
Abstract
Iron chelators have been utilized clinically to treat patients with iron overload conditions. There is a growing body of evidence linking iron dysregulation and reactive oxygen species (ROS) overproduction as underlying factors in Cystic Fibrosis (CF) disease. The chronic inflammation can lead to progressive airway destruction. Alleviation of this chronic inflammation is a potential target for CF treatment and thus, this research investigated the dose-response effects of DIBI, a novel iron chelator, on inflammation in CF nasal epithelial cells. Polarized CF cells were stimulated with, lipopolysaccharide (LPS), co-treated with DIBI (LPS+DIBI), or DIBI alone (DIBI). We demonstrated that DIBI modulated the release of IL-6 and IL-8 in CF cells in a dose-dependent manner. Reduction of extracellular iron with the lower doses of DIBI (25 and 50μM), increased IL-6 secretion in non-induced cells. LPS challenge increased IL-6 and IL-8 secretion which was suppressed by high dose (200μM) DIBI administration. This study demonstrates the therapeutic potential of iron chelation therapy to treat the dysregulation of the immune response in CF patients.
Introduction
Iron chelators have been utilized clinically to treat patients with iron overload conditions such as hemochromatosis and patients who require frequent blood transfusions. In this study, DIBI, a novel, water-soluble, and non-toxic iron chelator was used. It has a polyvinylpyrrolidone (PVP) backbone with nine broadly distributed MAHMP pyridinone chelating groups and one molecule of DIBI strongly binds three molecules of iron with full hexadentate coordination [1]. DIBI has been found to modulate inflammatory responses including lowering excess cytokine production in animal models of both sterile endotoxin-induced inflammation and in polybacterial peritoneal sepsis [2].
Cystic Fibrosis (CF) is an autosomal recessive genetic disorder that is caused by mutations in the gene encoding the Cystic Fibrosis Transmembrane Regulator (CFTR) protein, a cAMP-activated chloride channel [3]. Given the dual role that CFTR plays in the directional transport of ions and in the regulation of fluid secretion and absorption, CFTR dysfunctions lead to disturbances in airway surface liquid homeostasis and inadequate hydration of respiratory tract secretions [4]. This leads to accumulation of viscous mucus and impaired mucociliary clearance within the airways of CF patients [5, 6]. Therefore, patients with CF have the risk of recurrent infections in their airways.
CFTR dysfunction not only impairs the mucociliary clearance, but it also affects immune functions. CFTR mutations have been associated with baseline constitutive activation of pro-inflammatory signaling pathways in the absence of infection [7]. Despite this heightened immune response including excess iron-dependant production of reactive oxygen species (ROS), pathogen clearance is not effective. The resulting chronic inflammation can lead to progressive airway destruction characterized by bronchiectasis and emphysema [8], and eventually, respiratory failure [9, 10]. Alleviation of this baseline inflammation is a potential target for CF treatment.
There is a growing body of evidence showing iron dysregulation and ROS overproduction to be an underlying dysregulation in CF [11]. Continuous activation of NF-κB also leads to an increased amount of ROS through the activation of NADPH oxidase of neutrophils contributing to the oxidative stress and lung injury seen in CF patients [12, 13]. In addition to the important quantities of ROS released by neutrophils within the bronchial lamina, bronchial ciliated and alveolar type II epithelial cells are also able to contribute to the production of ROS through two isoforms of NADPH oxidase expressed on the apical compartment [13]. It has been further shown that before the onset of chronic bacterial colonization, epithelial DUOX continuously releases H2O2 and is thus a predominant source of ROS with respect to the NADPH oxidase from phagocytes [14]. Although the production of ROS might otherwise be an effective antimicrobial strategy, it has been observed, clinically that the resolution of the underlying infection does not normalize markers of oxidative stress within the CF lung [15]. This suggests that within the context of CF, ROS may be less effective for killing pathogens. However, excess airway ROS could be an important source of oxidative stress, and chronically damage the pulmonary epithelium.
Increased levels of iron and iron-related proteins have also been observed locally in the respiratory sputum and secretions of CF patients [16], which increases the risk of infection with inhaled pathogens. Thus, this research sought to investigate the dose-response impact of DIBI on inflammation in CF nasal epithelial cells. Given the pivotal role of iron in innate immune response, we hypothesized that iron chelation modulates excess inflammation and analyzed IL-6 and IL-8 inflammatory mediators release.
Material and methods
Cell cultures
CF nasal epithelial cell line (JME/CF15), homozygous for the ΔF508 mutation in the CFTR gene was generated as previously described [17]. For cell culture, the JME/CF15 cells were grown in Dulbecco’s Modified Eagle’s Medium supplemented with 10% FBS; 0.088% transferrin, 0.088% T3, 0.0088% epithelial growth factor (EGF); 1.76% hydrocortisone; 0.044% insulin, 0.088% epinephrine; and 0.176% adenine. The cells were grown until they reached confluence in T25 flasks (Corning, Corning, NY) at 37 °C in a 5% CO2 atmosphere. Cell culture media and supplements were purchased from GIBCO (Burlington, ON, Canada) and Sigma Aldrich (St. Louis, MO, US). Upon confluency, the cell cultures were detached with 0.1% trypsin-EDTA and were seeded into transwells (24 mm diameter with 0.4μM diameter pores; Corning Life Sciences, Acton, Mass.) with both apical and basolateral compartments filled with 2 mL of the described culture media. After one-week, and the formation of tight cell monolayers, the apical medium was removed. The monolayers were then maintained under air-liquid interface (ALI) conditions to achieve polarization and differentiation. Following the removal of the apical medium, the medium in the basolateral compartment was changed every two days until the cells achieved of a dry apical surface and were deemed ready for stimulation.
Cell stimulation
Following cell polarization and tight junction (TJ) formation, transwells were designated as either control (medium), stimulation with bacterial toxin lipopolysaccharide (LPS), co-treatment with DIBI (LPS+DIBI) or DIBI alone (DIBI). Stimulation with LPS (from Escherichia coli, serotype O26:B6; Sigma-Aldrich, Oakville, ON) was performed in fresh phenol red-free medium at concentrations of 100, 200, or 300 ng/mL for bidirectional stimulations of the cells. Once optimal LPS stimulation conditions were established (200 ng/mL dose for 24 hours), the experimental groups were tested: control (medium only), LPS (200 ng/mL), LPS+DIBI (25, 50, 100 or 200μM), DIBI only (25, 50, 100 or 200μM). The appropriate stimulation medium was applied to both the apical and basolateral surfaces at volumes of 0.5 mL and 2.5 mL, respectively. The cells were left to incubate for 24 hours in a 37 °C and 5% CO2 environment before cell harvesting.
Determination of cytokine secretion
Enzyme-linked immunosorbent assay (ELISA) kits were used to measure the IL-6 and IL-8 levels in the apical and basolateral cell culture supernatants. Monoclonal human IL-6 and IL-8 antibodies (Invitrogen, Carlsbad, CA) were used. The cytokine levels were normalized to the total protein concentration of the lysed cells, as measured via the Bradford Colorimetric Assay (BCA).
Statistical analysis
Data were analyzed using Prism 8 (GraphPad Inc., La Jolla, CA, USA). Normality of data was confirmed by Kolmogorov-Smirnov test. Data was analyzed using unpaired t-tests, or one-way ANOVA followed by the Newman-Keuls test for comparison of three or more groups. All sample analyses were performed in technical duplicates with the cytokine levels normalized to their protein content. Data was expressed as a mean value±standard deviation (SD). N represents the number of independent experiments. Significance was assumed at P < 0.05.
Results
IL-6 and IL-8 release in response to DIBI
To investigate the influence of DIBI iron chelator on cytokine secretion JME/CF cells were incubated with DIBI at 25, 50, 100, 200μM for 24 hours. Control cells secreted on average 196 pg/mL and 50 pg/mL IL-6 into the apical and basolateral compartments, respectively. These baseline control values were then used to measure relative changes as induced by the various treatments. Cells incubated with the lower concentrations of DIBI (25 and 50μM), exhibited approximately a 2.5-fold increase in apical secreted IL-6 over control levels (Fig. 1, part A). Interestingly the higher concentrations of, DIBI (100μM and 200μM) did not stimulate apical IL-6 secretion, with IL-6 concentrations remaining close to the control levels. Basolateral IL-6 secretion was also significantly increased with the lowest concentration of DIBI (25μM) (Fig. 1, part B), while DIBI at 50, 100, or 200μM did not appear to influence basolateral IL-6 secretion, over control levels.
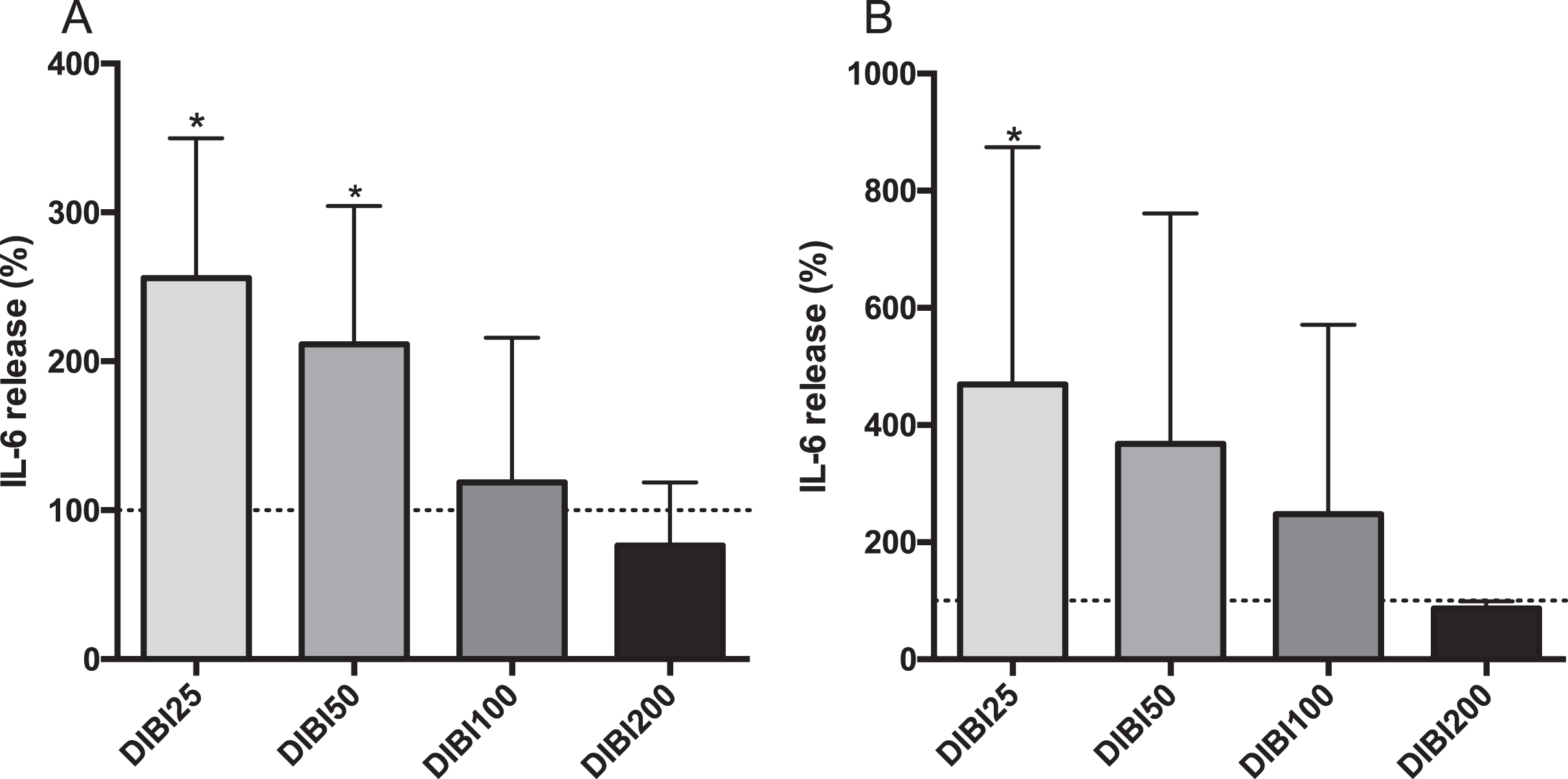
IL-6 secretion from CF15 cells in response to DIBI treatment. Apical (A) and basolateral (B) IL-6 secretions of polarized JME/CF15 cells. Upon polarization, CF15 cells were treated with DIBI (25, 50, 100, 200μM) or medium alone for 24 hours. The supernatant IL-6 secretion level was normalized to total protein concentration and the data represents the percentage change from control levels (medium only, dashed lines) corresponding to average values of 196 pg/mL and 50 pg/mL IL-6 in apical and basolateral compartments, respectively. Data is presented as mean±SD, (N = 3-4 independent experiment per group). *P < 0.05 vs control. DIBI25:25μM; DIBI50:50μM; DIBI100:100μM; DIBI200:200μM.
In the case of IL-8 secretion, control cells secreted on average 64 pg/mL and 32 pg/mL IL-8 into the apical and basolateral compartments, respectively. None of the treatments (DIBI 25, 50, 100 or 200μM) induced significant IL-8 changes over control levels in either the apical or basolateral compartments (Fig. 2).
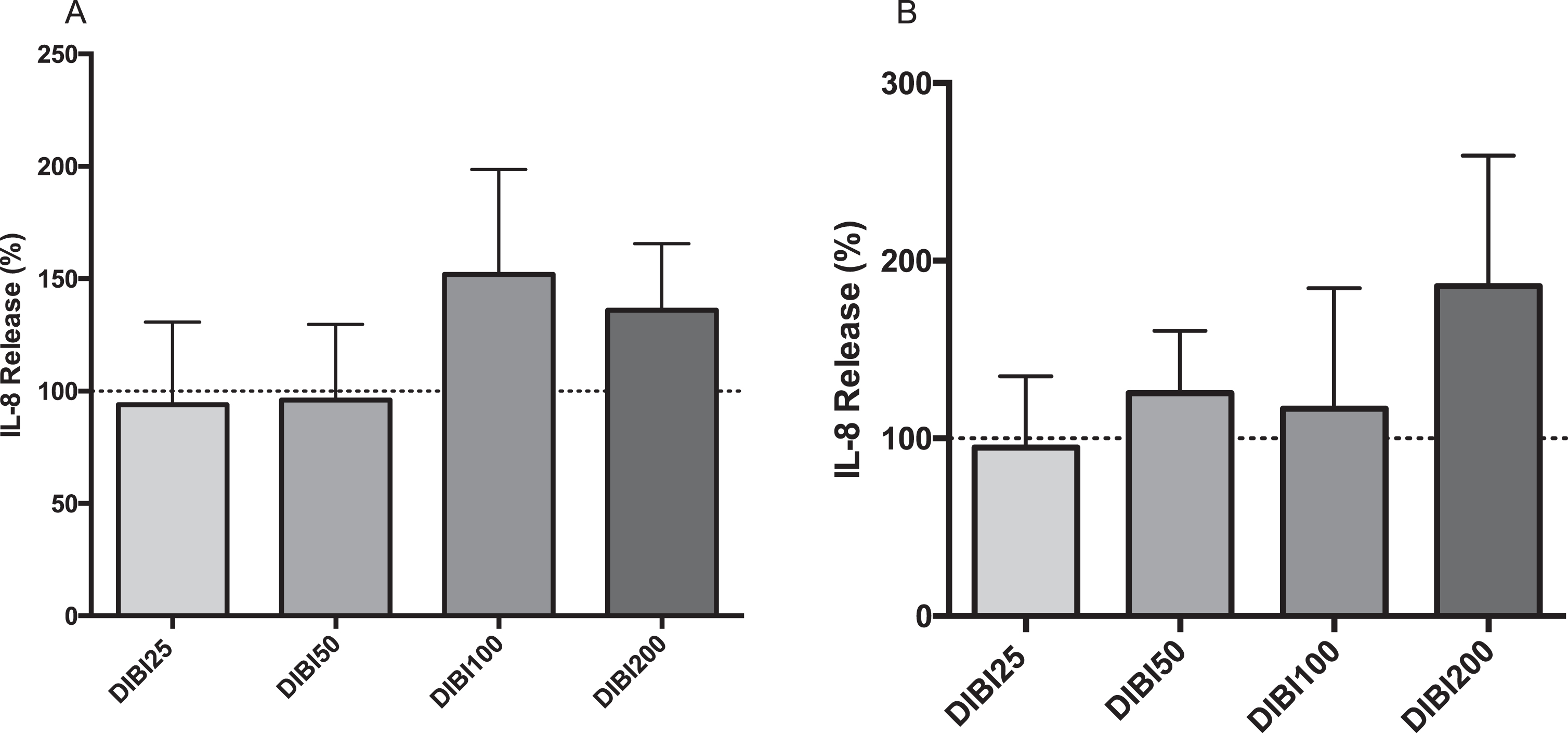
IL-8 secretion from CF15 cells in response to DIBI treatment. Apical (A) and basolateral (B) IL-8 secretions of polarized CF15 cells. Upon polarization, CF15 cells were treated with DIBI (25, 50, 100, 200μM) or medium alone for 24 hours. The supernatant IL-8 secretion level was normalized to total protein concentration and the data represents the percentage change from control level (medium only, dashed lines) corresponding to average values of 64 pg/mL and 32 pg/mL IL-8 in apical and basolateral compartments, respectively. Data is presented as mean±SD, (N = 3-4 per group). *P < 0.05 vs control. DIBI25 : 25μM; DIBI50 : 50μM; DIBI100 : 100μM; DIBI200 : 200μM.
JME/CF15 cells were stimulated with various doses of LPS (100, 200, 300 ng/mL) for 24 hours to determine the optimal LPS concentration to induce an inflammatory stimulus. Compared to controls, LPS at 200 ng/mL increased IL-6 secretion the highest in both apical (Fig. 3, part A), and basolateral (part B) compartments. IL-6 concentrations in the apical compartment were > 500 pg/mL and around 100 pg/mL in the basolateral compartment, representing >2.5-fold and 2-fold increases over controls, respectively. Based on these results, subsequent LPS stimulation experiments employed 200 ng/mL LPS with 24 hours exposure.
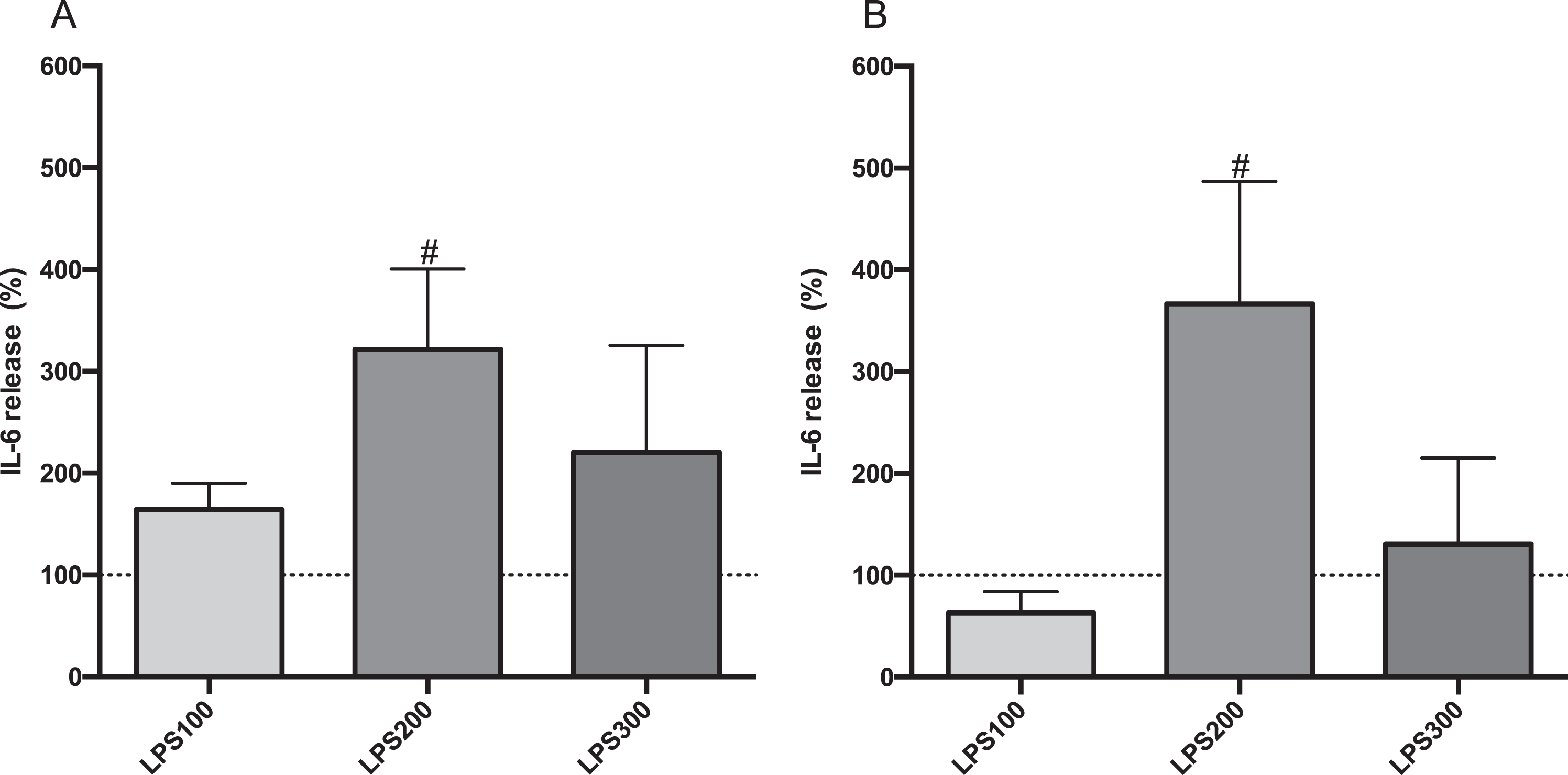
IL-6 secretion from CF15 cells’ in response to LPS stimulation. Apical (A) and basolateral (B) IL-6 secretions of polarized CF15 cells. Upon polarization, CF15 cells were stimulated with lipopolysaccharide (LPS) at 100, 200, or 300 ng/mL for 24 hours. The supernatant IL-6 secretion level was normalized to total protein concentration and the data represents the percentage change from control level (medium only, dashed lines) corresponding to average values of 196 pg/mL and 50 pg/mL IL-6 in apical and basolateral compartments, respectively. Data is presented as mean±SD, (N = 3-4 independent experiments per group). *P < 0.05 vs control. LPS100: 100 ng/mL; LPS200: 200 ng/mL; LPS300: 300 ng/mL.
Apical IL-6 secretion significantly increased in LPS-stimulated CF15 cells with DIBI treatments of 25μM and 50μM (LD25 and LD50), compared to LPS level (Fig. 4, part A). LPS + DIBI 100μM treatment (LD100) did not affect IL-6 secretion compared to LPS-challenged cells. On the other hand, cells treated with LPS + DIBI 200μM (LD200), showed reduced IL-6 secretion. For the basolateral compartment, LPS-induced cells treated with DIBI 25, 50, and 100μM showed no changes in IL-6 secretion, while DIBI 200μM reduced IL-6 basolateral secretion significantly (Fig. 4, part B).
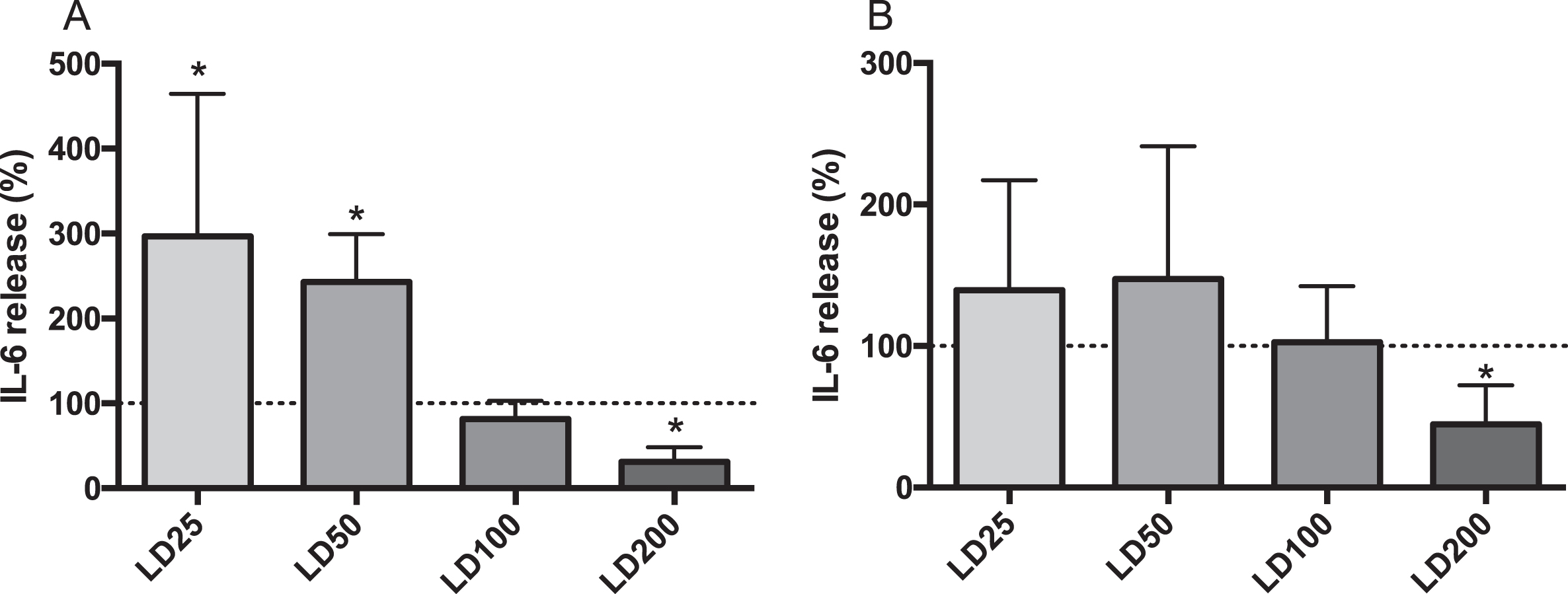
Effects of DIBI on LPS-stimulated IL-6 secretion from CF15 cells. Apical (A) and basolateral (B) IL-6 secretions of polarized CF15 cells. Upon polarization, CF15 cells were challenged with lipopolysaccharide (LPS; 200 ng/mL) and treated with DIBI (25, 50, 100, 200μM) or media alone for 24 hours. The supernatant IL-6 secretion amount was normalized to total protein concentration and the data represents the percentage change from control level (medium only, dashed line) corresponding to average values of 526.5 pg/mL and 99.53 pg/mL IL-6 in apical and basolateral compartments, respectively. Data is presented as mean±SD, (N = 3-4 independent experiments per group). *P < 0.05 vs LPS. LD25: LPS+DIBI 25μM; LD50: LPS+DIBI 50μM; LD100: LPS+DIBI 100μM; LD200: LPS+DIBI 200μM.
LPS induced secretion of IL-8 into both the apical and basolateral compartments. LPS + DIBI at 25, 50, or 100μM had no significant effect on IL-8 secretion into the apical compartment compared to LPS alone (Fig. 5, part A). However, the highest dose of DIBI (200μM) significantly reduced apical IL-8 secretion by 20%. A similar response was observed in the basolateral compartment, where both DIBI 100 and 200μM reduced IL-8 secretion significantly by 24% and 42%, respectively. (Fig. 5, part B).
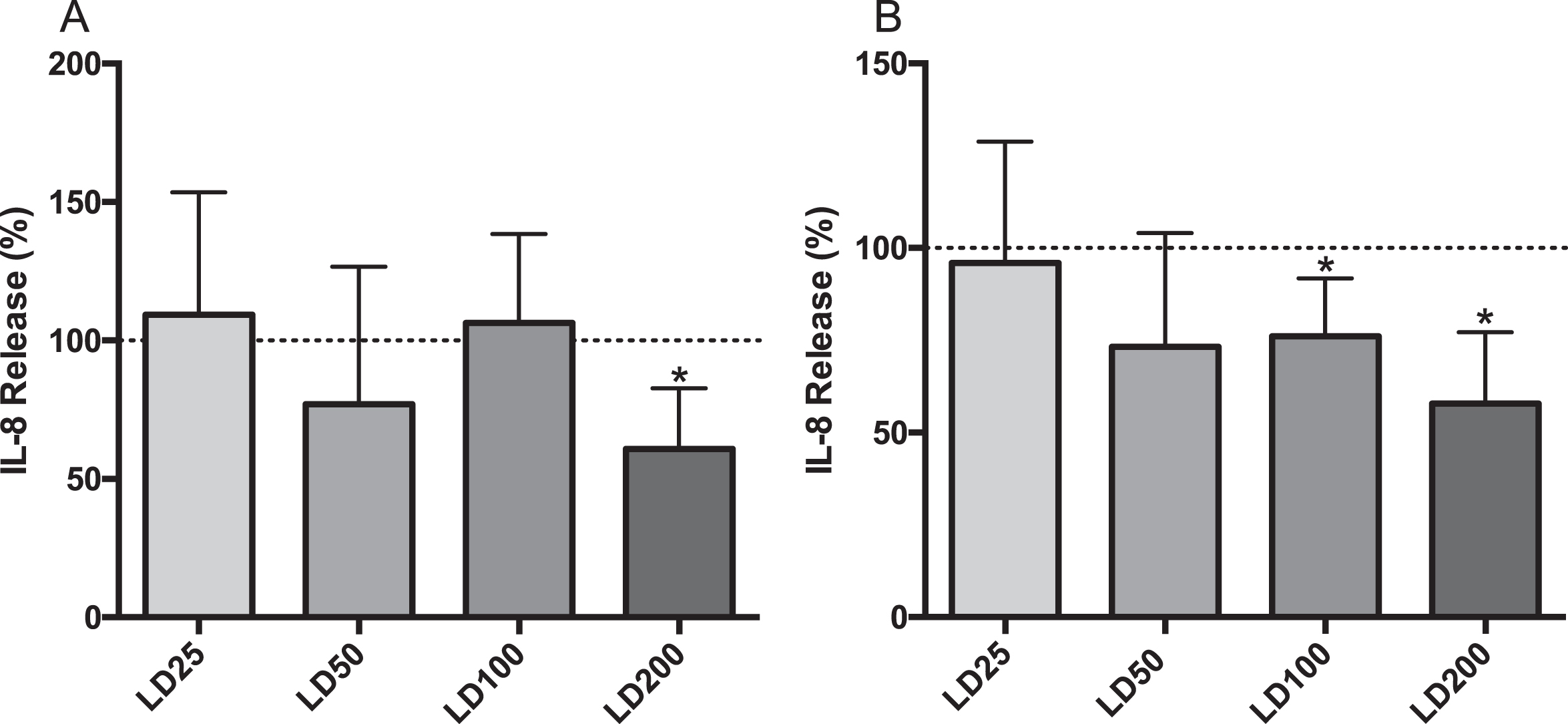
Effects of DIBI on LPS-stimulated IL-8 secretion from CF15 cells Apical (A) and basolateral (B) IL-8 secretions of polarized CF15 cells. Upon polarization, CF15 cells were challenged with lipopolysaccharide (LPS; 200 ng/mL) and treated with DIBI (25, 50, 100, 200μM) or media alone for 24 hours. The supernatant IL-8 secretion amount was normalized to total protein concentration and the data represents the percent change from control level (medium only, dashed line), corresponding to average values of 79.5 pg/mL and 54 pg/mL IL-8 in apical and basolateral compartments, respectively. Data is presented as mean±SD, (N = 3-4 independent experiments per group). *P < 0.05 vs LPS. LD25: LPS+DIBI 25μM; LD50: LPS+DIBI 50μM; LD100: LPS+DIBI 100μM; LD200: LPS+DIBI 200μM.
Iron dysregulation in CF patients has been shown to impact the chronic inflammation and bacterial infection that is observed in these patients [16, 18–20]. Thus, iron chelation presents a novel treatment to attenuate chronic airway inflammation in CF patients [21]. Iron and iron-related proteins levels are elevated in bronchoalveolar lavage fluid, macrophages, and explanted lungs of CF patients [22]. Further contributing to the increase of P. aeruginosa growth and biofilm formation in the airways, bronchial epithelial cells with ΔF508-CFTR mutations have increased iron release from their apical membrane. Interestingly, iron chelation was able to reduce this biofilm formation [23], supporting the therapeutic application of iron chelation. In the current study, we demonstrated that the novel iron chelator, DIBI, modulates the release of IL-6 and IL-8 in a dose-dependent manner, in human nasal epithelial cells JME/CF15, homozygous for the F508del-CFTR mutation. Lower doses of DIBI (25 and 50μM), on its own increased IL-6 secretion. Additionally, LPS challenge increased IL-6 and IL-8 secretion which was suppressed by high dose (200μM) DIBI administration.
We have established a working hypothesis consistent with both our present findings as well as with other reports as shown in Fig. 6. Reduction of extracellular iron concentration by low dose iron chelation could act as a signal of bacterial invasion with iron consumption. This signal might then induce the activation of nuclear factor (NF) pathways, such as p38 or ERK1/2, initiating pro-inflammatory cascades, with release of IL-6 and IL-8 as shown in respiratory [24] and intestinal epithelial cells [25] consistent with our findings. These various findings then suggest that iron homeostasis disruption by low dose DIBI or actual infection might act as a pro-inflammatory signal to warn of iron-consuming microbial invasion. Therefore, reduction of extracellular iron concentrations by (low) dose DIBI which leads to an increase in NF-κB P65 signal activation could be related to cellular bacterial infection sensing mechanisms (Fig. 6, part A).
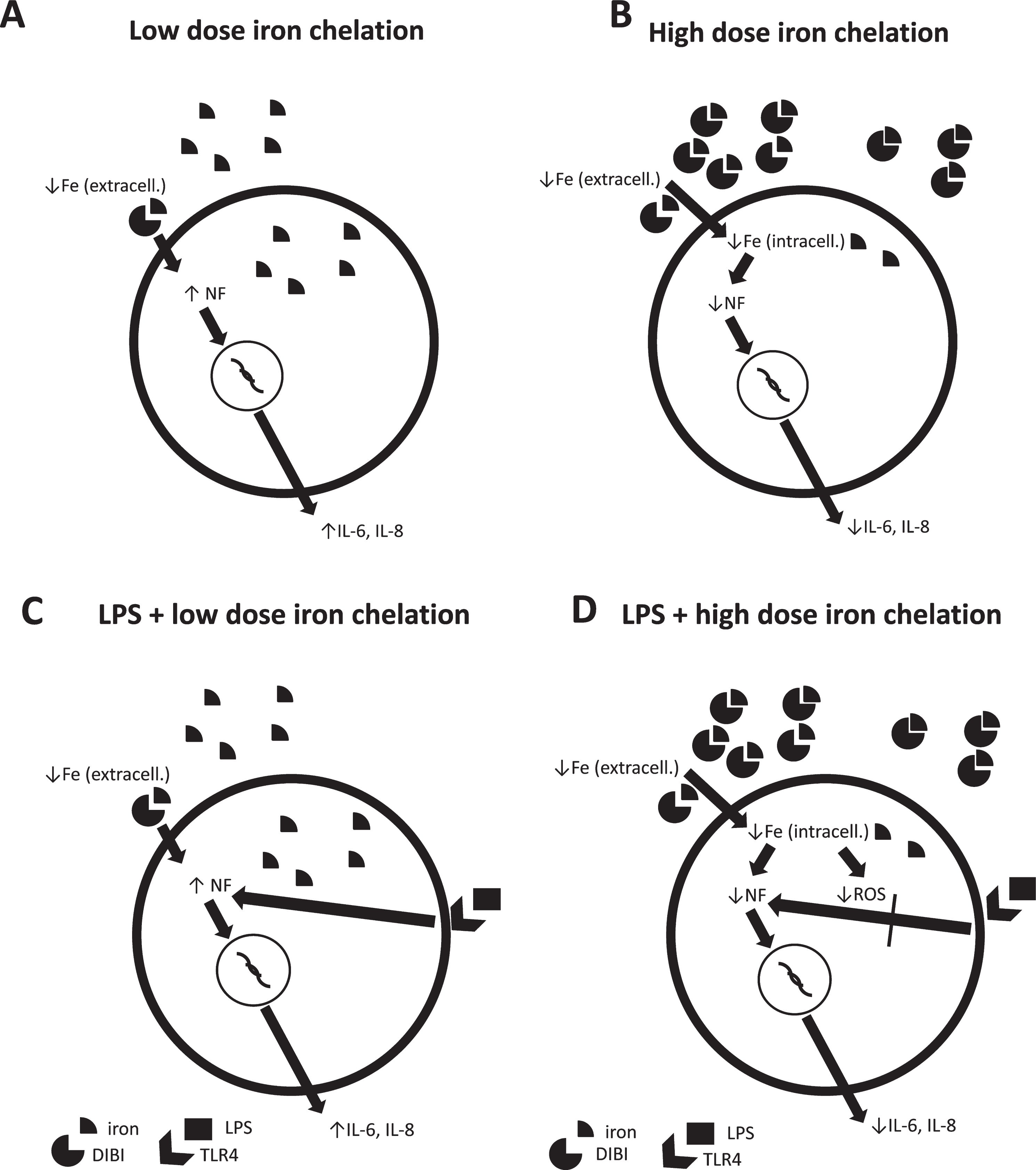
Working hypothesis of dose response effects of DIBI on pro-inflammatory cytokine responses in normal and LPS-induced CF15 cells. A) Low dose iron chelation reduces extracellular iron concentration. In response, nuclear factor (NF) pathways are initiated resulting in increased release of IL-6 and IL-8. B) High dose iron chelation does not only cause extracellular but also reduction in intracellular iron levels. This then results in decreased intracellular NF synthesis and reduced IL-6 and IL-8 release. C) LPS binds to toll-like receptor (TLR-) 4 and causes activation of NF pathways that support the IL-6 and IL-8 synthesis. D) High dose iron chelation with LPS-induced cells suppresses excess IL-6 and IL-8 release by direct (iron-related reduction of protein and NF synthesis) and indirect (reduction of ROS-induced upregulation of IL-6 and IL-8) mechanisms due to reduced intracellular iron levels.
Higher dose iron chelation might not only reduce extracellular iron levels but also reduce intracellular levels of free iron. In this study, at basal conditions the CF15 cells were not sensitive to higher doses of iron chelation (100 and 200μM). The reduction in iron bioavailability by iron chelation could lower intracellular NF synthesis and subsequently reduce IL-6 and IL-8 release [26]. However, when stimulated with LPS, higher DIBI doses, which most likely mobilized some intracellular iron to extracellular environment, suppressed LPS-induced IL-6 and IL-8 secretion. In line with this finding, high doses of DIBI did not increase nuclear localization of NF-κB P65 with LPS unlike treatment with low dose DIBI in preliminary experiments (data not shown). This suggests the reduction of the inflammatory phenotype of these cells to protect in response to bacterial challenge. Several mechanisms might play a role here (Fig. 6, part D). Iron chelation might result in reduction of iron-dependent protein tyrosine phosphatase activity [27] and reduced nuclear factor synthesis [28] with reduced downstream effects on cytokine release. Also, the decreased ROS-induced upregulation of IL-6 and IL-8 due to reduced iron bioavailability for ROS generation could play a role [29]. These results, taken together highlight the dynamic role of iron and iron chelation on the immune response.
We did not observe IL-8 changes with low dose DIBI (25 and 50μM) administration, despite an in increase in NF-κB P65 signal activation with low dose (25μM; preliminary data). This might be explained by the high baseline activation of pro-inflammatory pathways in CF15 cells. Carrabino et al. measured the activity of NF-κB and IL-8 secretions of explant cultures of nasal epithelial cells from CF and non-CF patients at baseline and with P. aeruginosa/IL-1β/H2O2 co-stimulation [30]. This study demonstrated the higher baseline activation of NF-κB and subsequently IL-8 secretion in the CF cells, compared to non-CF cells. In a separate study by Massengale et al., [31] airway epithelial CF cells had a lower induction of IL-8 mRNA and protein levels in response to IL-1β stimulation, as compared to its isogenic CFTR-corrected control, further emphasizing IL-8 secretion dysregulation in CF cells.
In our study the CF15 cells were polarized, enabling differential measurement of IL-6 and IL-8 secretion on both apical and basolateral compartments. Direct comparison of the revealed higher IL-6 and IL-8 secretion on the apical side. This polarized response creates a chemotactic gradient, promoting trans-epithelial migration of leukocytes into the apical compartment from basolateral compartment. This finding is consistent with previous studies which demonstrated a polarized response in human airway epithelial cells during inflammation [32–34]. In the context of CF, where high neutrophil recruitment and activation are responsible for inducing damage, the reduction of IL-6 and IL-8 observed by higher dose of DIBI could be beneficial by alleviating this response.
Overall, the highest dose of DIBI studied (200μM) did not induce IL-6 and IL-8 release in CF15 cells at basal condition. However, it lowered LPS-induced IL-6 and IL-8 secretion, displaying anti-inflammatory properties. Therefore, this study shows a therapeutic potential of iron chelation therapy to treat the dysregulation of the immune response in CF patients.
Conflict of interest
BH has a beneficial interest in Chelation Partners Inc.
Footnotes
Acknowledgments
This work was supported by the Cystic Fibrosis Foundation, Cystic Fibrosis Canada and NSERC discovery grant to VC. MA was supported by Killam Pre-doctoral Scholarship (Canada). The authors thank Dr. David Hoskin and Javad Ghassemi for their help and expertise with cytokine measurements. Additionally, the authors thank Chelation Partners Inc. for providing DIBI.