
Abstract
Background:
Therapeutics that lower mutant huntingtin (mHTT) have shown promise in preclinical studies and are in clinical development for the treatment of Huntington disease (HD). Multiple assays have been developed that either quantify mHTT or total HTT but may not accurately measure levels of wild type HTT (wtHTT) in biological samples.
Objective:
To optimize a method that can be used to resolve, quantify and directly compare levels of full length wtHTT and mHTT in HD samples.
Methods:
We provide a detailed quantitative immunoblotting protocol to reproducibly resolve full length wtHTT and mHTT in multiple HD mouse and patient samples.
Results:
We show that this assay can be modified, depending on the sample, to resolve wtHTT and mHTT with a wide range of polyglutamine length differences (ΔQs 22–179). We also demonstrate that this method can be used to quantify allele-selective lowering of mHTT using an antisense oligonucleotide in HD patient-derived cells.
Conclusion:
This quantitative immunoblotting method can be used to reliably resolve full-length HTT alleles with ΔQs≥22 and allows for direct comparison of wtHTT and mHTT levels in HD samples.
INTRODUCTION
Huntington disease (HD) is caused by an autosomal dominant CAG repeat expansion in exon 1 of the huntingtin gene (HTT) resulting in an expanded polyglutamine (polyQ) tract in the huntingtin (HTT) protein [1]. CAG repeat lengths > 35 are pathogenic, with longer repeat lengths typically resulting in earlier age-of-onset and increased disease severity [2–4].
HTT is a conserved 348 kDa protein which acts as a scaffold to facilitate molecular interactions and functions in numerous cellular pathways [5–10]. The polyQ expansion in mutant HTT (mHTT) results in both the loss of normal, wild type (wt) HTT functions and toxic gain-of-functions leading to cellular dysfunction and neuronal death [11]. Preclinical studies have demonstrated that HTT lowering prevents HD-like behavioural and neuropathological phenotypes in HD rodent models [12–14].
HTT lowering can be achieved by inhibiting transcription, degrading the RNA transcript, or promoting clearance of the protein. Although HTT is essential for embryonic development [15] and is critical for survival of neurons in the brain [16, 17], short-term partial reduction of wtHTT is tolerated in both mice and non-human primates [18, 19]. However, it remains unknown how lowering of wtHTT will be tolerated long-term. One strategy to overcome this potential limitation has been to develop therapeutics selectively against mHTT by either targeting the CAG expansion directly [20–23] or by targeting polymorphisms in linkage disequilibrium with the mutant allele [12, 24–29]. Allele-selective lowering of mHTT has been achieved in preclinical studies using analogous modalities, including: RNA interference [19, 28], antisense oligonucleotides (ASOs) [12, 29], and synthetic zinc finger proteins [22]. Notably, multiple HTT lowering therapeutics are in clinical development for the treatment of HD.
The ability to reliably quantify both wt and mHTT in the same sample is necessary for evaluating the allele selectivity of candidate mHTT lowering therapeutics. Multiple antibody-based assays have been developed to detect mHTT and total HTT [30–37]. Assays used to detect soluble mHTT rely on antibodies raised against the expanded polyQ tract for specificity. Consequently, assays to specifically quantify wtHTT have been challenging to develop due to the lack of unique epitopes that distinguish it from mHTT.
Immunoblotting (or western blotting) has been used by our group and others to resolve full-length (FL) wt and mHTT in various HD animal models and human HD samples [12, 38–48]. However, reproducibly resolving FL HTT alleles by immunoblotting has proven to be challenging due to the large size of the HTT protein and the relatively small differences in polyQ lengths (ΔQ) between wtHTT and mHTT. Here, we provide a detailed immunoblotting protocol (Protocol 1) that utilizes gels with a low ratio of bis-acrylamide to acrylamide (1:200; low bis). This low bis ratio results in fewer crosslinks between acrylamide chains and thus creates larger pores in the gel which helps resolve small size differences in high molecular weight (MW) proteins. We demonstrate that this method is reproducible and can be modified to reliably resolve wtHTT and mHTT with a range of ΔQs in multiple HD mouse models and HD patient samples. We also show the utility of this assay in quantifying allele-selectivity of an ASO targeting mHTT.
MATERIALS AND METHODS
Animals
Experiments were performed using the HD mouse models: BACHD [49], YAC128 [50], Hu97/18 [44], Hu128/21 [45], and Q175FDN [51–53]. Animals were maintained under a 12 h light: 12 h dark cycle in a clean barrier facility and given free access to food and water. Experiments were performed with the approval of the Animal Care Committee at the University of British Columbia (A20–0107).
Cell lines
HD patient-derived lymphoblastoid cell lines GM03620, GM02150, and GM04724 were obtained from the Coriell NIGMS Human Genetic Cell Repository at the Coriell Institute for Medical Research. All cell lines were grown in RPMI 1640 media containing 10% fetal bovine serum and 1X Penicillin-Streptomycin (Table 1).
Reagents used for low bis immunoblotting method
HD patient brains
All samples were collected, stored, and accessed with informed consent and ethical approval from the UBC/Children’s and Women’s Health Centre of British Columbia Research Ethics Board (H06-70467, H06-70410).
Genotyping
Genomic DNA was extracted and purified using the DNeasy Blood & Tissue Kit (Table 1) and HTT CAG repeat sizing for HD mouse lines were performed by Laragen Inc. (Culver city, CA, USA). Sequenced CAG repeats are reported (Table 2). HTT CAG repeat sizing for human HD lymphoblast and brain samples was performed as previously described [29].
Gel running distance for the low bis immunoblotting method as determined by the ΔQ between wt and mHTT. MW, molecular weight
aSequenced CAG size bHuman HTT transgene CAG tracts contain CAA interruptions cTransgenic human HTTdMouse HtteChimeric mouse/human exon 1 inserted into mouse Htt.
Low bis immunoblotting
Immunoblotting was performed as described in Protocol 1 with the following specific details. For HD mouse brain blots, one cortex hemisphere from each line was lysed in 500 μL of SDP lysis buffer. For HD patient-derived lymphoblast blots, 5–10E+6 cells were lysed in 28 μL of SDP lysis buffer. For HD human brain blots, a small piece of frozen cerebellum tissue (approximately the size of a grain of rice) was lysed in 100 μL of SDP buffer. Fifty μg total protein from each sample was prepared for immunoblotting. HD mouse cortex lysates were run for ∼2 h, HD patient-derived lymphoblast lysates were run for ∼3.5 h and HD patient cerebellum lysates were run for ∼4 h, respectively. For HD mouse brain samples, membranes were incubated with anti-HTT antibodies: MAB2166 (mouse, 1:2000); HDB4E10 (mouse, 1:1000); MAB2168 (mouse, 1:500); EM48 (mouse, 1:200); BKP1 (mouse, 1:500); MW1 (mouse, 1:500); H7540 (rabbit, 1:1000); D7F7 (rabbit, 1:1000) and anti-calnexin (rabbit, 1:10000) as a loading control diluted in 5% BSA in PBST containing 0.01% NaN3 on a rocker overnight at 4°C (Table 1). For HD lymphoblast samples, membranes were incubated with anti-HTT antibodies: MAB2166 (mouse, 1:2000); HDB4E10 (mouse, 1:1000); H7540 (rabbit, 1:1000); D7F7 (rabbit, 1:1000) and anti-non-muscle myosin (rabbit, 1:2000) as a loading control diluted in 5% BSA in PBST with 0.01% NaN3 overnight on a rocker at 4°C. For HD patient brain samples, membranes were incubated with anti-HTT antibody MAB2166 (mouse, 1:2000) and anti-non-muscle myosin (rabbit, 1:2000) as a loading control diluted in 5% BSA in PBST with 0.01% NaN3 overnight on a rocker at 4°C. Following 3 × 10 min washes with PBST, membranes were incubated with IR dye 800CW goat anti-mouse (1:5000) and AlexaFluor 680 goat anti-rabbit (1:5000) labeled secondary antibodies diluted in 5% skim milk in PBST for 1 h at room temperature, protected from light (Table 1). Finally, membranes were washed for 4×5 min with PBST and imaged using the LI-COR Odyssey Infrared Imaging System (Li-Cor Biosciences).
Densitometry was performed on mHTT, Htt/wtHTT, calnexin and non-muscle myosin bands using the LiCor Image Studio Lite software and median signal intensity following background subtraction was used for analysis. For mouse brain blots, Htt/wtHTT and mHTT MAB2166 signal intensities were normalized to calnexin signal and data is presented both as relative Htt/wtHTT and mHTT to normalized BACHD signal for the same allele and as the ratio of normalized mHTT: Htt/wtHTT. For lymphoblast blots, wtHTT and mHTT MAB2166 signal intensities were normalized to non-muscle myosin signal and data is presented both as relative wt and mHTT normalized GM02150 signal for the same allele and as the ratio of normalized mHTT: wtHTT. N values in text and figure legends represent the number of biological replicates that were run and probed with the MAB2166 antibody. Additional blots were run to probe with other HTT antibodies. Symbols represent independent data points, bars represent the mean of all biological replicates and error bars represent the standard error from the mean.
3–8% Tris-acetate gradient gel immunoblotting
Fifty μg total protein of HD mouse cortex samples and HD lymphoblast samples (as described above) were loaded into NuPAGE™ 3–8%, Tris-Acetate gradient protein gels (Catalog #: EA03785BOX). Gels were run in Tris-acetate SDS Running Buffer (50 mM Tricine, 50 mM Tris Base, 0.1% SDS, pH 8.24) at 190V and transferred in NuPage transfer buffer under identical conditions to those described in Protocol 1.
ASO synthesis and preparation
A gapmer with a phosphorothioate-modified backbone and 4 LNA modifications in each wing targeting rs72239206 in human HTT was synthesized by Qiagen at a 200 nmol scale and purified by standard desalting. Lyophilized ASO was resuspended in Dulbecco’s phosphate-buffered saline (DPBS; no calcium or magnesium) and incubated at 37°C for 1 h. ASO concentrations were calculated based on: Concentration (M) = OD260 × dilution factor/ ɛ260 where ɛ260 = extinction coefficient of oligo at 260 nm.
Allele-selective lowering of mHTT
GM03620 lymphoblastoid cells were genotyped as previously described [29], and identified to be heterozygous for the A1 haplotype, in phase with the CAG expansion, with a C haplotype on the wt allele. GM03620 lymphoblasts were treated with ASO as in [25]. Briefly, the cells were seeded in a 12-well plate at a density of 1.25E+5 cells/well. The following day, a serial dilution was performed with ASO in growth medium and ASO was added to the cells to yield final concentrations from 312 nM to 5000 nM. Cells were incubated for 240 h prior to harvesting. Densitometry was performed as described above and FL wtHTT and mHTT MAB2166 signal intensities were normalized to the non-muscle myosin loading control and presented relative to untreated for each HTT allele. N values in text and figure legends represent the number of independent biological replicates performed. Statistical analysis was performed by two-way ANOVA with posthoc analysis (Sidak’s multiple comparison test) using GraphPad Prism version 8. Symbols represent the mean of all biological replicates and error bars represent the standard error from the mean. Alpha values of < 0.05 were considered significant for all analyses. The relative IC50 of the ASO against mHTT was calculated by fitting a non-linear curve with a variable slope to the mHTT data.
RESULTS
Using the low bis immunoblotting protocol, we were able resolve mHTT and mouse Htt or human wtHTT alleles in brain lysates from BACHD [49], YAC128 [50], Hu97/18 [44], Hu128/21 [45], and Q175FDN [51–53] mouse models of HD with ΔQs ranging from 77–179 (Fig. 1A; Supplementary Figure 1A-C; Table 2). Differences in FL mHTT polyQ length and ΔQs between HD mouse lines can be qualitatively observed. Relative levels of Htt/wtHTT and mHTT (Fig. 1B) and the ratios of mHTT: Htt/wtHTT for each line are presented to demonstrate the reproducibility of this method (N = 4 per line, Fig. 1C). Calnexin has previously been used as a loading control to normalize HTT levels for quantification in mouse samples by immunoblot [24, 27]. We found calnexin levels to be consistent between lines and reproducible between gels suggesting it can be used reliably as a loading control for large MW proteins in mouse samples. Additional mouse lysates run using the low bis method were probed with HTT antibodies raised against different epitopes that preferentially recognize either both mouse Htt and human HTT (Fig. 1D, E; Supplementary Figure 1D-F), human HTT (Fig. 1F, G; Supplementary Figure 1G) or mHTT (Fig. 1H; Supplementary Figure 1H). As expected, antibodies raised to distinct epitopes recognize FL HTT (or Htt) and potential cleavage fragments differently. We then compared the low bis method to another method commonly used in the literature for resolving HTT alleles, 3–8% Tris-acetate gradient gels [31, 46–48]. The same samples were run head-to-head using both methods under similar electrophoresis and identical transfer conditions (Fig. 1I; Supplementary Figure 1I-K). The 3–8% Tris-acetate gradient gels were able to resolve HTT from each line. However, the separation was not as clear or reproducible as with the low bis method.
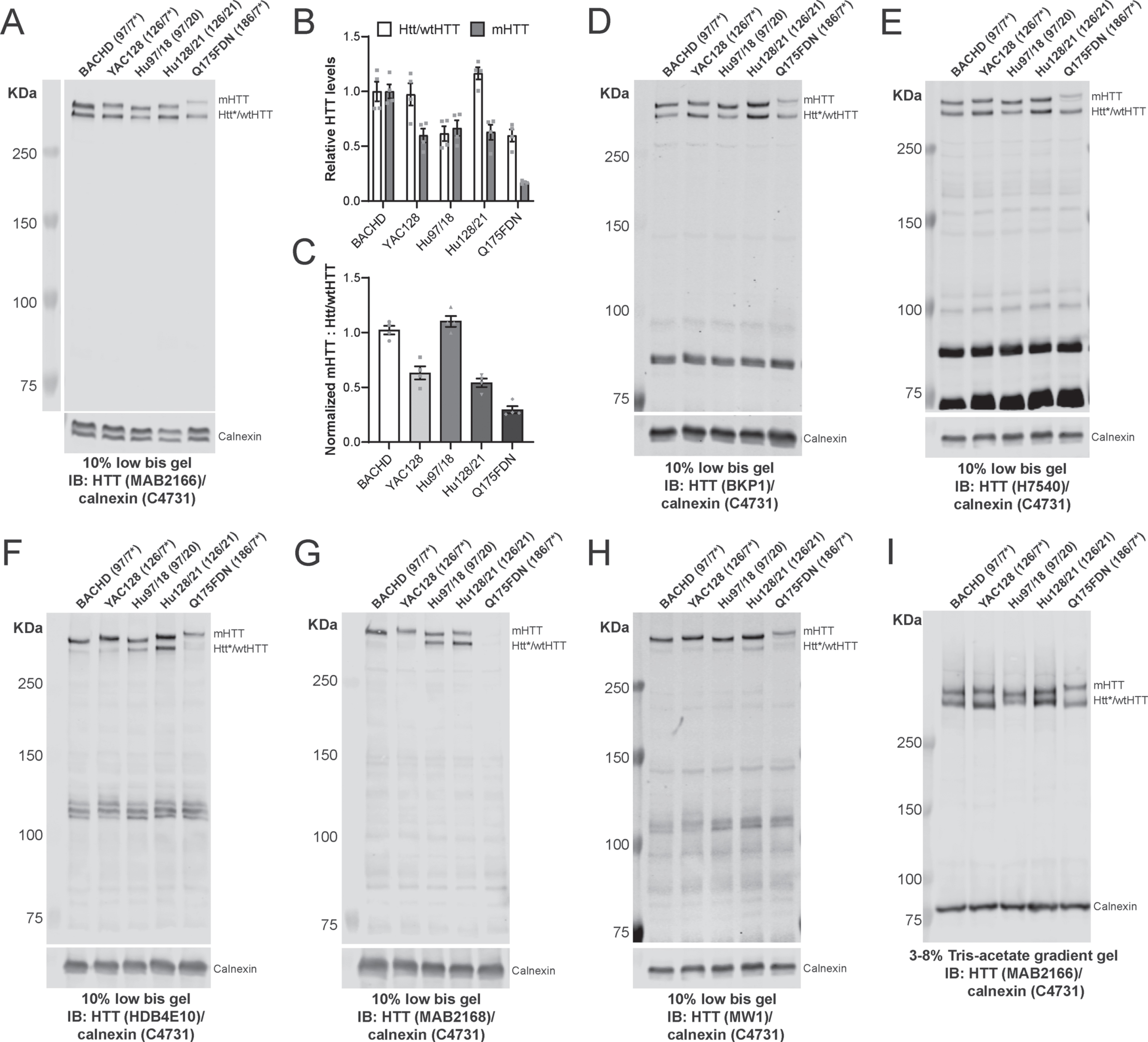
Low bis immunoblotting method resolves HTT alleles in HD mouse models. A) Cortical lysates from BACHD, YAC128, Hu97/18, Hu128/21 and Q175DN mice were run using the low bis protocol and probed with the MAB2166 HTT antibody. Densitometry was performed on MAB2166 signal and normalized to calnexin signal to quantify HTT levels. B) Relative levels of Htt/wtHTT and mHTT C) as well as the ratios of mHTT: Htt/wtHTT for each line are shown (N = 4 per line). Additional cortical lysates were run using the low bis method and were probed with D) BKP1, E) H7540, F) HDB4E10, G) MAB2168 and H) MW1 HTT antibodies. I) Cortical lysates from each line were also run on 3–8% Tris-acetate gradient gels under similar conditions to compare HTT resolution between methods. The number of CAG repeats of the HTT expanded and wild type (or mouse Htt) allele were determined by sequencing and listed next to the corresponding sample. *denotes mouse Htt which is present in the BACHD, YAC128 and Q175DN lines.
Most mouse models of HD harbour extremely long CAG repeat lengths rarely seen in individuals with HD. Therefore, we next wanted to evaluate whether the low bis method could be used to reliably resolve HTT alleles with ΔQs more representative of juvenile and adult-onset HD observed in the clinic. We ran lysates from three HD patient-derived lymphoblast lines, GM02150, GM03620, and GM04724 with ΔQs ranging from 27–52 using the low bis method (Fig. 2A; Supplementary Figure 2A-C; Table 2). Relative levels of wtHTT and mHTT (Fig. 2B) and the ratios of mHTT: wtHTT for each line are presented to demonstrate the reproducibility of this method (Fig. 2C). Non-muscle myosin has previously been used as a loading control for quantification of HTT in HD patient lymphoblasts by immunoblot [25, 29]. Additional lymphoblast lysates were run using the low bis method and probed with HTT antibodies raised against different epitopes to highlight that they recognize FL HTT and potential HTT cleavage fragments differently (Fig. 2D; Supplementary Figure 2D, E). We also tested the allelic separation of HTT in HD lymphoblasts using the 3–8% Tris-acetate gradient gel method. We found that 3–8% Tris-acetate gradient gels were able to resolve mHTT and wtHTT in GM04724 lymphoblasts but not in the GM03620 or GM02150 lines (Fig. 2E; Supplementary Figure 2F, G). In our hands, the low bis method was better suited for resolving HTT alleles with ΔQs<41 compared to the 3–8% Tris-acetate gradient gel method. Finally, to further evaluate the resolution limit of the low bis method, we ran cerebellar lysates from 3 HD patient brains (HD1-HD3) with a ΔQs ranging from 17–27 (Fig. 2F; Table 2). We were able to resolve FL wt and mHTT in both the HD1 and HD2 samples with ΔQs of 27 and 22, respectively. However, we were unable to clearly resolve a ΔQ of 17 in the HD3 sample which suggests that resolution limit of this assay may be a ΔQ≥22.
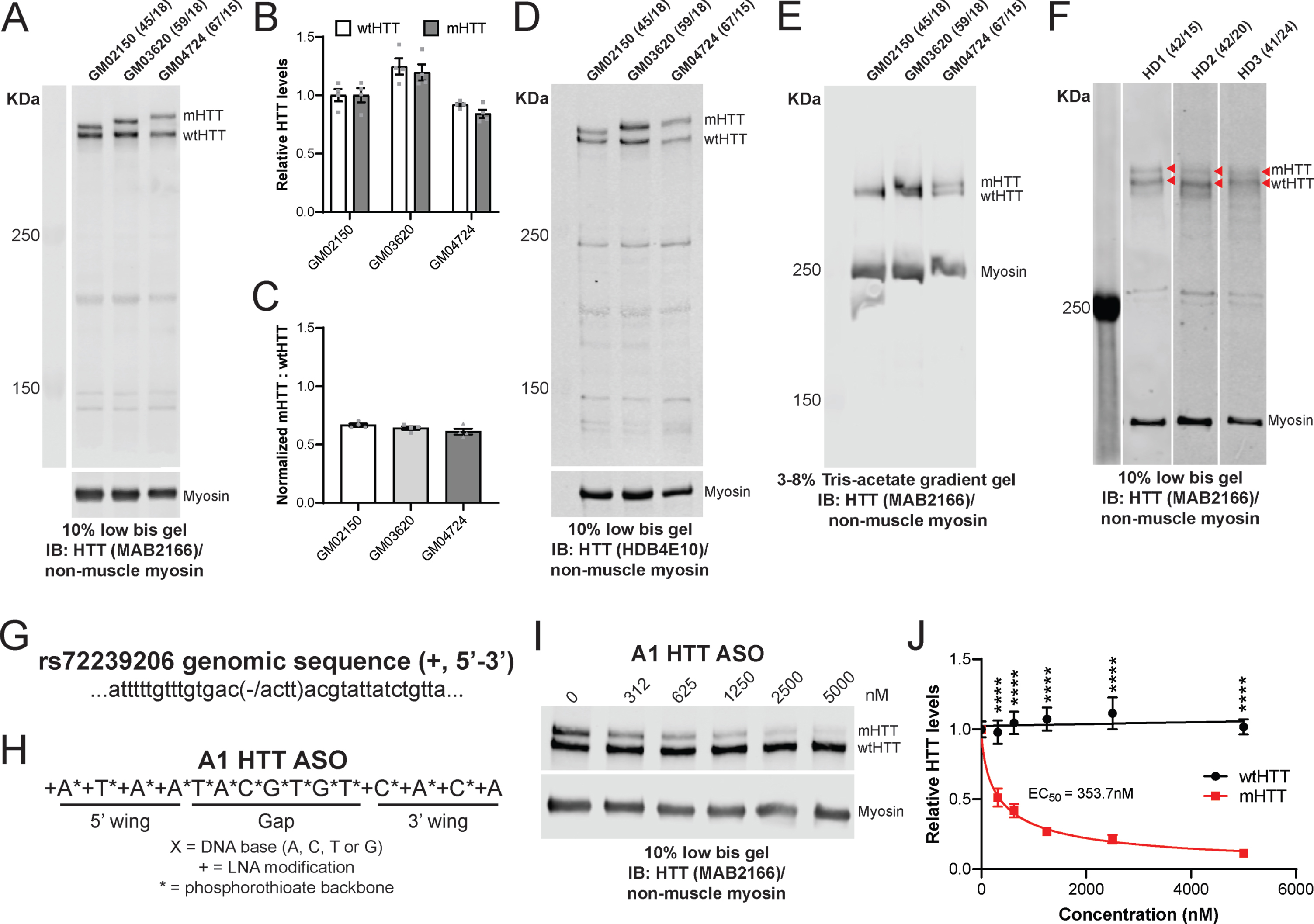
Low bis immunoblotting method resolves HTT alleles in HD patient samples and enables quantification of allele-selective lowering of mHTT. A) Lymphoblast lysates from GM02150, GM03620 and GM04724 lines were run using low bis gels and probed with the MAB2166 HTT antibody. Densitometry was performed on MAB2166 signal and normalized to calnexin signal to quantify HTT levels. B) Relative levels of wtHTT and mHTT C) as well as the ratios of mHTT: wtHTT for each line are shown (N = 4 per line). D) Additional lymphoblast lysates were run using the low bis method and probed with the HDB4E10 HTT antibody. E) Lymphoblast lysates from each line were also run on 3–8% Tris-acetate gradient gels under similar conditions to compare methods. F) Cerebellar lysates from three HD patients (HD1- HD3) were run using the low bis protocol and probed with MAB2166 to further evaluate the potential resolution limit of this assay. Red arrows indicate FL wtHTT and mHTT in each sample. The number of CAG repeats of the HTT expanded and wild type allele were determined by sequencing and listed next to the corresponding sample. Graphical representation of (G) the genomic sequence surrounding INDEL rs72239206 and (H) the sequence of the ASO gapmer targeting rs72239206 (A1 HTT ASO). Representative low bis immunoblot (I) and quantification (
To further demonstrate the utility of the low bis protocol, we tested whether it could be used to evaluate allele-selectivity of an ASO against mHTT. The HD patient-derived lymphoblast line GM03620 was identified as heterozygous at rs72239206 (Fig. 2G), a HTT polymorphism that defines the most common haplotype associated with HD, A1 [29]. We designed an ASO gapmer molecule complementary to the target transcript sequence (A1 HTT ASO, Fig. 2H) and treated GM03620 lymphoblasts with increasing doses of the A1 HTT ASO (312–5000 nM). Lymphoblasts were collected following a 240 h treatment and lysates were then run using the low bis protocol to resolve wt and mHTT (Fig. 2I; Supplementary Figure 2H-J). We observed that the A1 HTT ASO potently reduced mHTT in a dose-dependent manner (IC50 = 353.7 nM) while wtHTT levels remained relatively unchanged (Fig. 2J; N = 5 per dose; Two-way ANOVA dose p < 0.0001, HTT allele p < 0.0001, interaction p < 0.0001; Sidak’s multiple comparison test ****p < 0.0001). We found levels of non-muscle myosin to be reproducible between gels in both lymphoblast and brain samples and its levels were not altered in response to ASO-mediated HTT lowering. This suggests that non-muscle myosin can be used reliably as a loading control for high MW proteins in human samples.
DISCUSSION
Antibody-based assays used to detect HTT rely on different antibody combinations for the detection of mHTT and total HTT [30–36]. Current assays used to quantify soluble mHTT in biological samples rely on antibodies raised against the expanded polyQ tract for allele specificity. However, such antibodies (e.g., MW1) only preferentially detect mHTT through increased binding stoichiometry to longer polyQ tracts, and also bind to wtHTT polyQ tracts (Fig. 1H; Supplementary Figure 1H). This suggests that assays specific for mHTT may also quantify wtHTT, but to a lesser extent. Efforts to develop assays specific for wtHTT have been challenged by the lack of unique epitopes that distinguish it from mHTT. As a result, wtHTT concentrations are inferred from total HTT and mHTT levels in HD samples rather than directly measured [54–56]. Immuno-depletion of mHTT from samples using expanded polyQ antibodies could provide a means to enrich wtHTT concentrations for detection, however this approach would be limited by the same issues of antibody specificity against mHTT which could result in the inaccurate measurement of wtHTT levels. Using the low bis immunoblotting method, both alleles of FL, soluble, monomeric HTT can be reliably quantified on the same blot using the same antibody which allows for direct comparison of wtHTT and mHTT levels.
Despite the strengths of the low bis method, the assay is limited by its low throughput, the inability to resolve FL HTT alleles with small ΔQs (<22), and the relatively low sensitivity against HTT. More sensitive methods with limits of detection in the femtomolar range have been developed to quantify mHTT in the cerebrospinal fluid (CSF) of individuals with HD [32, 57] and to measure target engagement in HTT lowering clinical trials for HD [58]. Additional studies are required to further investigate what species of HTT are present in human CSF and whether these assays are measuring full length mHTT or smaller fragments of mHTT.
Using the low bis method, we show that we can reliably resolve FL wt and mHTT with ΔQs≥22 suggesting that this assay can be used to separate HTT alleles representative of adult-onset HD. We also demonstrate that we can reproducibly quantify allele-selective HTT lowering of mHTT in HD lymphoblasts.
Footnotes
ACKNOWLEDGMENTS
The authors would like to thank Mark Wang, Seunghyun Ko and Qingwen Xia for their technical support; Jeffrey Carroll, Niels Skotte and Amber Southwell for their contributions to the development of the low bis immunoblotting method. Project operational support for M.R.H. provided by a CIHR Foundation grant (FDN 154278).
CONFLICT OF INTEREST
The authors have no conflict of interest to report.