
Abstract
Metabolomics, one of the most recent additions to the systems biology toolkit, has been used to study the metabolism and physiology of living organisms. In the field of food research, metabolomics has lately gained popularity as a technique for determining the quality, processing, and safety of raw materials and final food products, as well as for identifying biomarkers linked to food intake and disease. This review aimed to provide an overview of the current understanding of the analytical techniques used for metabolomics and their application to berries with a special emphasis on the in vivo studies. Specifically, identification of biomarkers of biological activity and in vivo metabolites of berries using metabolomics approach is covered.
Introduction
Metabolomics allows for the simultaneous identification of a large number of compounds while also correlating their bioactivity [1]. The rapidly expanding field of metabolomics has enabled the high-resolution characterization of hundreds or thousands of metabolites from complicated samples in a single measurement. Metabolomics has been used to discover biomarkers of certain illnesses [2], investigate unknown metabolic pathways, stress tolerance mechanisms [3], and improve and create microbial strains [4] in a variety of biological systems, including microbial, mammalian, and plant systems. Food metabolomics is the application of metabolomics to food systems. This can be implemented in all aspects of the food system, from farm to consumers, including crops quality monitoring, industrial food processing, and determination of the relationship between human and food metabolome (e.g., for berries see Fig. 1). Given that food’s rheological and sensory characteristics (appearance, odor, texture, flavor, taste) as well as specific attributes (nutritional value and health benefit) are largely determined by its chemical content, metabolomics provides tremendous opportunities to obtain valuable detailed information that can be directly correlated to food safety, quality, and biological characteristics, etc. [5].
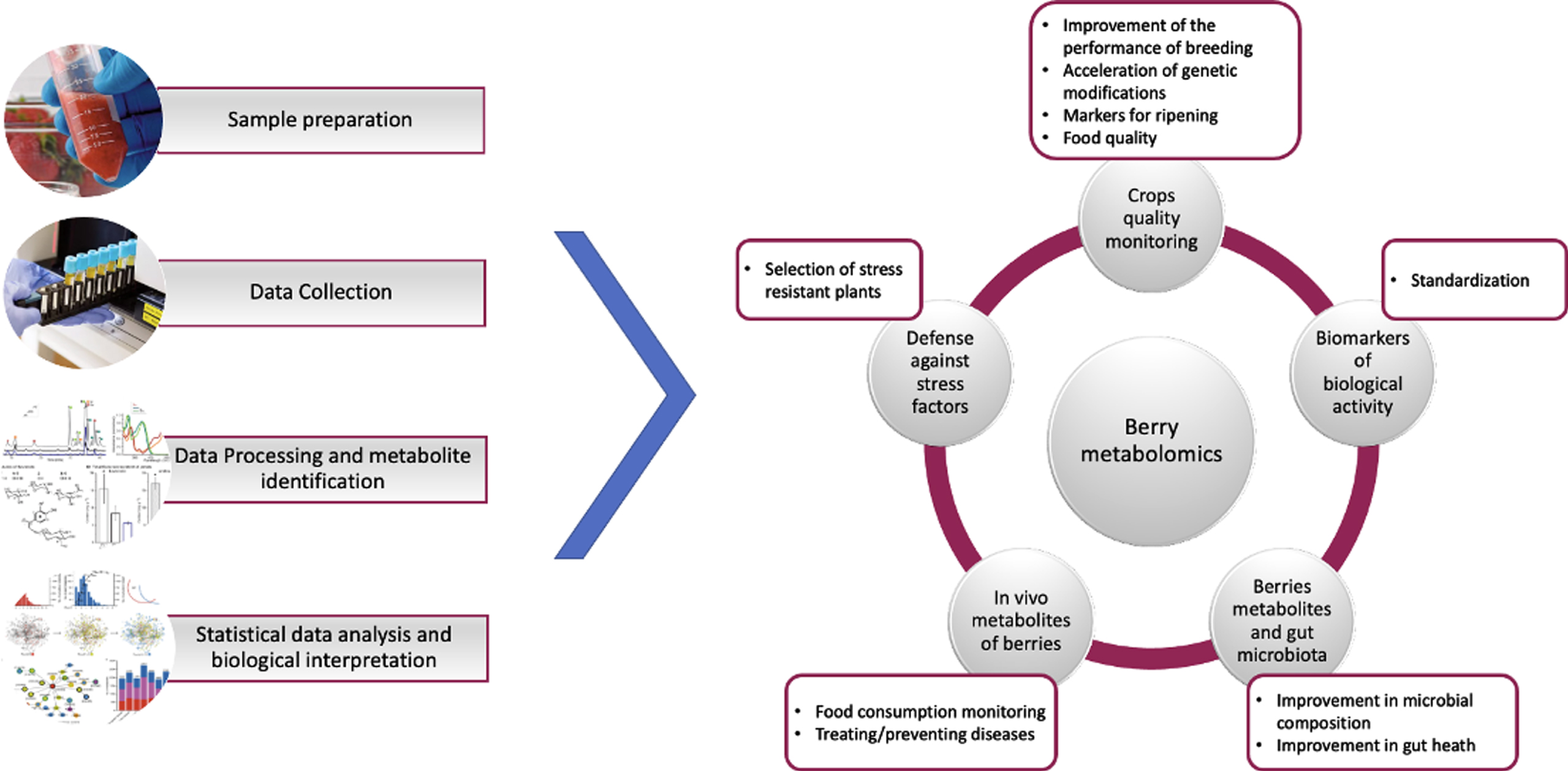
General steps of a metabolomic analysis and application areas of metabolomics analyses for berry fruits.
Metabolic profiling can only become practicable as a result of recent technical advancements in small molecule isolation and detection. These include separation techniques such as gas chromatography (GC), high performance (HPLC) or ultra-performance (UPLC) liquid chromatography, capillary electrophoresis (CE), as well as the coupling of these instruments to precise detection techniques such as nuclear magnetic resonance (NMR), mass spectrometry (MS), and near-infrared spectrometry (NIR) [6].
Within the plant kingdom, berries are one of the richest sources of numerous polyphenols. Regarding the polyphenolic composition, anthocyanins constitute the major phenolic compounds identified in berries. Apart from anthocyanins, they are also a rich source of proanthocyanidins, tannins (e.g., ellagitannins), flavonols (e.g., kaempferol), phenolic acids (e.g., ferulic acid), and flavanols (e.g., epicatechin) [7–9]. These compounds have been proven to have a wide range of biological activities such as anti-inflammatory [10, 11], anti-cancer [12–14], anti-diabetic [15, 16], etc.
Identifying the possible availability of polyphenols after digestion is essential since past research have shown that if a polyphenol’s bioavailability is low, it will have a limited influence on health because polyphenol bioactivity is dependent on bioavailability [17]. Most phenolic compounds exist in the food matrix in their native forms, such as polymers, esters or glycosides. However, once they are ingested, they may not be the most bioactive molecules because of their high catabolism and weak absorption rates. Therefore, they must be hydrolyzed to become absorbable in the gastrointestinal tract. At this point, both gut microbiota and intestinal enzymes have a significant role in this hydrolyzation process in which the size of the parent molecule is reduced, and its physiological functions are altered. Accordingly, instead of the main polyphenol, its metabolites might be able to reach the systemic circulation and target organs and exhibit their biological activities [18–20]. Correspondingly, the consumption of polyphenols in the diet directly affects the human health and disease due to the interaction between food metabolome and endogenous host metabolome. Therefore, it is important to track the relationship of berry intake in humans with diseases through the disclosure of dietary biomarkers. On the other hand, advances in the analytical instrumentation provide complete and simultaneous identification of metabolites in human biological samples. In this review, identification of biomarkers of biological activity and in vivo metabolites of berries using metabolomics approach is also evaluated. Besides, determination of the effect of berry metabolites on gut microbiota is discussed. In short, recent applications of metabolomics to berries covering from fields to the consumers are addressed.
One of the most important aims and challenges in the productions of berry fruits is sustainability. It includes constant monitoring and management of the crop during cultivation, harvest, packaging and transport. High-quality crops/products generate higher profits and result in less waste, which in general is high in the case of soft fruits. Metabolomics, defined as the targeted and untargeted small-molecule analysis supported by bioinformatics, provides information on the relationship between genetic conditions or physiological state of the plant/crop and the chemical characteristics. Metabolome-based breeding enables the selection of plants with the desired level of nutrients, yield or quality [21, 22]. This knowledge contributes to the improvement of the final product by introducing the procedures/steps related to pre-harvest and post-harvest treatments.
Improvement of the performance of breeding
The quick progress in plant metabolomics stimulates the development of more rational designs of cultivation associated with yield or quality [23]. As a consequence, the correlation of pre-harvest factors with the chemical composition of berries leads to the improvement of the performance of breeding. Therefore, a number of recent metabolomic studies focused on the relationship between berries development and quality in order to understand their biology [24-26]. When combined with transcriptomics or genomics, this information can further deepen the knowledge about phenotypic response to the genetic and environmental conditions such as: origin of the species/variety [27]; climatic conditions; production system; irrigation systems; fertilization, or phytohormones.
Origin of the species/variety
Metabolomics showed that fruit origin is a determining factor for the gain and loss of strawberry flavor. It was observed that their aroma is changed in the process of development and domestication. Wild species were abundant in olefinic monoterpenes and myrtenyl acetate, while cropped strawberries were rich in the monoterpene linalool and the sesquiterpene nerolidol [28], which obviously affects their flavor and taste. Similarly, blueberries differed in firmness, flavor and harvesting time depending on the variety studied. The application of metabolomic, ionomic and proteomic approaches revealed enrichment of proteins associated with the tricarboxylic acid cycle, nitrogen, small molecules and cell wall in muro recycling processes in firm fruits. In contrast, mannitol and fatty acid metabolism were enhanced in the soft variety [29]. Varieties of wild strawberries differing in color also displayed distinct metabolomic profiles. Red fruits had higher quality since they were rich in flavonoids, free amino acids, and vitamins in comparison to white fruits, which were poor, particularly in pelargonidin O-hexosyl-O-hexoside, pelargonidin O-hexoside and petunidin O-rutinoside, which might contribute to color development [30]. Such observations, made on the basis of metabolomic analysis may be helpful for the guided development of new varieties based on their biochemical quality.
Climatic conditions
Climatic conditions, including the location of the plantation, sunlight, rainfall or temperature fluctuations, are reflected by the quality of the crops and their biochemical state. Metabolomics can be applied to select the best natural cultivation or harvesting sites of berries [31, 32]. It was shown that the content of primary and secondary metabolites contributing to the nutritional value and taste of Vaccinium spp. berries are changing with sunlight and rainfall, temperatures and increasing altitudes [33]. Bilberries from different natural habitats differed significantly in the quantity of sugars, organic acids and phenolic compounds. Fruits from sites with high photosynthetic active radiation contained higher levels of total sugars, anthocyanins, flavonols and hydroxycinnamic acids and lower levels of organic acids compared to bilberry fruit from low-light locations [34]. Blueberries required higher solar exposure for normal ripening and anthocyanin accumulation [35]. Also, in cranberries, exposure to light had a positive effect on the accumulation of anthocyanins [36]. Similar observations were made for grapes, which were enriched in sugar, amino acids, Na, and Ca depending on the geographic conditions, and depleted in organic acids [37]. In addition to exposure to sunlight and altitude, temperature also affected anthocyanins biosynthesis in grapes. High temperature resulted in difficulties in pigments accumulation, while the low temperature at night promoted anthocyanin biosynthesis [38]. A more detailed study revealed that high temperature probably induces peroxidases and oxidoreductases responsible for anthocyanin degradation [39] and lowers the expression levels of critical enzymes of the anthocyanin biosynthesis [40]. By collecting information correlating variable climatic conditions and the chemical quality of berries, metabolomics can help the observations and indicate the best cultivation conditions. Moreover, metabolomics distinguishes crops according to the production systems (net-house or open-field) like it was done for tomatoes. Using headspace solid-phase microextraction (HS-SPME) coupled with gas chromatography to provide chemical biomarkers for tomatoes grown under fully and partially controlled conditions, Lee et al. found that the majority of volatiles released by berries was significantly increased in the net-house-grown tomatoes with geranyl acetone and D-limonene as potential biomarkers specific to these conditions [41]. Such approach might be applied for the verification of the crops production conditions.
Irrigation
Irrigation is considered a critical factor in increasing crops yield and quality however, the new type of water-saving irrigation technology, called regulated deficit irrigation (RDI), was proved to be effective in the cultivation of grape berries. Metabolomics integrated with transcriptomics revealed that plants subjected to controlled lack of water increased reducing sugar content and total anthocyanin content. In addition, the expression levels of key genes in the anthocyanin biosynthetic pathway were upregulated, resulting in higher contents of metabolites involved in the pathway (cinnamic acid, naringenin chalcone, naringenin and eriodictyol) [42]. This new approach validated by metabolomic studies might be instrumental in increasing climate warming, where drought tolerance becomes essential under natural conditions [43].
Phytohormones
Metabolomics provides an understanding of the regulation of fruit development and ripening caused by natural and synthetic growth stimulators, as it was proved for synthetic cytokinin used in fruit production. For example, the effects of the treatment of kiwi fruit with N-(2-chloro-4-pyridyl)-N’-phenylurea were explored with LC-MS and GC-MS, which revealed that during fruit development, the production of proteins involved in defense and synthesis of polar metabolites related to sugar homeostasis, cell wall modifications, tricarboxylic acid (TCA) cycle and myo-inositol pathway were increased [44]. In a similar way, the mechanism of action of other synthetic substances promoting the growth of plants may be elucidated using metabolomics.
Markers for ripening
Metabolomics provides detailed characteristics of fruit development in terms of chemical composition. This enables the determination of proper time of harvest or storage conditions, aiming to reduce or delay off-flavour formation, discolouration and nutritional losses [45]. Considering large distances which have to be covered to provide the customer with off-season fruits (import from warm countries) and the need for prolonged shelf life, the crops are often harvested before ripening. This implies post-harvest treatments, which should be tightly controlled.
Biochemical markers for ripening
Metabolomics-assisted crop improvement includes characterization of biochemical markers of ripening that better reflect fruit maturity than harvest time [46]. Monitoring of primary and secondary metabolites over time and within fruit sections was extensively done for Solanaceae fruits. Fruit development was correlated with the accumulation of coumaroyl- and caffeoyl-glycosides in the fruit peel of tomato, eggplant and pepper. Flavonol-tetra-glycosides, were elevated only in tomato fruits, while naringenin chalcone, one of the major ripening markers in tomato fruits, was also conserved among almost all pepper species [24]. Another example was monitoring biochemical markers of on-vine or off-vine ripening in tomatoes, which unlined changed metabolic state after the fruit harvest. Sucrose and glucose levels in off-vine ripened red fruit were fivefold lower than in on-vine ripened fruit. There were also lower levels of flavor-affecting malate in on-vine ripened red tomato fruit, as well as significantly lower putrescine levels, which plays an important role in suppressing fruit senescence, suggesting that ripening after harvesting might extend a fruit life [47].
Storage conditions
The metabolomics-driven analysis followed by linear modelling was applied to select the best post-harvest storage conditions for strawberry fruits, which has quite a short post-harvest time [25]. Among ambient, CO2-enriched and O3-enriched atmospheres, the ozone-enriched atmosphere appeared to be the best in counteracting post-harvest deterioration. The levels of metabolites related to fermentation (e.g. pyruvate) and produced off-aroma compounds such as ethanol, acetaldehyde and ethyl acetate were decreased, indicating better organoleptic and nutritional properties. Additionally, O3-treatment induced metabolic reconfiguration manifested by increased levels of sugars and sugar phosphates, playing a role in plant cells osmotic adjustment in response to cold and dehydration stresses [25]. The other application of metabolomics in untargeted and targeted mode was understanding the post-harvest red drupelet disorder. This physiological disorder is often found after cold storage and affects blackberries causing the post-harvest reddening of drupelets, which results in economic loss due to a reduction in marketability. The detailed monitoring of metabolite changes during color reversion revealed a decrease in anthocyanins, sugars and organic acids, lipids, and free amino acids levels after seven days of storage. However, at the harvest time, the metabolic profile of black drupelets and these turning red were already differentiated by higher levels of lysophosphatidylcholine in red drupelets, indicating that red drupelet disorder may be related to cell membrane integrity and susceptibility to mechanical damage [48]. The metabolomic analysis explained the disorder and indicated the handling factor as important during harvest and storage. Besides atmospheric conditions and handling, the use of chemicals during berries storage was also investigated. The effects of post-harvest treatment of kiwifruit with synthetic cytokinin (N-(2-chloro-4-pyridyl)-N’-phenylurea) was monitored with LC-MS and GC-MS. Metabolomic analysis showed that cytokinin suppressed ethylene production and retained central placenta softening, which results in prolonged fruit life [44]. As indicated above, metabolomics is useful for selecting the best post-harvest treatment, aiming to increase crop quality and reduce waste.
Acceleration of genetic modifications
The combination of metabolomics with transcriptomics can also provide valuable information about yield-related phenotypic traits. Determination of the key putative genes correlated with metabolites of interest gives a basis for organisms’ genetic modifications toward the improvement of nutritional qualities of berries. For this purpose, used gene-metabolite network analysis in tomato samples and assigned several candidate genes putatively involved in lipid biosynthesis or regulation. They found negative correlations between various triacylglycerols and lipid-derived volatiles, indicating indirect interdependence between lipid biosynthesis and flavour intensity [49]. In a similar manner, integrated metabolomic and transcriptomic revealed genes related to the synthesis of flavonoids, alkaloids and polyphenols in wild strawberries (Fragaria pentaphylla), providing new insights into the molecular mechanisms of the color and nutrients in strawberries [30]. These candidate genes might be subjected to biotechnological engineering for crop improvement.
On the other hand, untargeted metabolomics was suggested as a routine protocol for assessing genetically modified to supervise production of intended and unintended metabolites in transgenic fruits. Although developed genome-editing technologies introduce desirable traits more quickly and more precisely, the safety assessment process for new modified crops should also be robust, and accurate and this can be assured by metabolomics [50].
Food quality
The quality of products available on the market constantly needs supervision. For example, adulterations occur in dry berries, however more frequently in food products. The markers are needed to confirm the quality and eliminate the possibility of adulteration. Metabolomics enables fast and robust identification of biomarkers and separation of samples into groups based on their qualitative and quantitative composition.
Adulteration markers
The biomarkers discriminating goji berries: Lycium chinense and Lycium barbarum were revealed by LC-ESI-TQ-MS and GC-EI-MS. L. chinense was more abundant in chlorogenic acids, asparagine and quinic acid, whereas L. barbarum accumulated more lycibarbarphenylpropanoids A-B, coumaric acid, fructose and glucose [51]. What is more, even goji berries obtained from different geographical locations can be distinguished. An example of the product adulteration is the addition of non Zhongning goji berries (NZNG) to Zhongning Goji berries (ZNG) in marketed Daodi medicinal material (DMM) in China. To detect adulterated products, the untargeted metabolomics approach based on ultra-high performance liquid chromatography quadrupole time-of-flight mass spectrometry (UHPLC-Q-TOF-MS) was used. As a result, seven compounds were identified as biomarkers of authentic samples (quercetin, N-methylcalystegine C1, N-trans-feruloyltyramine, N-trans-feruloyloctopamine, succinic acid, gingerglycolipid B and a previously unreported glycoside of pyrrolidine alkaloid) and were statistically proved to be reliable and robust combinative biomarkers that can be used to distinguish between ZNG and NZNG [52].
Organic crops
In line with the growing awareness about the impact on human health of the chemicals used to cultivate fruits, grows the appreciation of organic systems. Metabolomics is a suitable tool to differentiate fruits from organic and conventional production to prevent food fraud in products labelled as “organic”. In targeted mode, it verifies the absence of pesticides and chemical fertilizers, whereas untargeted discriminative analysis reveals the chemical fingerprints of different agricultural management. The absence of synthetic pesticides and fertilizers in organic farming stimulates plants to produce more secondary metabolites in order to protect themselves against phytopathogens and assimilate nitrogen more effectively. Therefore, organic production is considered twice more beneficial for humans, and the determination of reliable chemical markers to discriminate between organic and conventional products is essential [23].
The comprehensive metabolite profiling by advanced technologies based on high performance liquid chromatography-high-resolution accurate mass spectrometry (HPLC-HRAMS) supported by bioinformatic tools has been used for marker search in organic and conventional tomatoes grown in a greenhouse under controlled agronomic conditions. The set of analyses revealed specific biomarkers present in organic crops: L-tyrosyl-L-isoleucyl-L-threonyl-L-threonine, trilobatin, phloridzin, tomatine, phloretin and echinenone. It also confirmed the absence of pesticides and the presence of one insecticide obtained from the fermentation of Saccharopolyspora spinosa and allowed in organic production [23]. In the other study, untargeted ultra-performance liquid chromatography with triple quadrupole mass spectrometric (UPLC-QqQ-MS/MS) metabolomics was proved to be efficient in determination of differences in chemical composition of cape gooseberry organic and conventional samples. Two withanolides (steroids built on an ergostane skeleton) and one fatty acyl glycoside were assigned as biochemical markers of organic fruits of this species [53]. Also, untargeted NMR metabolomics showed that organic samples of pomegranate had higher levels of essential amino acids, organic acids and phenolics, indicating their better nutritional quality. Interestingly, pomegranates cultivated conventionally had significantly higher concentrations of acetone, aspartic acid, and ethanol which are markers of physiological stress [54]. This confirms that the organic cultivation system is some kind of challenge for plants; therefore, they adapt by changing their metabolic pathways.
Insight into plants/fruits environmental challenges (defense against stress factors)
Recently metabolomics has been proposed as a prediction tool for plants performance under environmental stresses. Because metabolome is the final state of the cell, the product of genome but also the fingerprint of many internal and external stressors, it is considered as the best reflection of the interaction between the biological systems and their environment [43]. Interaction with the environment includes biotic and abiotic stress conditions, which can significantly affect the quality of fruits. Metabolome based breeding is a kind of strategy to select stress-resistant plants [23].
Abiotic stress
Abiotic stress is mostly related to open field conditions, where crops are exposed to several stress factors simultaneously. Plants respond to a combination of different abiotic stresses at the same time and studying each of the different factors applied independently does not fully reflect the environmental challenges of plants [43]. Nevertheless, metabolomics was successfully applied to explain metabolic reorganization in berry yielding plants under challenging conditions such as drought, extreme temperatures, salinity or light [39, 55]. Metabolomics was also helpful to explore the use of certain chemicals as elicitors increasing the stress tolerance [3, 56].
Ferrandino and Lovisolo (2014) extensively reviewed the effects of abiotic stress on grapevines, highlighting the correlation between the polyphenol accumulation in berries and different stress conditions. Metabolomics was a significant contribution in these studies, which indicated that abscisic acid is a key signaling molecule associated with abiotic stress. Its production stimulated especially by water scarcity, light, or temperature-induced the accumulation of secondary metabolites in berry pulp, seeds and skin as a defense against cell damage. Moreover, the exogenous application of abscisic acid resulted in hastened ripening and increased color of berries, especially in stress conditions [39]. The use of metabolomics and multivariate statistical analysis also revealed other signaling compounds in grapevines under water stress. Plants without irrigation produced higher amounts of 2-hexenal and 3-hexenal, which were closely related to changes in proline, hydrogen peroxide, malondialdehyde, catalase and superoxide dismutase levels, indicating that these two volatiles act as signaling molecules [55].
Through the application of metabolomics and genomics, light stress –solar UV-B irradiation was correlated with accumulation of quercetin and kaempferol in grape tissues, as well as carotenoids in leaf tissue [57]. Two different intensities of ‘field-like’ UV-B light were also examined and resulted in different metabolic profiles of grapevines. The low UV-B increased the levels of triterpenes sitosterol, stigmasterol and lupeol, in young leaves, whereas in mature leaves the accumulation of antioxidants such as diterpenes, tocopherols, phytol, E-nerolidol. High UV-B irradiation induced the production of monoterpenes such as carene, alfa-pinene and terpinolene [58]. On the contrary, UV-C treatment did not affect the composition of strawberry metabolites such as anthocyanins, glucuronides and glucosides of quercetin as well as kaempferol, pelargonidin rutinoside, catechin, and ellagic acid which were analyzed by ultra-high performance liquid chromatography high resolution mass spectrometry (UHPLC-HRMS) [59].
The role of some external molecules as plant elicitors was also investigated with metabolomics. An ultrahigh pressure liquid chromatography accurate mass quadrupole time-of-flight mass spectrometry with electrospray ionization (UHPLC-ESI/QTOF-MS) untargeted metabolomic analysis enabled to define metabolites increased under chitosan treatment and potentially involved in grapevines defense. After ten days of exposure, higher amounts of pentacyclic triterpenoids were detected, which may be important in fungal defense [56]. Also, omeprazole applied as substrate drench for tomato plants in NaCl salinity stress altered the production of metabolites profiled via UHPLC-Q-TOF-MS: polyamine conjugates, alkaloids and sesquiterpene lactones. Cutin biosynthesis, alteration of membrane lipids and heightened radical scavenging ability related to the accumulation of phenolics and carotenoids were observed indicating the hormone-like activity of omeprazole [3]. Such findings introduce the possible plant protecting applications of external molecules, which might be useful if plant cultivation in difficult climatic conditions.
Biotic stress
The influence of bacteria, viruses, fungi, parasites, insects, weeds, and other plants on crops always trigger a metabolic response. This sometimes exert a positive effect on plant performance through the development of resistance [60]. GC-MS metabolomics enabled the profiling of tomato fruit volatile metabolites in response to the toxigenic fungi: Aspergillus niger, Aspergillus flavus, and Fusarium oxysporum. Control ripe fruits released oleic acid amide as the most significant metabolite, whereas infected fruit produced increased amounts of octadecenoic acid in all three infection cases. Additionally, ten metabolites were unique to A. flavus while A. niger and F. oxysporum had 4 metabolites unique to each of them [61]. Such findings can be helpful in establishing the disease biomarkers and prevention of disease outbreaks.
Identification of biomarkers of biological activity
The chemical constituents of fruits affect not only their taste and nutritional value but also their potential beneficial effects on human health. The most explored group of secondary metabolites of berry fruits are polyphenols.
As a result of metabolomic analysis supported by statistical calculations, the antioxidant and α- glucosidase and α-amylase inhibitory activity of kiwi fruit were correlated with the presence of polymeric procyanidins, flavonols (16 quercetin and kaempferol derivatives), phenolic acids (7 compounds) and one anthocyanin compound [62]. Antioxidant and anti-inflammatory activities of blueberries were strongly correlated with the presence of anthocyanins, flavan-3-ols, flavonols, phenolic acids, and resveratrol quantified with LC-IT-TOF-MS [63]. Chemical markers suitable for the standardization of antioxidant activity of Lonicera caerulea fruits were determined by ultra-performance liquid chromatography with electrospray ionization tandem mass spectrometry (UPLC-ESI-MS/MS) followed by statistical analysis. Cyanidin-3 -glucoside, rutin, chlorogenic and 3,5-dicaffeoylquinic acid were found to be the most suitable for quality control and to ensure consistent, reproducible biological activity [64]. LC-DAD-ESI-QTOF-MS investigation of non-anthocyanin phenolic compounds in juice and wine made from white bilberry fruits showed that phenolic acids were the most predominant, accounting for approximately 80%of the total phenolic content, with p-coumaroyl monotropeins and caffeic acid hexoside being the major compounds. Fermentation process did not influence the content of p-coumaroyl monotropeins, however, contributed to the hydrolysis of glucosides and the release of free caffeic acid in wine [65]. Also, UPLC-ESI-MS/MS with statistical analysis was applied to determine qualitative and quantitative cranberry biomarkers. Among the identified phenolic compounds hyperoside, quercetin, and procyanidin A2 were predominated and were selected as standardization markers of bioactivity [66]. In another study, biosynthetic pathway-based non-targeted metabolite profiling applied by ultra-performance liquid chromatography-quadrupole-time of flight (UPLC-Q-TOF) MS and ultra-high-performance liquid chromatography linear trap quadrupole tandem mass spectrometry (UHPLC-LTQ-MS/MS) analyses have led to a better knowledge of variations in the metabolite distribution among berry species, as well as accurate component information for future plant uses [67]. Table 1 indicates a summary of the literature data relevant to the topics mentioned above.
A summary of literature data on the application of metabolomics on berry quality monitoring
A summary of literature data on the application of metabolomics on berry quality monitoring
PCA-principal component analysis; OPLS-DA-Orthogonal Projections to Latent Structures Discriminant Analysis; PLS-DA-partial least-squares regression discriminant analysis; PLS-partial least-squares discriminant analysis; ROC-Receiver Operating Characteristic analysis; RDI-regulated deficit irrigation
The intense biotransformation of polyphenols by the host’s enzymes and gut microbiota results in a significant number of metabolites that might enter the systemic circulation. Correspondingly, the consumption of polyphenols in the diet has a direct impact on human health and disease due to interaction between food metabolome and endogenous host metabolome. Therefore, it is important to track the relationship of specific food intake and dietary patterns in humans with diseases through the disclosure of dietary biomarkers. Analytical instrumentation advancements now provide complete and simultaneous identification of metabolites in human biological samples such as serum, urine and feces [69–71].
The metabolic output of berry polyphenols in urine
Phenolic metabolites in human urine after a single dose of cranberry syrup have been identified by using UPLC-Q-TOF-MS. For this purpose, 2 male and 2 female healthy volunteers consumed 0.6 mL/kg of cranberry syrup and urine samples were collected before (0 h) the consumption and at time points, 2, 4, 6 h after the consumption of cranberry syrup. In total, 32 metabolites were tentatively identified, including phase I metabolites and phase II metabolites, such as methylated and glucuronide conjugated forms. One coumarin (scopoletin), free phenolic acid derivatives (dihydroxybenzoic acid, dihydroferulic acid 4-O-β-D-glucuronide, coumaroyl hexose, and caffeoyl glucose) and flavonols (myricetin, methoxyquercetin 3-O-galactoside, and quercetin) were the detected metabolites of cranberry syrup polyphenols in the human urine [72]. Similarly, in a very recent pilot study, polyphenol metabolites of wild blueberry drink were evaluated by using UPLC-Q-TOF-MS in 24 h urine before and after the 4-week intervention to investigate the effects of daily wild blueberry drink (containing 253 mg anthocyanins and 766 mg total polyphenols per day) ingestion for four weeks on cognition and mood in fifteen healthy primary school children. The results revealed that metabolic derivatives of eight classes of polyphenols were detected in the urine samples (pyrogallol derivatives, phenylacetic acid derivatives, valerolactone derivatives, benzoic acid derivatives, propionic acid derivatives, catechol derivatives, cinnamic acid derivatives, hippuric acid) and hippuric acid levels were significantly increased at the second week. The authors also suggested that hippuric acid might be a promising metabolite to look into when accounting the health and cognitive advantages in both adults and children [73]. In a dietary intervention study applied to 52 post-menopausal women, urinary metabolites of elderberry extract were identified by HPLC-Q-Trap-MS/MS after acute consumption or daily consumption for 12 weeks. According to the results, 17 phenolic acid and 11 anthocyanin conjugates were detected in the urine following acute anthocyanin-rich elderberry extract consumption. When compared to plasma metabolites, urine has been found to have higher sulfated, methylated and glucuronidated anthocyanin conjugates, phenylacetic acids, cinnamic acids such as sinapic acid and p-coumaric, 3,5-dihydroxybenzoic acid and 4-hydroxybenzyl alcohol [74]. In another study, urinary metabolites of ellagic acid from strawberries were not affected by food processing. To investigate the processing effect on the urolithin content of urine samples from 20 healthy volunteers by LC-MS/MS, they were subjected to a randomized study where 200 g of fresh strawberry or equivalent dose of strawberry puree were consumed with 2 weeks interval during 4 weeks. As a consequence, the amount of free ellagic acid content increased 2.5-fold due to processing; however, urolithin glucuronides in urine were not affected [75]. Despite the high polyphenolic content of fresh berries, their processing by-products also still involve significant amounts of phenolic compounds likely to include the one unidentified before. Recently, urinary metabolites from grape pomace extract were evaluated in an open, controlled, randomized and crossover trial where 6 male and 6 female volunteers consumed 500 mL/70 kg bodyweight grape pomace beverage after 8 hours of fasting. For the identification of metabolites, urine samples collected before (0 h) and 24 h after the consumption of beverage were analyzed with liquid chromatography coupled with an electrospray ionization hybrid linear ion trap quadrupole-Orbitrap-mass spectrometry (LC/ESI-LTQ-Orbitrap-MS) method. As a result, 41 phenolic components were detected in grape pomace primarily comprised of procyanidins, phenolic acids and flavonols, whereas 77 metabolites were identified in urine samples composed of glucuronides, sulfate derivatives and microbiota metabolites [76]. In a cohort study, samples from selected 475 subjects from a large cohort study with 36,994 participants between the years 1992–2000 were assessed in terms of urinary excretions of thirty-four dietary polyphenols. Relative concentrations in samples over the reference sample were determined by UPLC-ESI-MS-MS. Consequently, a correlation between berry consumption and p-coumaric acid and (+)-catechin in urine samples was found with the spearmen correlation coefficients of 0.20 and 0.19, respectively [77]. Further data from the literature were summarized in Table 2.
Identification of metabolites in human and animal biological samples in urine, blood and feces
Identification of metabolites in human and animal biological samples in urine, blood and feces
n.a.: not available
Plasma strawberry anthocyanins of 14 adults having BMI: 26±2 kg/m2 were characterized on the basis of meal timing where the subjects consumed a strawberry drink containing 12 g of freeze-dried strawberry powder 2 h before the breakfast, during the breakfast or 2 h after the breakfast. The blood plasma samples taken every hour for 10 h were analyzed by UHPLC-QqQ-MS/MS technique. The findings showed that when the strawberry drink was ingested, pelargonidin glucuronide was the most prevalent metabolite circulating in the blood of the individuals, independent of meal timing. Also, strawberry intake with or without a meal resulted in a peak plasma concentration of anthocyanins and their metabolic conjugates within 1–3 hours and when parent pelargonidin-based compounds were taken with a meal, they were found in the greatest concentration in the plasma [78]. Later on, the same group investigated the plasma metabolome of 38 healthy older adults after chronic consumption of 12 g of freeze-dried strawberry powder for 90 days. In all, twenty-one polyphenolic metabolites were measured in plasma, including three anthocyanins/metabolites, three urolithin metabolites, and fifteen phenolic acid metabolites. Pelargonidin glucuronide was found to be the most abundant of the anthocyanin metabolites, which is in full agreement with their early findings [79].
Anthocyanin bioavailability can be influenced by different factors such as the motility and permeability of the gastrointestinal tract as well as gut microflora. Among these variables, the food matrix also has a significant impact on anthocyanin bioavailability. In a recent study, co-digestion of blackberry anthocyanins with ethanol was evaluated in terms of its effect on the anthocyanin bioavailability. After analyzing the blood plasma samples with high-performance liquid chromatography with diode-array detection (HPLC-DAD) and ultra-performance liquid chromatography with photodiode array detection-quadrupole-time of flight mass spectrometry (UPLC-PDA-Q-TOF-MS), the results revealed that ethanol increases cyanidin-3-glucoside metabolism, allowing it to convert more easily into methylated derivatives (methyl-cyanidin-glucuronide and 3’-methyl-cyanidin-3-glucoside). This impact was stronger in overweight and obese people, whose Cy3glc metabolism appeared to be impaired [80]. In another study, plasma metabolome in female college students following cranberry juice intake was examined using a global ultra-performance liquid chromatography- quadrupole orbital ion trap- high resolution mass spectrometry (UHPLC-Q-Orbitrap-HRMS) technique. Endogenous and exogenous metabolites in plasma were observed to change after cranberry juice consumption. There was an increase in endogenous metabolites such as itramalic acid, hydroxyoctadecanoic acid, aconitic acid, vanilloylglycine, 4-acetamido-2-aminobutanoic acid, hippuric acid, 2-hydroxyhippuric acid, dihydroxyquinoline, and glycerol 3-phosphate as well as exogenous metabolites namely quinic acid, catechol sulfate, 3,4-dihydroxyphenyl ethanol sulfate, vanilloloside, coumaric acid sulfate, 5-(trihydroxphenyl)-gamma-valerolactone, ferulic acid sulfate, 3-(hydroxyphenyl)proponic acid, trihydroxybenzoic acid, and hydroxyphenylacetic acid [81]. Similarly, in another study investigating the plasma metabolome of cranberry juice polyphenols in young, healthy men by using UPLC-Q-TOF-MS, 60 metabolites including conjugated and non-conjugated phenolic acid derivatives and flavonoid derivatives were detected in plasma after the consumption of cranberry juice. Among these, hippuric acids (44.7%), catechols (22.4%), benzoic acids (14.7%), phenylacetic acids (10%), and cinnamic acids (7%) were the most abundant metabolites [82].
The metabolic output of berry polyphenols in feces
The gut microbiota affects the bioavailability of both parent polyphenols and their metabolites through participating in the metabolism of dietary polyphenols. For example, anthocyanins are catabolized by bacteria in the colon, resulting in the production of new phenolic compounds. These new compounds may be absorbed and have different bioactive characteristics than their parents, as well as they may have a significant impact on systemic or local health [83]. Fecal metabolites of aronia berry and aronia extract polyphenols following the administration to the diet of C57BL/6J mice were analyzed by using HPLC-DAD technique and protocatechuic acid and cyanidin were found to be detectable by the proposed method [84]. The impact of wild blueberry intake on the metabolism of anthocyanins in Sprague-Dawley rats was studied by Bo et al. (2010) [85]. Thirty-two rats were fed a control (C) or a wild blueberry-enriched diet (8%) for 4 or 8 weeks. LC-MS/MS was used to determine the anthocyanin profile in feces. Sixteen anthocyanins (3 of them were acylated) were identified in feces where peonidin-galactose, petunidin-arabinose, and malvidin-glucose-acetyl were determined as the most abundant anthocyanins. After 4 weeks of blueberry intake, the main phenolic acid found in feces was 3-4-diyhydroxybenzoic acid, but after 8 weeks, there was a substantial decrease, showing that this molecule may be converted to benzoic acid by the gut microbiota. Feces metabolites of pomegranate juice were analyzed after the consumption of 200 mL pomegranate juice per day by 12 heathy adults for 4 weeks. After supplementing with pomegranate juice, the concentrations of fecal phenolic metabolites including 3-phenylpropionic acid, catechol, hydroxytyrosol, and urolithin A increased significantly [86].
Determination of the effect of berry metabolites on gut microbiota
Having a balanced gut microbiota is a crucial component in leading a healthy life since they are thought to play an essential part in their host’s defense system [17]. Dietary phenolics and their metabolites, which cannot enter the systemic circulation, have a major impact on the intestinal environment via modulating the gut microbiota [96]. Although earlier polyphenol research focused on their antimicrobial properties, today the notion of polyphenols as possible prebiotics to modify the gut microbial composition is growing. Our gut hosts over a thousand “species-level” phylotypes, most of which are bacteria from only a few phyla that are commonly detected in feces. Nonetheless, a large portion of the microbiota observed in diluted fecal specimens cannot be cultivated on culture medium. Despite the variation in estimates between studies and between individuals, about 40–80 percent of the microscopic fecal counts are not recoverable by culture. However, the description of the whole repertoire of human gut microbiota and emphasizing the precedence of bacteria became feasible owing to culture-independent high throughput molecular biology methods [83, 97]. These molecular approaches entail the amplification of 16S rRNA genes (16S rDNA) from microbial DNA isolated from samples collected from specific habitats using Polymerase Chain Reactions (PCR) [98]. Commonly, it is thought that decreasing the Firmicutes/Bacteroidetes ratio and colonization of particular beneficial bacterial species protects the host from certain diseases [99]. However, some other genera such as Salmonella, Pseudomonas, Bacillus or Staphylococcus can be human pathogens. Recent evidence showed that phenolic compounds from berries and their metabolites generated in the human body have a discriminating effect on bacteria [99]. For example, berry polyphenols have been shown to produce prebiotic-like effects in the gut in animal experiments. Significant increase in Bifidobacterium populations have been rendered in rats fed with blackcurrants (FISH method) [100] and blueberry extracts (HTS [101] and FISH [102] methods). Similarly, the gut microbiome of diabetic mice was shown to be modulated by strawberry anthocyanins. After 10 weeks of 2.35%freeze-dried strawberry intake, researchers found a substantial rise in Bifidobacterium abundance by using 16S rRNA amplicon sequencing. However, the amount of Akkermansia, which was found to have anti-inflammatory effects, has decreased significantly [103]. Change of the gut microbiota by berry intake is further detailed in Table 3.
A summary of recent literature data on the modulation of gut microbiota by berry intake
A summary of recent literature data on the modulation of gut microbiota by berry intake
PCR: Polymerase chain reaction; DGGE: denaturing gradient gel electrophoresis; HTS: high throughput sequencing; FISH: fluorescent in situ hybridization. (+): Increase in the number of microorganisms, (–): Decrease in the number of microorganisms.
Food component analysis, food quality/authenticity evaluation, food intake tracking, and physiological follow-up in food intervention studies are only a few of the applications for metabolomics in food science and nutrition research. Thanks to the application of the metabolomics approach to berries, it is possible to improve breeding performance, accelerate genetic modifications, select stress-resistant plants, provide food quality, and standardize the crops depending on the biomarkers of biological activity. Dietary intervention studies provide a unique opportunity that allows exploring the response of human metabolism to berry consumption, especially in monitoring the change in disease biomarkers. The chemical composition of the berry fruits changes from a species to another, and considering the possible synergies between compounds and interactions with the fruit matrix, studying the impact of the whole berry rather than individual components is required. However, two limitations continue to restrict the potential effect of metabolomics: 1-) technology and 2-) databases. Major advancements are required to make metabolite identification and quantification technologies more durable, automated, and comprehensive. Many additional references, spectral or chromatographic databases on food metabolites need to be produced and should be open access.
Footnotes
Acknowledgments
G. Catalkaya and E. Capanoglu would like to acknowledge Istanbul Technical University, Scientific Research Projects (BAP) Unit (Project No: MDK-2019-41928).
Author contributions
The authors confirm contribution to the paper as follows: Study conception and design E. Sieniawska; supervision: E. Capanoglu; draft manuscript preparation: G. Catalkaya, M. Maciejewska-Turska, G. Kai; review and editing the original draft: G. Catalkaya, E. Sieniawska, E. Capanoglu.
Conflict of interest
Dr. Capanoglu and Dr. Sieniawska are members of the Editorial Board of this journal but were both not involved in the peer-review process and neither had access to any information regarding the peer review of their paper.