
Abstract
Background:
The ability of healthy individuals to detect biological motion by using a small number of moving points is well established in animals and humans. Perception of human movements may depend on internal models that drive self-generated movements and influence motion discrimination (Reed CL et al. 1995 and 2007). As a person’s motor repertoire deteriorates, the accuracy of these models may also decrease.
Objective:
Determine if people with symptomatic Huntington’s disease (HD) have difficulty perceiving movements.
Methods:
In this study point-light displays were created with a Vicon Motion Capture System by recording one individual with (impaired) and one individual without (healthy) Parkinson’s disease using a 13 joint marker set. Participants were asked to distinguish between three movements and determine if the movement was impaired or healthy. The ability of participants with and without HD to distinguish movement patterns and the time to perception were recorded.
Results:
Analyses found participants with HD had a decreased ability to correctly detect movements and point-light image type. The stair climbing motion showed the largest effect as participants with HD had more difficulty correctly identifying both the movement and whether it was impaired or healthy. In addition, the participants without HD showed an improvement as trials progressed which could not be observed in the HD cohort.
Conclusions:
As people with symptomatic HD have difficulty perceiving movements further investigations using point-light displays should be done to determine if these impairments might serve as an easily administered, non-invasive marker of disease state.
INTRODUCTION
Huntington’s disease (HD) is an autosomal-dominant, progressive neurodegenerative disorder characterized by impaired motor function, including choreatic movements and abnormal eye movements [1], as well as cognitive decline and behavioral changes [2]. These symptoms are the result of an abnormal length of the huntingtin protein [2] that leads to brain atrophy especially affecting the striatum [3, 4]. As there is growing evidence that the basal ganglia play a key role in complex visual object recognition, atrophy in this region leads to deficits in visual perception/function [5, 6]. Some evidence suggests that HD also affects visual motion perception [7].
Biological motion perception is an innate ability of both animals [8, 9] and humans [10] to identify different types of motion accurately using only point-light displays [10]. To create these displays, small lights are attached to the head and main joints of the body and filmed during movement. When healthy observers without previous training view these point-light images moving on a dark background, they easily identify the moving point-lights as a person and recognize the motion. When the motion stops, however, viewers no longer perceive the point-light images as a person. Using point-light displays, later investigators demonstrated that infants exhibit a predisposition to look at biological motion [11] and the ability to recognize biological motion reaches adult levels by five years of age [12].
The motor dysfunction of HD may create abnormal biological motion perception in people with HD. Several studies argue that perception of biological motion operates via a template matching approach [13, 14] in which the brain compares the point-light stimuli to neuronal templates of human movements [15]. In this model, the observer's self-generated movements influence motion discrimination [16, 17]. The deficient somatosensory feedback [18] and impaired memory of limb position [19] characteristic of HD may lead to an inaccurate internal model of normal voluntary movement and deterioration of the neuronal template may occur [15]. Thus, people with symptomatic HD might not only lose the ability to perform movements accurately but also have difficulty perceiving healthy movements. Consistent with this idea, a recent study demonstrated that individuals with Parkinson’s disease (PD) exhibit impaired biological motion perception when observing point-light walking [20].
Activation patterns of mirror neurons in the parietal and premotor cortex suggest a strong link between perception and action [21]. In addition, there is increasing evidence that internal representations of perception and action may overlap [22]. Given the potential relationship between movement performance deficits and impaired perception in people with HD, this study aims to determine if HD impairs biological motion perception compared to persons without HD.
MATERIAL AND METHODS
Participants
Twenty-three individuals with clinically- or genetically-confirmed HD (10 Male, aged 50.6± 13.2 years) and twenty individuals without HD (11 Male, aged 50.8±14.2 years) participated in this study. A neurologist experienced in HD examined all individuals with HD and thirteen of the twenty participants without HD using the Total Motor Score (TMS) of the Unified Huntington Disease Rating Scale (UHDRS) [23]. TMS is the sum of 15 individual motor items, each rated from 0 to 4 (maximum score of 124), with a higher score indicating greater impairment. Participants with HD had an average TMS of 29.3 (±23.4) points while the thirteen control participants had a mean TMS of 1.6 (±2.2) (Table 1a,b). For practical reasons we did not include persons suffering from major depression as reported by the neurologist performing the UHDRS and routine psychiatric assessments during out-patientvisits.
Demographics
Age, TMS (total motor score) and composite ocular pursuit, saccade initiation and saccade velocity scores on the UHDRS in the horizontal and vertical direction. While participants were similarly aged, those with HD displayed greater motor impairment with slowed ocular pursuit and initiation. Data of the thirteen participants examined by a neurologist. **p < 0.01, ***p < 0.001.
Demographics of the HD group
Age of onset, number of premanifest participants, disease duration and medication intake
Video creation
We created point-light displays of biological motion with the VICON MX (©Vicon Motion Systems Ltd. UK) using a 10 Vicon-MX camera system to track the position of infra-red reflective markers attached to landmarks on the recorded body. A dedicated motion analysis computer received signals from all the cameras and automatically tracked the markers to establish highly accurate 3D trajectories. Subjects had markers placed on C7 and bilateral acromia, greater trochanters, lateral epicondyles of the femur and humerus, ulnar heads, and lateral malleoli. Three motions were recorded from two volunteers, one a healthy 45-year-old female and the other a 61-year-old female with Hoehn and Yahr stage III Parkinson’s disease tested while off medications treating movement impairments: walking back and forth along a 10 meter path (WALK), walking up and down stairs (STAIRS), and rising from sitting down in a chair to standing up (STS). In addition to these three movement patterns, the healthy volunteer performed jumping jacks to be used as practice trials for the participants.
Experimental procedure
Prior to any data collection all participants provided Informed Consent according to the procedures set forth by the Declaration of Helsinki. Subjects were seated comfortably at a standard height table. A Hewlett Packard ZBook Workstation with a 15.6-inch monitor positioned approximately 50 cm from the edge of the table was used to display point-light images. At a distance of 50 cm and a screen width of 31.7 cm, the visual angle of the screen was 35.2 degrees. If participants had corrective lenses they were allowed to wear those during the test. The point-light motions were white dots moving on a black background. Each biological motion dot was 16 pixels (Fig. 1A), subtending 0.15 degs [°] of visual angle at the typical viewing distance. In addition to the motion point-lights, 400 white dots consisting of 26 pixels (5x5 squares with an extra pixel on the side) were displayed on the screen (Fig. 1B). These dots moved randomly to create visual background noise. It was necessary to increase the dot size of the background noise because the biological motion dots moved slightly slower making them appear brighter. By increasing the dot size of the background noise more light was added to each point countering this effect. The image was refreshed at 62.5 Hz.

Point-light images of biological motion (A) alone and (B) embedded in 400 random moving points. Points have been slightly enlarged for graphic clarity.
After allowing participants to choose between a track pad and mouse to navigate the program, a screen invited participants to practice up to three test trials showing point-light motion of the healthy subject performing jumping jacks. Apart from programmed catch trials, each trial began with the point-light motion embedded in 400 randomly moving dots. Over 32000 ms, the number of randomly moving dots decreased linearly until only the point-light motion pattern was left. The subject’s instructions were to push the mouse button or track pad as soon as they recognized the movement pattern and determined whether the observed motion was healthy or impaired. The test trials served to acquaint participants with the procedure and demonstrate that they could identify motion from point-lights. These three trials were not included in the data analysis. Every participant completed 42 trials randomly presenting different movement patterns of the healthy and impaired motion. The point-light motion also changed randomly from a front to side view with 93% of the motion patterns upright and three catch trials without any visual background noise. The time taken to stop the motion and the number of noise dots remaining on the screen were recorded. After pushing a mouse button or the track pad a simple choice question popped up on the screen that asked the participant to choose whether the motion was made by a healthy or impaired person. After making their choice, a second multiple choice question about the movement pattern itself appeared. Participants had to choose between six motions: Jumping jacks, walking (WALK), sitting and standing (STS), climbing stairs (STAIRS), ice skating, and standing still. Experimental trials only included WALK, STAIRS and STS conditions. The remaining choices were distractors to determine if there were selection biases. The subjects had as much time as they needed to complete the two multiple choice questions. The subjects began the next trial by pushing a mouse button or track pad. From each trial we collected the latency to identify the motion, which covaried with the visual background noise, and the accuracy of the identification of the movement pattern and the health of the point-light display motion. Three catch trials provided data without the influence of visual background noise. To prevent selection anxiety, all participants were informed prior to the start of data collection that movement presentation was random and, as a result, that they may see some motions repeatedly and other motions not at all.
Statistics
The statistical evaluation was carried out with RStudio (Version 3.4, R Core Team (2017), R Foundation for Statistical Computing, Vienna, Austria). To analyze differences between the participants with HD and the participants without HD, we used the Fisher exact test to perform a 2x2 (group x motion or group x health) analysis for nominal data, or Wilcoxon rank-sum test for non-parametric data. Practice effects were analyzed using a linear model. Differences were considered significant at a level of p≤0.05.
RESULTS
Recognition of movement pattern
We compared the ability of participants with HD and non-HD peers to identify the movement pattern independent of the health of the person making the movement. Collapsed across all movements, participants with HD had significantly more difficulty correctly identifying the biological motion than control subjects (participants with HD 74.9%, participants without HD 87.5%, p < 0.001). Examining the movement types individually revealed that specific movements were responsible for this disparity between groups. There was no significant difference between participants with HD and controls in identifying walking (correct WALK: 89.6% of the HD group and 90.2% of the controls, p = 0.8916), but participants with HD performed significantly worse when identifying the STS and STAIRS movement patterns than participants without HD (STS: participants with HD 72.5%, controls 81.2%, p = 0.0106; STAIRS: participants with HD 62.8%, participants without HD 91.5%, p < 0.001) (Fig. 2).
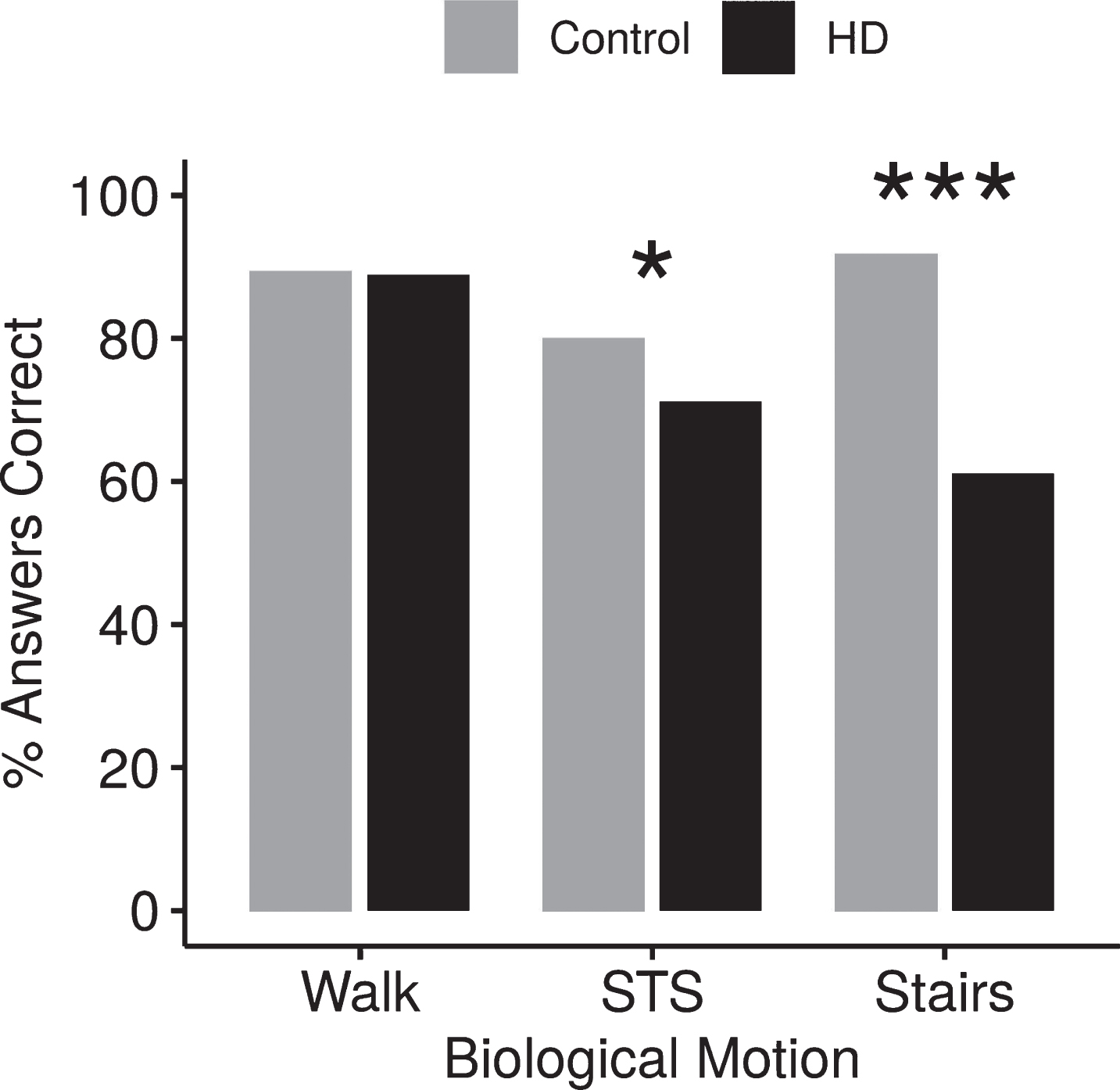
Effect of movement pattern on accuracy. Errors in detecting movement, collapsed across health, shows participants with HD perceive WALK but are significantly worse than their healthy peers in detecting STS and STAIRS. *p < 0.05, ***p < 0.001.
Latency and background noise
Participants without HD identified all three movement types more quickly than participants with HD (Table 2). The mean (±SD) time for detecting the WALK movement was 22.91 s (±6.1 s) for the participants with HD while the participants without HD identified the movement in 16.65 s (±7.12 s; p = 0.0032). Similarly, detecting the STS motion took 21.22 s (±6.26 s) for participants with HD and 16.53 s (±8.01 s; p = 0.0373) for the group without HD. The greatest difference occurred in identifying stair climbing. Whereas control participants detected the STAIRS motion in 12.61 s (±6.64 s), participants with HD required 20.68 s (±6.94 s), almost 67% more time than their peers. Thus, individuals without HD identified stair climbing more accurately and more quickly than other movements, whereas participants with HD identified and responded to the stair climbing movement at the same rate and speed as for the other movements.
Effect of movement pattern on time
Mean time (SD) for participants with HD and non-HD peers to determine biological motion observed. Participants with HD were always significantly slower in motion perception. *p≤0.05, **p≤0.005.
The difference in detection latency between participants with and without HD could reflect HD motor impairments in pushing the mouse button/track ball or more difficulty in perceiving the movement within the random background noise. The absence of background noise in the catch trials distinguished between these two possibilities. Because no significant difference existed in the latency in these catch trials (mean participants with HD = 7.54 s (±7.01 s), participants without HD = 7.93 s (±7.88 s), p = 0.9492), HD motor impairment did not contribute to the difference in detection latency. There were, however, significant differences between the groups in latency and accuracy of motion detection in all trials with background noise (mean latency: participants with HD = 21.62 s (±10.0 s), participants without HD = 15.27 s (±10.5 s), p < 0.001; accuracy: participants with HD 74.9%, participants without HD 87.5%, p < 0.001) as well as trials for each individual movement pattern. This difference reflected the ability of participants with and without HD to identify biological motion coherence in random visual noise. There were 13 coherently moving points in each trial. At the beginning of each trial, the coherently moving points were embedded in 400 randomly moving points, a motion coherence of 3.25%. As latency and the number of randomly moving background points were inversely correlated, participants without HD identified the coherent biological motion in 15.27 s, a coherence of 6.2%, whereas participants with HD required 21.62 s, a 10% coherence, to identify the biological motion.
Ability to distinguish between healthy and impaired movement
The ability of participants with and without HD to identify whether the point-light motion represented a healthy or impaired movement was independent of the movement pattern. Overall, participants without HD were better at correctly identifying a healthy motion than participants without HD (p≤0.01) but neither group was particularly skilled at identifying impaired motion (Fig. 3). When examining responses according to movement pattern, both participants with HD and participants without HD were only slightly better than chance at distinguishing whether STS or WALK motions were healthy or impaired (STS: participants with HD 53.8%, participants without HD 54.6%, p = 0.8722; WALK: HD 60.7%, control 56.5%, p = 0.3168). In contrast, participants without HD correctly identified whether the point-light person was healthy or impaired in 84.1% of the STAIRS trials while participants with HD only identified the correct health for STAIRS in 69.7% of the trials (p < 0.001).
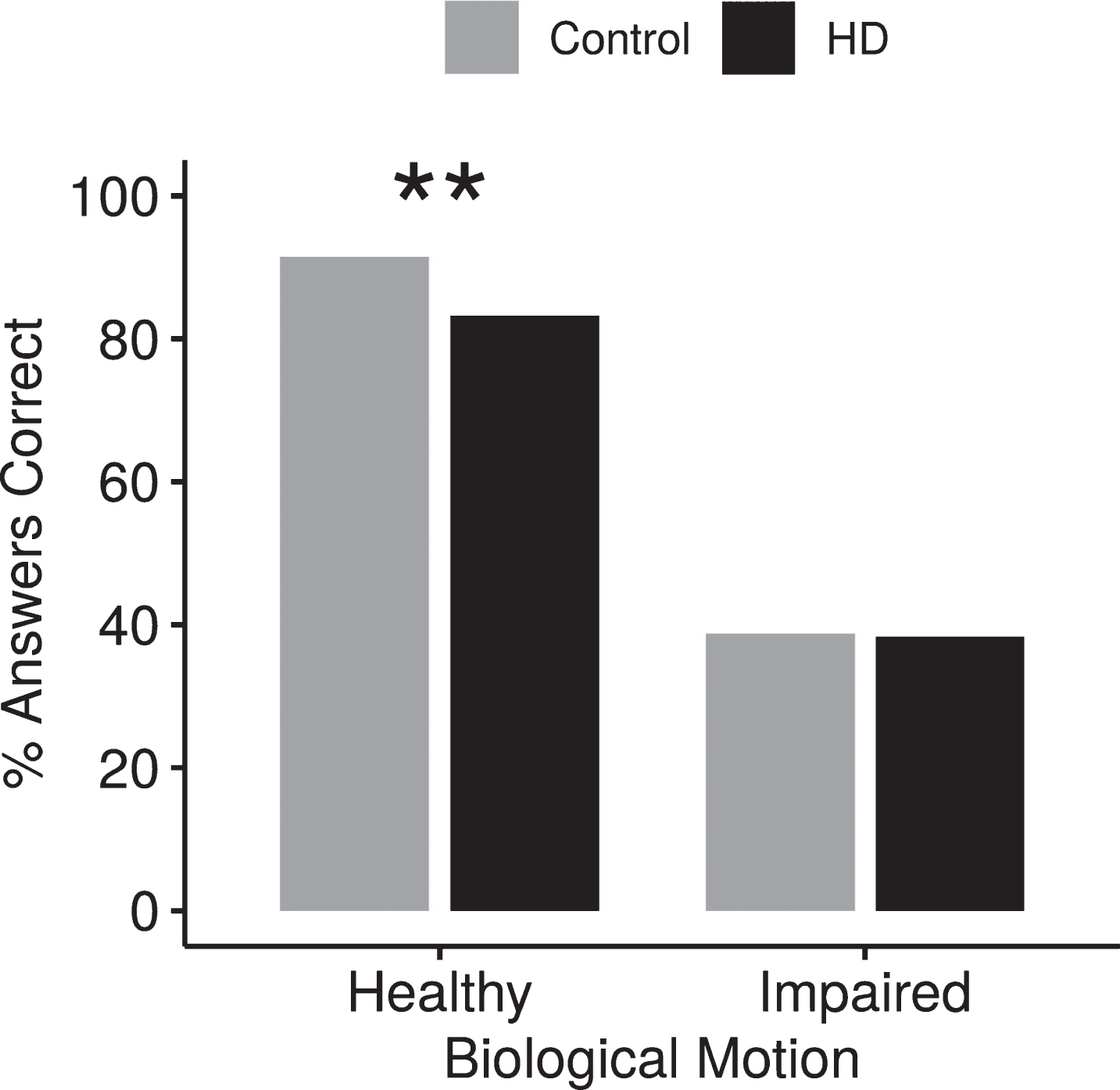
Effect of healthy or impaired motion on accuracy. Participants with HD were significantly worse at detecting healthy biological motion than non-HD peers. Both groups had difficulty identifying impaired biological motion. **p < 0.01.
Closer examination of the STAIRS data revealed that individuals without HD rarely misidentified the healthy point-light motion as impaired motion (7.4% incorrect responses), but misidentified the impaired motion as healthy on 24.4% of the trials. In contrast, HD subjects misidentified the healthy point-light motion as impaired on 16.8% of the trials and misidentified the impaired motion as healthy on 44.2% of the trials, a significantly higher error rate (p≤0.05).
Identifying both movement pattern and healthy vs. impaired motion
For participants with and without HD, correctly identifying both the movement pattern and healthy vs. impaired point-light motion was near chance levels when observing WALK and STS. However, the STAIRS movement revealed a significant difference between the groups (Fig. 4). Fifty-four percent of the participants with HD and 51.4% of those without HD could detect both the movement pattern and whether it was impaired or healthy when observing WALK (p = 0.4583) and 41.1% of the participants with HD and 45.1% of participants without HD correctly identified both movement pattern and whether the motion was healthy or impaired in STS trials (p = 0.3316). Accuracy of identifying both movement and health increased to 80.1% for the participants without HD in the STAIRS condition. In contrast, participants with HD identified both movement pattern and health type correctly in less than half the STAIRS trials (45.4%, p≤0.001).
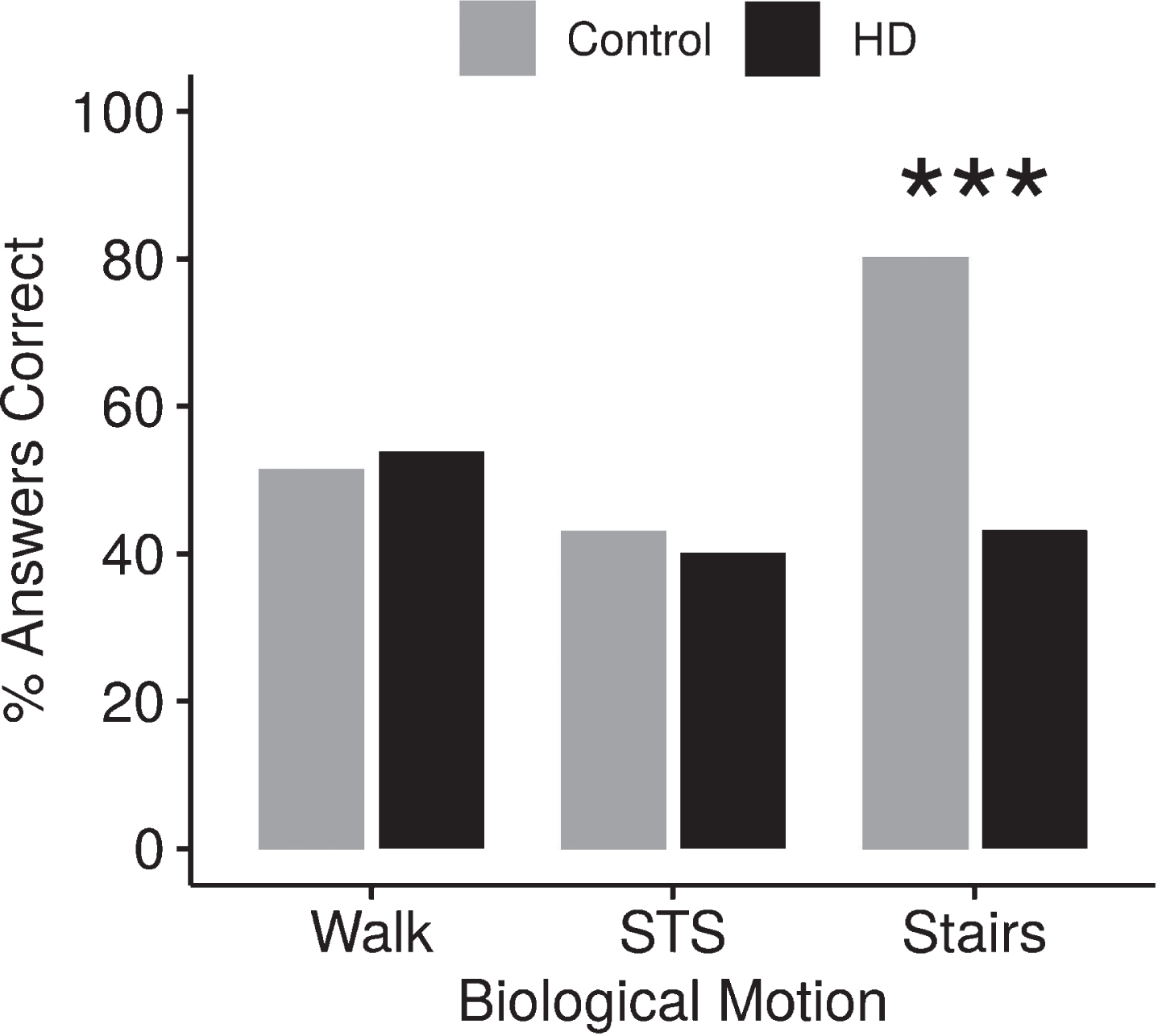
Effect of movement and health on accuracy. When asked to identify both the movement and whether the movement was impaired, both groups had similar accuracy during WALK and STS but a significant difference between the groups was seen for STAIRS. ***p < 0.001.
Practice effects
To determine if performance on this task was influenced by practice, we split the 42 test trials into three blocks (block 1 = trials 1–14, block 2 = 15–28 and block 3 = 29–42) and compared the results between groups when both movement pattern and motion health were answered correctly. Although the participants without HD always performed significantly better in accuracy and speed than the participants with HD (block 1 : 51.4% vs. 39.8%, p = 0.0052; block 2 : 58.2% vs. 49.1%, p = 0.0271; block 3 : 65.7 vs. 52.2, p < 0.001 for participants without HD and with HD, respectively), both groups improved across trials (Fig. 5). We applied a linear model to compute the rate of change from block 1 to block 3 and to predict accuracy and response latencies based on group and block interaction. Accuracy and speed were significantly improved from block 1 to block 3 for both groups (≤0.001). While the influence of practice on accuracy showed no significant difference between groups, participants without HD demonstrated a significantly greater rate of improvement in latency (i.e., decreased time to identify biological motion) than participants with HD (p ≤ 0.01).
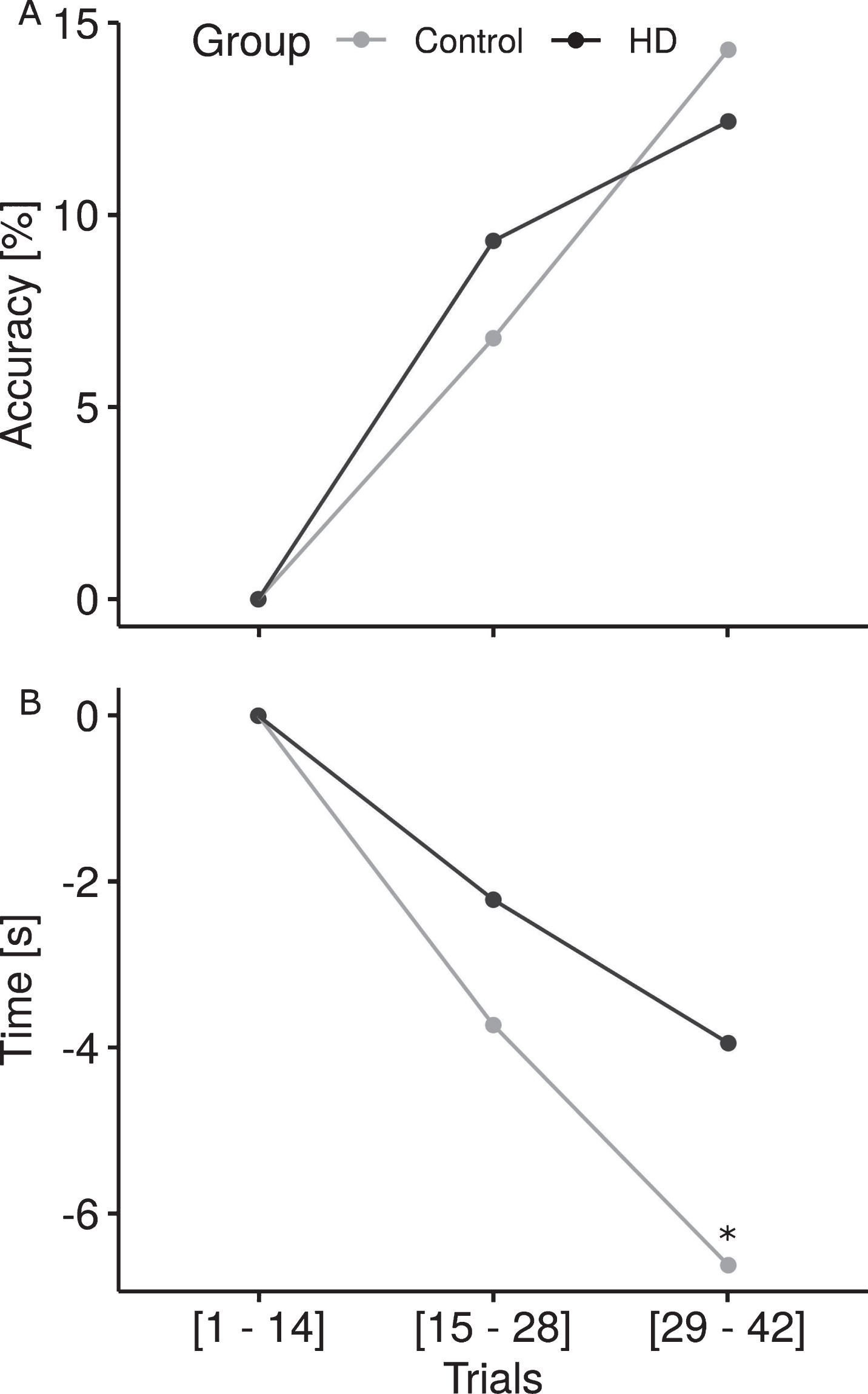
Practice effects on accuracy and latency. (A) Both groups improved accuracy over blocks (p≤0,001), however, participants with HD began to plateau while those without HD continued to learn at a steady rate. (B) Participants without HD demonstrated a greater practice effect on latency with a significantly greater rate of decline in the time taken to identify biological motion from block 1 to block 3 compared to those with HD (*p≤0.01).
Influence of viewing angle and stimulus inversion
To detect whether the viewing angle affected detection of movement pattern and health type, we compared front to side view when both movement and point-light image type were identified correctly. There was no significant difference between correct identification of front or side views although participants without HD were significantly better than those with HD for both viewing angles (correct identification front view: participants with HD 46.8%, participants without HD 60.8%, p < 0.001; correct identification side view: participants with HD 47.2%, participants without HD 56.0%, p = 0.00894). There was no significant difference between the participants with HD and participants without HD when the movements were shown inverted (correct identification of health and movement: participants with HD 33.3%, participants without HD 40.0%,p = 0.4671).
DISCUSSION
Movement pattern recognition
Participants with and without HD performed equally well in identifying walking biological motion even when embedded in noise. Significant differences appeared between the two groups, however, when identifying the sit-to-stand and stair climbing motions (Fig. 2). For participants with HD, performance dropped 17.1% for sit-to-stand and 26.8% for stair climbing biological motion relative to their accuracy in identifying the walking motion. In contrast, participants without HD demonstrated only a 9% decrease when observing sit-to-stand biological motion and actually improved by 1.3% with point-light images of stair climbing. The performance drop in correctly detecting the sit-to-stand motion by participants may result from the absence of feet and leg movement in this motion [24]. The impaired performance of participants with HD in correctly identifying sit-to-stand and stair climbing motion may also reflect individuals’ own difficulties with these motions. People with HD are slower than their healthy peers in performing sit-to-stand and exhibit significantly more sway [25]. Moreover, Grimbergen et al. (2008) reported a great number of falls in HD occurs during stair climbing [26]. The abnormal sensory feedback from movements created by the motor impairments of HD subjects coupled with frequent unawareness of their movement abnormalities [27] may distort their internal perception of normal movement. The ability to recognize coherently moving lights as a specific biological motion presumably occurs as an external movement is interpreted based on an internal model of the same movement, characteristic of mirror neurons. Consistent with this interpretation, fMRI studies of biological motion show activation of premotor cortex [21], the same region containing mirror neurons [28]. The decreased dynamic balance, mobility and motor performance that create an increased risk for falling in individuals with HD [29] reduces the sensory experiences of stair climbing and sit-to-stand movements and the abnormality of the movement distorts the internal perceptual model of the movement. This distortion may diminish the ability of the participant with HD to detect some types of biological motion. Because even people highly impaired by HD walk during their everyday life, biological motion perception may be less affected for walking than less practiced movements like stair climbing.
Latency
There are at least four possible explanations for the increased latency of participants with HD in detecting biological motion. Motor impairments could have slowed response latency in these participants compared to the non-HD peers. This explanation is unlikely, however, because the response latency of the two groups was not different on the catch trials. This result indicates that background noise affected those with HD more than participants without HD. The random background noise could distract the person with HD from following the coherently moving dots in three possible ways. One possibility is that the longer latency to identify biological motion results from impaired top-down attention processing. Using a simultaneous perception task, Finke et al. (2007) demonstrate that both participants with and without HD accurately identify a single item but that those with HD exhibit increased errors and processing time when there were multiple overlapping stimuli [30]. Detecting a movement covered by background noise depends on participants ability to ignore the internal and external noise, perceive the given stimulus [31] and compare it to existing templates of the particular motion [32]. A decline in top-down attention processing created by the background noise could increase reaction time in HD.
Another explanation is that HD oculomotor impairments decrease the ability of participants with HD to ignore the background noise. For example, the initiation of volitional saccades is delayed in people with HD and seemingly irrepressible reflexive saccades take the eyes away from fixation points [33]. Given that the Total Motor Score (see Table 1) of our participants shows noticeable eye movement abnormalities, detecting a movement embedded in randomly moving points may have required more time for HD than non-HD participants.
A third possibility is that HD impairs motion perception. Because latency and motion coherence changed linearly with time, participants with HD required a higher motion coherence to recognize the biological motion embedded in the random noise than did non-HD peers. Consistent with difficulties in motion perception, smooth pursuit eye movements are frequently impaired in HD [33, 34]. It seems likely that all three factors may have contributed to the prolonged latency of our participants with HD in identifying biological motion correctly.
Moreover, medication intake could have had a somnolent effect. While none of the participants appeared sedated in any way or complained of this side effect, we admit that medication intake is a limitation of our study and should be addressed in further investigations. Nevertheless, this explanation is unlikely, as participants with HD did not differ in their latency from participants without HD when no background noise was present.
Ability to distinguish between healthy and impaired movement
Participants with and without HD are equally adept at determining whether the individual making the WALK and STS motions in the point-light displays is healthy or impaired. In contrast, participants without HD correctly identified the health in 84.1% of the STAIRS trials while participants with HD were only correct on 69.7% of those trials. In addition, participants with HD identify the impaired biological motion as healthy (erroneous identification) on 44.2% of the trials, whereas participants without HD make that error on only 16.8% of the trials. These data indicate that our participants with HD could rarely distinguish impaired biological motion from healthy biological motion when the point-light image depicted stair climbing. The significant difference between the groups for the STAIRS motion may reflect the problems participants with HD experience in the execution of this motion. We propose that balance problems in HD lead to a decreased ability to climb stairs before preventing other elements of the motor repertoire [35–37]. The perception of human biological motion operates via a template matching approach [13, 14]. A person's own motor repertoire shapes these templates and therefore the impaired motor performance would determine evolving templates in HD [15]. If HD templates are changing as the disease progresses, such that ‘normal’ motion is increasingly supplanted by impaired, significantly different templates between the two participant groups would exist. Participants with HD may find it difficult to distinguish between healthy and impaired movement as a recognition between the previous ‘normal’ template and the new ‘impaired’ template would be required.
Identifying movement and health
When participants correctly identified whether the point-light image displayed a healthy or impaired movement, they usually identified the movement correctly as well. For participants without HD,identifying both the movement health and movement pattern correctly was only 6.3% less frequent than correctly identifying movement health alone. Participants with HD showed a response accuracy similar to non-HD participants for the WALK and STS movement patterns, identifying both movement health and movement pattern correctly only 9.4% less than just correctly identifying the movement health. In contrast, when viewing STAIRS, participants with HD were 24.3% less accurate identifying both the movement pattern and whether the motion was made by someone who was healthy or impaired compared to just correctly identifying the movement health. Our participants with HD only identified healthy or impaired point-light stair climbing on approximately two-thirds of the trials and correctly identified both on less than half of these trials.
Practice effects
Participants without HD demonstrated continued improvements throughout exposure to biological motion trials. While the participants with HD initially improved, they appeared to level off in the final block of trials with a slower rate of improvement overall. It may be that participants at HD approached a ceiling such that a threshold level of noise prevented biological motion perception. The coherence level required for those with HD was 10% compared to the approximately 6% threshold for those without HD. As coherence thresholds measured behaviorally have been shown to accurately reflect neuronal responses to coherence of visual stimuli [38], it is possible that neuronal damage occurring in HD limits the training response in the participants with HD. This may be an interesting avenue for further investigation related to the disease progression in HD.
Inversion effect
Inverting the biological motion image reduced the detection of health and movement pattern for both groups to chance levels. The difficulty in recognizing inverted point-light images is reflective of the assumption of gravity when interpreting biological motion. Impaired perception of inverted biological motion occurs in adults [39] and in infants as young as three months [40]. The importance of gravity for interpreting biological motion has also been shown in animals [41]. Such consistent findings between humans and animals suggests that beyond clinical testing, biological motion perception may be an appropriate measure in animal models of HD. Schuldenzucker et al. (2017) demonstrated the Libechov Minipig as a potential large animal model for HD with comparable size, weight and digestion. Introduction of a biological motion test in such existing animal models may complement the findings in this study and increase opportunities to understand the changes in biological motion perception as HD progresses.
Biological motion perception and other cognitive changes in HD
HD is known to effect cognition in addition to distinctive motor and behavioral symptoms. Cognitive changes include decreased information processing speeds, impaired working memory and learning, and poor emotional recognition [42]. The interaction of cognition and biological motion perception has shown that biological motion perception is related to social cognition but largely independent of other cognitive abilities [43]. We have shown that detecting biological motion is intact in HD but with increased latencies that may represent the impaired processing speed known to be part of the disease process. In addition, while perception of walking was generally unimpaired, perceiving more complex motions like stair-climbing was more difficult for participants with HD. In addition, the difficulty distinguishing coherent motion from random noise may reflect a previously demonstrated impairment of spatial working memory in HD [5]. The lack of specific cognitive outcome measures to correlate to the biological motion perception data is a major limitation of our study. Neuropsychological testing to determine potential deficits in attention, visual spatial working memory, and concentration would have allowed us to more fully explain differences between groups as these are known to be impaired in HD. The interaction of cognition with motion perception in HD is an area that deserves further attention in future work.
Conclusion
People with HD showed distinct deviations in detecting and interpreting biological motion compared to healthy peers. In addition, learning from repeated exposures to the biological motion images appeared to be decreased in HD. If mirror neurons in the premotor cortex that subserve both action and perception are disrupted, not only eye movements but whole body movement can be affected. In persons with HD this might result in choreatic movements and an impaired perception of biological motion even before other symptoms occur. These perception errors may serve as an early biomarker for disease onset and progression and to evaluate the success of novel therapeutics in HD.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
Footnotes
ACKNOWLEDGMENTS
CE and LM received funding for this project from the Thomas Hartman Center for Parkinson’s Research. This publication is part of the unpublished dissertation by Tamara Matheis.