicators including weight loss rate, firmness, contents of total soluble solids (TSSs), ascorbic acid (AsA), malondialdehyde (MDA), hydrogen peroxide (H2O2), and activities of catalase (CAT), ascorbate peroxidase (APX), and superoxide dismutase (SOD) were investigated. Additionally, the DNA methylation rate of ‘Fengzao’ grapes were measured using the methylation-sensitive amplification polymorphism (MSAP) technique.
RESULTS:
Application of MT effectively delayed grape senescence in all treatment groups compared with the control, with the longest delay observed in the 0.5 mM treatment. Additionally, the rate of DNA methylation decreased in all the 0.5 mM MT treatment groups, indicating a potential role of MT in demethylation.
CONCLUSIONS:
Our results suggest that the exogenous application of MT can delay the senescence of grapes during postharvest.
Introduction
Grape (Vitis vinifera L.) has been cultivated for thousands of years and is one of the most important fruits all around the world. Grapes are rich in phenolic compounds. For example, flavan-3-ols (catechin and epicatechin), flavonols (quercetin and its glycoside derivative rutin), and stilbenes (trans-resveratrol) [1]. Phenolic compounds exhibit several important biological, pharmacological, and medicinal properties and a strong antioxidant activity, which can slow the progression of aging and reduce the risk of cancer and heart disease. At present, grape is cultivated on 7 Mha globally, of which China accounts for 12% of the total acreage, ranking second in grape production worldwide. Of the total land used for grape production in China, 84.1% is used for the cultivation of table grapes. However, senescence and disease have seriously restricted the market development of table grape [2], table grapes exhibit softening, rotting, and browning in a short time postharvest [3]. Approximately 20% of grape production is lost during storage and transportation, which severely restricts the development of fresh grapes.
Melatonin (MT) is a naturally occurring indoleamine present in various plant organs [4]. In addition to its role as a dark signal and plant growth-promoting regulator, MT also functions as an antioxidant and a free radical scavenger in plants to protect various plant tissues and organs, particularly reproductive tissues against internal and environmental oxidative stress [5–7]. MT directly reduces electrophilic radical species, owing to its electron-rich aromatic indole ring, and enhances cellular detoxifying enzymes, especially glutathione peroxidase, glutathione reductase, and superoxide dismutase (SOD). Moreover, MT as a signaling molecule that modulates ethylene, abscisic acid (ABA), jasmonic acid (JA), and salicylic acid (SA) pathways and is involved in stress tolerance, pathogen defense, and delaying senescence. High MT levels in seeds likely provide antioxidative protection and promote seed viability and germination capacity [8, 9]. However, little information is available on the effect of MT on the shelf-life and quality of grapes postharvest.
In recent years, studies have been conducted to extend the shelf-life of grapes. Currently, the most popular methods used to prolong the shelf-life of grapes include modified atmosphere packaging, low-temperature storage, and SO2 fumigation [10]. Additionally, spraying 1.5 mM SA on grapes before harvest can improve grape quality and prolong their life after harvest. SA prevents fruit softening by affecting the activities of major cell wall degradation enzymes such as cellulase, polygalacturonase, and xylanase [11, 12]. Postharvest treatment of polyamines can also maintain the quality and extend the shelf-life of table grapes. A 0.5 mM dose of polyamine can delay the degradation of total soluble solids (TSS), help maintain a high total phenolic content, and reduce the degradation rate [13].
Senescence is the last stage of plant development and is characterized by dramatic changes in cell metabolism and cell structure degeneration [14]. Factors such as hormone regulation, reactive oxygen species (ROS), protein degradation, and caloric restriction have a major impact on plant senescence [15]. DNA damage leads to increased somatic mutation rate, which eventually results in functional defects and enhanced senescence [16]. Changes in DNA methylation rates and patterns affect DNA repair and lead to altered mutation rates. DNA methylation is mediated by DNA methyltransferases, which transfer methyl groups to nitrogenous bases of DNA, adenine and cytosine. In animals, DNA methylation is thought to be one of the causes of senescence [17]. Plant nuclear genomes show a wider range of cytosine methylation patterns because of the presence of a wider range of methylation sites than animal genomes [18]. DNA methylation represents an important mechanism underlying the epigenetic regulation of plant senescence since major developmental transitions, plant stress responses, and leaf senescence are all under epigenetic control [19]. Therefore, the investigation of DNA methylation changes is of great significance for understanding the effect of MT on postharvest grapes.
The objective of this study was to investigate the effects of MT application on the senescence of grape berries during storage at room temperature. Grapes of Vitis labrusca L. cv. ‘Fengzao’ were treated with 0.05, 0.1, 0.5, and 1 mM MT, and physiological indices of grape ripening were measured. For the purpose of investigating the significance of DNA methylation-related changes in the senescence process of grape fruit, the DNA methylation rate of ‘Fengzao’ grape was determined by adopting the methylation-sensitive amplification polymorphism (MSAP) technique. Our results provide novel insights into the effects of MT on grape shelf-life and quality postharvest.
Materials and methods
Plant material and MT treatment
Vitis labrusca L. cv. ‘Fengzao’, an early-ripening bud mutant of ‘Kyoho’, was used in this study [20]. ‘Fengzao’ was collected from a grape plantation in Yanshi, Luoyang, China (longitude 112°26’15”–113°00’00”, latitude 34°27’30”–34°50’00”). Grape berries exhibiting uniform shape and appearance and no visible defects were selected for this study. All grape samples were harvested after ripening and analyzed at the Forestry College of Henan University of Science and Technology. The ripening berries are purplish black and good color uniformity. Fruit cluster average weight ranged from 475 to 555 g, averaged 485 g. Berry weight was near 8.5–13.0 g, and berries are elliptic to oblong in shape. It has a firm berry texture.
To conduct MT treatments, MT solutions at various concentrations (0, 0.05, 0.1, 0.5, and 1.0 mM) were prepared in distilled water. The selected grape berries were divided into five groups and immersed in five MT solutions (described above) for 2 h. Soak approximately 1000 g of grapes in a solution of 2 L. Grapes were then transferred to a plastic box, covered with a plastic wrap, and placed in a tissue culture room at 24±1 for 15 days. Samples were taken every 3 days for a total of 5 times. A portion of each sample used for the evaluation of weight loss, berry firmness, and total soluble solid (TSS) content. The remaining samples were collected and stored at –40 for subsequent analysis of ascorbic acid (AsA), malondialdehyde (MDA), and hydrogen peroxide (H2O2) contents and SOD, catalase (CAT), and ascorbate peroxidase (APX) activities. After treatment with 0.5 mM MT, samples were taken every 3 days for a total of 6 times and stored at –40 for subsequent analysis of MSAP.
Measurement of the physiological indices of berries
The weight of ‘Fengzao’ berries was measured before and after the 15-day MT treatment using an analytical balance to determine the weight loss rate. Weight loss[%] = (Initial weight-Sample weight/Initial weight)×100; this experiment was performed in triplicate. The firmness and TSS content of berries treated with MT were measured using the durometer (FT-327 Wuxi Yuheng Mea-suring Instrument, Wuxi, China) and handheld refractometer (WYT-4 Tianlei, Shanghai, China), respectively. Activities of SOD and CAT were measured as described previously [21]. MDA content was determined as described previously [21], with slight modifications. APX content was determined using the method of Sun B [22].
MSAP molecular marker
Genomic DNA of six grapes at different development stages in the control and 0.5 mM MT treatments was extracted using the modified cetyl trimethylammonium bromide (CTAB) method [23]. The isolated DNA was examined by 1% agarose gel electrophoresis, diluted to the required concentration, and stored at –20 for subsequent analysis. The methylation level of the isolated DNA was detected using the MSAP molecular marker [24, 25]. Briefly, genomic DNA isolated from 12 ‘Fengzao’ grape berries in the control group and treatment group were digested with EcoRI and Hpa/MspI restriction enzymes. Then the adapters of Eco-A1/ Eco-A2 and HM-A1/ HM-A2 were connected. Pre-amplification was performed as described previously to Portis E [25]. Selective amplification was performed in a 20μl volume, containing 5U Taq (TaKaRa, Dalian, China), 2.5 mM dNTPs, 10×buffer (Mg2 +), and 10μl selective amplification primer, using the following conditions: initial denaturation at 94°C for 3 min, followed by 36 cycles of denaturation at 94°C for 30 s, annealing at 56°C for 30 s, and extension at 72°C for 1 min. The PCR products were resolved on 30% denatured polyacrylamide gel and detected by silver staining.
Statistical analysis
Data were presented as mean±standard deviation (SD). Statistical analysis of data was performed using the SPSS 21.0 software. Differences among treatments were analyzed by independent sample t-test. A P value < 0.05 was considered significant. Figures were produced using GraphPad Prism 5.0.
Result
Phenotypic changes in ‘Fengzao’ berries induced by MT treatment
Different MT treatment groups showed varying degrees of delay in grape senescence (Fig. 1). The 0.1, 0.05, and 1.0 mM MT treatment groups and the control group showed browning and mildew, with the highest severity in the control group, whereas the 0.5 mM MT treatment group showed no browning or mildew (Fig. 1). Thus, the effect of MT on the freshness of grape berries was significantly higher than the control, and grape berries treated with 0.5 mM MT showed the best preservation (Fig. 1).

Effect of melatonin (MT) application on the senescence of ‘Fengzao’ grapes postharvest. MT was applied at concentrations of 0 (control), 0.05, 0.1, 0.5, and 1.0 mM after harvest.
With the increase in storage time, the water content and weight of grape berries decreased, although significant differences were observed between the MT treatment groups and control group (Fig. 2). The weight loss rates of berries in the 1, 0.1, and 0.05 mM MT treatment groups were 15.1%, 21.6%, and 11.9%, respectively lower than that of the control group (Fig. 2). The weight loss rate of grape berries was the lowest in the 0.5 mM MT treatment group (Fig. 2). The firmness of berries in the 0.5 mM MT treatment group was significantly higher than that of berries in the other MT treatment and the control groups (Fig. 3). This suggests that 0.5 mM MT treatment could effectively prevent the reduction in grape quality and firmness during storage (Fig. 3).
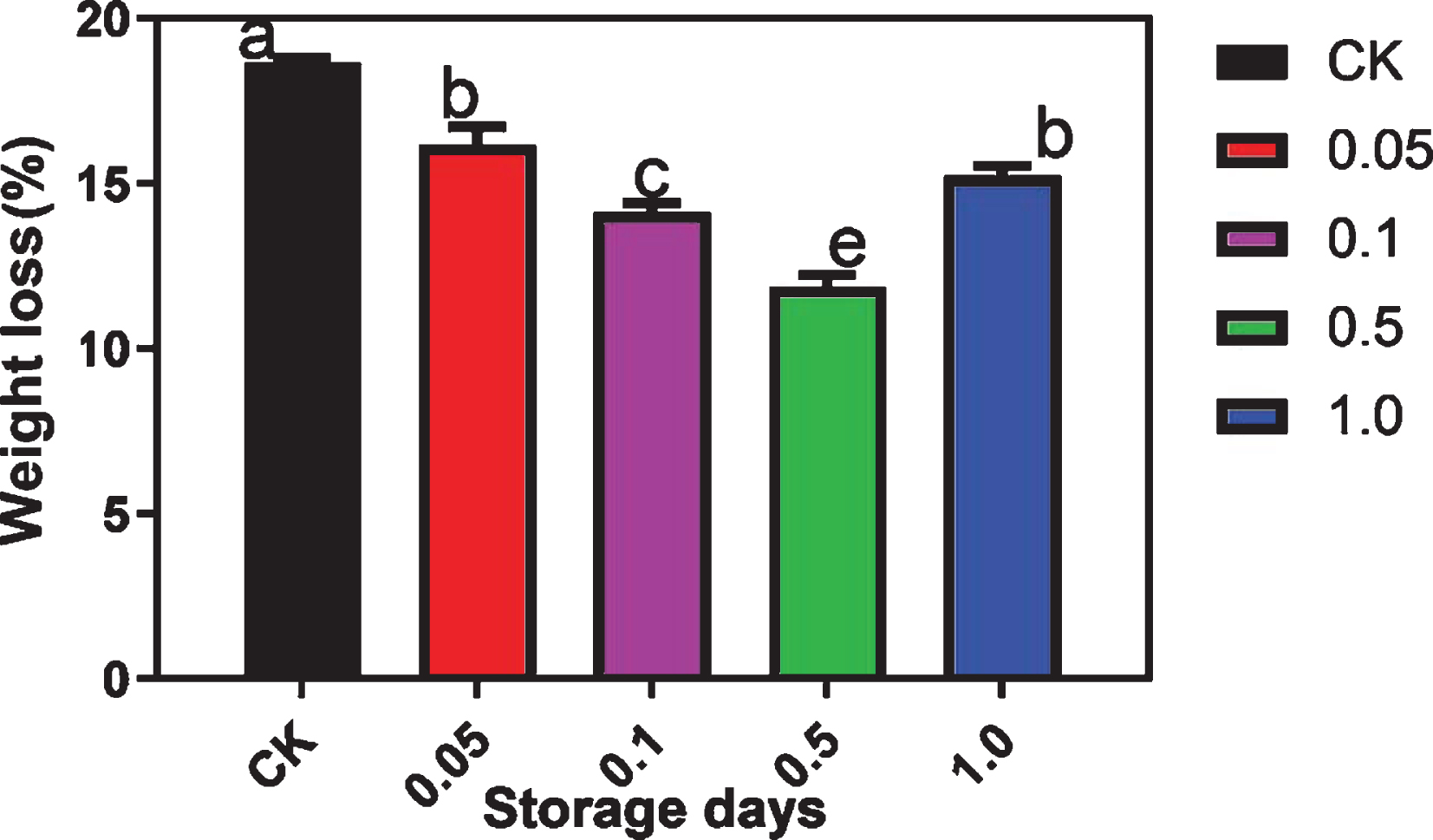
Effect of MT application on the weight loss rate of ‘Fengzao’ grapes during storage for 15 days.

Effect of MT application on the firmness ‘Fengzao’ grapes postharvest. MT was applied
at concentrations of 0 (), 0.05 ( ),
0.1 (
),
0.1 ( ),0.5 (
),0.5 ( ), and 1.0 mM (
), and 1.0 mM ( ).
Data represent mean±standard error (SE; n = 3). Significant
differences among means were determined using the least significant difference (LSD)
test (P < 0.05).
).
Data represent mean±standard error (SE; n = 3). Significant
differences among means were determined using the least significant difference (LSD)
test (P < 0.05).
The TSS content of grape berries gradually increased during storage (Fig. 4a), although this increase was significantly higher in MT treatment groups than in the control group (Fig. 4a). Among the MT treatment groups, the 0.5 and 0.05 mM MT treatment group showed the highest TSS contents (Fig. 4a). Additionally, the AsA content of grape berries increased, then decreased, and again increased upon MT treatment, and significant differences in the AsA content were observed among various MT treatment groups (Fig. 4b). The control treatment also showed a similar trend (Fig. 4b).
Effect of MT application on the level of oxidative stress markers in ‘Fengzao’ grapes
postharvest. (a, b) Concentrations of malondialdehyde (MDA) (a) and hydrogen peroxide
(H2O2) (b) in grape berries treated with 0 (control;
), 0.05 (), 0.1
(),0.5,(), and 1.0 mM
() MT. Data represent mean±SE
(n = 3). Significant differences among means were determined using
the LSD test (P < 0.05).
Changes in the MDA content of grapes were similar among all MT treatment groups and the control group (Fig. 5a). The MDA content of berries increased during the first 6 days of storage, reaching peak on day 6, and then declined from day 6 to day 15. At peak levels, the MDA content was the highest in the control group and lowest in the 0.5 mM MT treatment group (Fig. 5a). On day 15, the MDA content was the lowest in the 0.5 mM MT treatment group (Fig. 5a). These data showed that MT inhibited the production of MDA at a concentration of 0.5 mM, which slowed the senescence rate of grape berries (Fig. 5a).
Effect of MT application on the total soluble solid (TSS) and ascorbic acid (AsA)
contents of ‘Fengzao’ berries during storage. (a) TSS content; (b) ASS content.
Berries were treated with 0 (control; ), 0.05
(), 0.1 (), 0.5
(), and 1.0 mM () MT
postharvest. Data represent mean±SE (n = 3). Significant differences
among means were determined using the LSD test (P < 0.05).
The change trend of H2O2 content in the control group was similar to that in the 0.05 mM MT treatment group. The H2O2 content of the grape showed a decreasing trend in the first 6 days before storage, peaked on the day 6, and increased from the day 6 to the day 15 (Fig. 5b). 0.5 mM treatment group and 0.1 mM treatment group were similar to each other. The H2O2 content of the grape showed a decreasing trend in the first 3 days before storage, peaked on the day 3, and increased from the day 3 to the day 15 (Fig. 5b). The H2O2 content of the four treatment groups was significantly different from that of the control group on the day 6 (Fig. 5b). The H2O2 content of grape berries in MT treatment groups was remarkably lower than that in the control group during the most of storage period (Fig. 5b). The content of H2O2 was the lowest in the 0.5 mM MT treatment group and highest in the control group.
The CAT activity profile was similar among the different MT treatments and the control, with a gradual increase from day 0 to day 3, followed by a rapid increase from day 3 to day 15. The activity of CAT was significantly higher in the 0.5 mM MT treatment group than in the other MT treatments (Fig. 6a). During storage, the highest CAT activity was detected on day 3 in MT treated and control berries (Fig. 6a). We also examined the activity of APX, which was higher in all MT treatment groups than in the control group (Fig. 6a). The highest activity of APX was detected in the 0.5 mM MT treatment group (Fig. 6a). Furthermore, SOD activity showed similar profiles in the control and MT treatment groups (Fig. 6b). The activity of SOD first increased gradually and then decreased gradually during storage (Fig. 6c). The SOD activity of 0.5 mM MT group was always higher than that of the control group (Fig. 6c).
Effect of MT application on enzyme activities in ‘Fengzao’ grapes during storage.
(a–c) Activities of catalase (CAT) (a), ascorbate peroxidase (APX) (b), and superoxide
dismutase (SOD) (c) in berries treated with 0 (control;
), 0.05 (), 0.1
(),0.5 (), and 1.0 mM
() MT postharvest. Data represent mean±SE
(n = 3). Significant differences among means were determined using
the LSD test (P < 0.05).
Because treatment of grape berries with 0.5 mM was the most effective in delaying senescence, we investigated the DNA methylation level of berries in this treatment group and the control group (Supplemental Figure 1). Compared with the control, 0.5 mM MT treatment group showed a lower DNA methylation level. The methylation rate of grapes treated with 0.5 mM MT in six periods was significantly lower than that of the corresponding control group (23.3 < 28.9%, 23.2 < 26.7%, 22.8 < 25.3%, 22.7 < 25.2%, 20.9 < 24.6%, 20.0 < 24.0%) (Supplemental Figure 1) (Supplemental Figure 2). This suggests that MT treatment demethylates DNA.
Discussion
Senescence is a very complex process and marks the last stage of plant growth. In grape, senescence is generally associated with an increase in TSS, AsA, H2O2, and MDA contents and a reduction in berry firmness, weight, and APX, CAT, and SOD activities [26, 27]. In this study, spraying of 0.5 mM MT on ‘Fengzao’ grapes postharvest significantly reduced berry weight loss and helped maintain berry firmness, TSS content, and AsA content during storage at room temperature. Additionally, MT delayed senescence and helped maintain the quality characteristics of grape berries.
Membrane lipid peroxidation is a major cause of plant cell senescence [28]. Peroxidation of membrane lipids leads to the loss of membrane integrity, physical structure, and fluidity, which in turn affects protein function [28]. Loss of membrane integrity during senescence stress during the storage of ‘Fengzao’ grapes is generally linked with excessive accumulation of ROS including H2O2 and MDA. MDA is the end product of peroxides produced by intracellular free radicals as lipids, and it is considered to be the key substance to cause plant senescence because it can aggravate the damage of lipid to cell membrane [29]. The MDA content of each treatment group was lower than that of the control group, and the MDA content of 0.5 mM treatment group was the lowest. The results showed that MT treatment significantly inhibited the accumulation of MDA in the late storage period and played a role in reducing lipid peroxidation and delaying the senescence process of grapes.
Plant senescence is generally associated with a change in cell membrane integrity [30]. Altered composition and functionality of mitochondria and chloroplasts, and changes in a protein in organellar membranes [30]. These factors can negatively affect the function of protein complexes, causing oxidative stress. To cope with oxidative stress, plants employ different enzymatic antioxidant systems [31]. In the current study, among the enzymatic systems that remove ROS, the activities of CAT, APX, and SOD stand out. SOD protects cells from oxidative stress, while CAT and APX control the decomposition of H2O2. Previous studies suggest that high activity of these antioxidant enzymes and their synergistic effects are part of a mechanism that reduces lipid peroxidation and delays fruit senescence [32]. For example, MT-induced delay of peach fruit senescence is considered to be the result of simultaneous enhancement of SOD, CAT, and APX activities [33]. MT helps to remove ROS, inhibit lipid peroxidation, and delay peach fruit senescence.
Our results showed that MT treatment delayed the senescence of grapes after harvest. In a previous study, treatment of ‘Kyoho’ berries with 100μm azacytidine (5-azac), a methyltransferase inhibitor, promoted early ripening by 20 days compared with the control, and increased the DNA methylation rate at all stages of berry development [34]. By contrast, our results showed that treatment of grape berries with 0.5 mM MT can delay senescence. Our results are agreement with a previous study [35], which showed that the exogenous application of MT promotes fruit ripening and delays fruit senescence.
In higher plants, MT is an important metabolic regulator. The level of MT is high in young and reproductive tissues and declines with age to significantly level in postharvest fruits [36]. MT acts as an antioxidant during fruit and seed development to increase the survival and perpetuation of the species [37]. Additionally, MT plays an essential role in regulating plant growth and improving fruit quality and plant stress resistance. In tomato, exogenous MT activates the CBF1 signaling pathway by upregulating SLZAT2/6/12, triggering arginine pathway activity, and increasing cold resistance [38]. Our results suggest that MT protects cell structure, prevents DNA damage, and reduces the level of peroxides in plants by scavenging free radicals and inhibiting lipid peroxidation.
DNA methylation is widespread in plant cells and is involved in regulating plant gene expression and plays a very important role in plant growth and development. DNA methylation changes with plant development. Previous studies suggest that genomic DNA undergoes higher methylation during plant senescence than during other growth stages, leading to changes in gene transcription during plant senescence [39, 40]. Changes in expression or activity of DNA methyltransferases and/or DNA demethylases during plant senescence may be related to changes in DNA methylation levels [41]. Methylation of genes inhibits gene expression. When the gene is expressed, methylation levels tend to be low, and it needs to close the gene as it grows and develops. Repethylation occurs in the promoter or coding region of the gene, which inhibits gene transcription and deactivates the gene, thus terminating its expression. And some genes in the closed state need to be activated, so as to open the expression. Methylation levels of the same species also vary greatly in different stages and tissues. We used MSAP techniques to investigate differences in DNA methylation levels in melatonin-treated grape fruit, and further validate the correlation between aging and DNA methylation, thus aiming at explaining the aging mechanism in the field of epigenetics. In this study, the incidence of DNA methylation in fruit of grapes treated with melatonin decreased. These results indicate that DNA methyltransferase gene and deoxyribonucleic acid demethylase gene expression decreased the DNA methylation rate of grape during post-maturing.
In previous studies, it could significantly prolong the storage period of grapes through fumigating the grapes with 500μL/L SO2 for 1 hour. However, it caused bleaching damage and showed that residual SO2 was harmful to human health. When compared with the control, 0.5 mmol/L MT treatment increased the quality of ‘Fengzao’ grape fruit by 15 days. Different from SO2 treatment, MT treatment has no bleaching damage to fruit, and MT is a widely used health product to enhance human immunity and anti-aging. Therefore, it is safe to use melatonin to treat grape fruit.
Conclusions
In this study, clusters of ‘Fengzao’ grapes were immersed in different concentrations of MT solution for 2 h. All MT concentrations delayed the senescence of ‘Fengzao’ grapes stored at room temperature. however, the 0.5 mM MT treatment was the most effective. The treatment of MT significantly affected the physiological characteristics of berries. To understand the basis of this phenomenon, we investigated the DNA methylation levels of grapes in various treatments using the MSAP molecular marker. Our results showed that DNA methylation levels in MT treatment groups were lower than that in the control group, suggesting that MT extends the shelf-life of ‘Fengzao’ grapes by lowering the degree of DNA methylation. Thus, our results provide a safe, efficient, and cheap method to preserve grapes postharvest.
Conflict of interest
The authors declare no conflict of interest to this study.
Authors’ contributions
Yi-He Yu and Da-Long Guo designed the study. Yi-He Yu and Ya-Dan Sun wrote the majority of the original draft of the paper. He-Cheng Zhang, Lei-Lei Wang, Min-Li participated in generating the data for the study. Yi-He Yu, Da-Long Guo, Sheng-Di Yang and Ya-Dan Sun participated in the analysis of the data. Yi-He Yu and Ya-Dan Sun proofread the paper. All authors are in agreement with the manuscript.
Footnotes
Acknowledgments
This work was financially supported by the Henan province key technology R&D and extension program (202102110043), National Natural Science Foundation of China (31701893), National Key Research and Development Program of China (2018YFD1000105), the Training Project for Young Backbone Teachers of Colleges and Universities of Henan Province (No. 81), New Agricultural Research and Reform Practice Project in Henan Province (2020JGLX121), and Education and Teaching Reform Project of Henan University of Science and Technology (HKDJG2014076).