
Abstract
Background:
Synaptic changes occur early in patients with Huntington’s disease (HD) and in mouse models of HD. An analysis of synaptic changes in HD transgenic sheep (OVT73) is fitting since they have been shown to have some phenotypes. They also have larger brains, longer lifespan, and greater motor and cognitive capacities more aligned with humans, and can provide abundant biofluids for in vivo monitoring of therapeutic interventions.
Objective:
The objective of this study was to determine if there were differences between 5- and 10-year-old OVT73 and wild-type (WT) sheep in levels of synaptic proteins in brain and in neurofilament light chain (NfL) in cerebrospinal fluid (CSF) and plasma.
Methods:
Mutant huntingtin (mHTT) and other proteins were measured by western blot assay in synaptosomes prepared from caudate, motor, and piriform cortex in 5-year-old and caudate, putamen, motor; and piriform cortex in 10-year-old WT and OVT73 sheep. Levels of NfL, a biomarker for neuronal damage increased in many neurological disorders including HD, were examined in CSF and plasma samples from 10-year-old WT and OVT73 sheep using the Simoa NfL Advantage kit.
Results:
Western blot analysis showed mHTT protein expression in synaptosomes from OVT73 sheep was 23% of endogenous sheep HTT levels at both ages. Significant changes were detected in brain levels of PDE10A, SCN4B, DARPP32, calmodulin, SNAP25, PSD95, VGLUT 1, VAMP1, and Na+/K+-ATPase, which depended on age and brain region. There was no difference in NfL levels in CSF and plasma in OVT73 sheep compared to age-matched WT sheep.
Conclusions:
These results show that synaptic changes occur in brain of 5- and 10-year-old OVT73 sheep, but levels of NfL in biofluids are unaffected. Altogether, the data support a prodromal disease state in OVT73 sheep that involves the caudate, putamen and cortex.
Keywords
INTRODUCTION
Large animal models of disease can play an important role in pre-clinical research for therapeutics development as they possess similarities to humans when compared to rodents, including larger brains and a repertoire of motor behaviors and cognitive skills [1, 2]. In addition, they provide more biofluids which can be used to monitor disease progression in brain longitudinally. Large animal models of Huntington’s disease (HD) include those in pig, sheep, and non-human primate [1, 3–6]. A transgenic sheep (Ovis aries L.), named OVT73, was generated to express the full-length human mutant huntingtin (mHTT) (69 CAG+CAACAGCAACAG, or 73 glutamines) [7]. OVT73 HD sheep exhibit some characteristics of HD pathology as early as 6 months of age including reduced levels detected by immunoreactivity of GABA Aα1 receptor in the striatum and leu-enkephalin in the globus pallidus which originates from neostriatal neurons. Inclusions in cortical neuropil are seen at 18 months and in neuronal nuclei of piriform cortex at 3 years of age [8]. An altered metabolic profile and circadian abnormalities appear at 5 years [2, 10]. At 9 and 10 years of age, OVT73 HD sheep exhibit some neurological deficits, specifically in eye coordination and hind limb proprioception, which occur more frequently in females, and correlate with MRI findings in 10-year-olds that show evidence of white matter changes [11]. Altogether, these results indicate a disease process is underway within the first decade in this HD sheep model.
Loss of basal ganglia circuitry is thought to underlie subtle abnormal movements and cognitive changes that appear at early stages of HD [12]. There is support for synaptic dysfunction from studies in mouse models of HD, with changes in transcriptomes and proteomes of knock-in HD mice, as well as in electrophysiological recordings of medium spiny neurons in caudate putamen of transgenic mice expressing the full-length human transgene [13–16]. Some synapse-related transcripts and proteins that are changed in the neostriatum of HD mouse models include D1 and D2 receptors, DARPP32, SCN4B, and PDE10A [14, 17–22]. Synaptosomes are subcellular fractions enriched in presynaptic elements and some postsynaptic membranes. Western blot analysis of synaptosomes in HDQ140 mice showed significant changes in some synaptic proteins by 6 months of age, which coincide with behavioral deficits and the formation of mHTT aggregates in the caudate putamen in these models [15, 24].
Neurofilament light chain (NfL) is a subunit of the neurofilament family of proteins, which can polymerize to become intermediate filaments that extend from the neuronal cell body into axons. NfL detection in blood and cerebrospinal fluid (CSF) was identified as a biomarker for neuronal damage which is increased in patients with various neurological disorders including those with HD [25–27]. In a retrospective study of HD mutation carriers at early and late premanifest and manifest clinical stages 1 and 2, increased NfL levels in plasma correlated with decline in cognition and a rise in neuropathology assessed by MRI [25]. NfL detection in CSF is being used as a biomarker in clinical trials to assess treatment efficacy and safety and was found to be more predictive of progression in patients than mHTT levels in CSF [28].
In this study, we examined sheep synaptosomal fractions from caudate, motor cortex and piriform cortex of 5-year-old and caudate, putamen, motor cortex, and piriform cortex of 10-year-old WT and transgenic OVT73 HD sheep. We also analyzed biofluids (CSF and plasma) from 10-year-old sheep for NfL levels. Results showed significant changes in synaptic proteins in brains of OVT73 sheep compared to WT. However, there was no difference in NfL levels in plasma or CSF between the two groups, indicating that this biomarker may not be sensitive enough to detect early and subtle changes occurring in this HD model.
MATERIALS AND METHODS
Animals
Generation of the OVT73 sheep were reported previously [7]. The sheep express the full-length human huntingtin (HTT) cDNA bearing 69 CAGs+CAACAGCAACAG under human HTT promoter (1.1kb upstream of exon 1 sequence). The production of OVT73 transgenic sheep occurred at the South Australian Research and Development Institute (SARDI) in Turretfield Research Centre and was approved by the OGTR (NLRD 1037). The care of the OVT73 transgenic sheep was performed in accordance with the conditions approved by the Institutional Biosafety Committee (IBC) and Office of the Gene Technology Regulator (OGTR).
Collection of CSF and plasma
CSF and blood from ewes and rams were collected under anesthesia prior to euthanasia. CSF was drawn from lumbar puncture (preferred) or from the cisterna magna using a Spinal Tap Cannula (19 G x 9 cm). The CSF was promptly transferred into a pre-chilled conical bottom centrifuge tube and centrifuged at 2,000 g (10 min at 4°C) to remove cellular contamination. Supernatant was aliquoted and snap frozen in liquid nitrogen. Blood was collected from jugular venipuncture in lithium heparin tubes. The latter were gently inverted 5–8 times and were centrifuged at 2,000 g (10 min at 4°C). The resultant plasma was aliquoted and snap frozen in liquid nitrogen.
Brain tissue collection
5- and 10-year-old sheep were housed at SARDI and shipped to South Australian Health and Medical Research Institute (SAHMRI) 1 month prior to necropsy. The sheep were perfused with cold saline and brains were removed, immediately placed into cold saline and 8 mm slices were taken in coronal plane using a brain matrix. Each block was placed into a petri dish on top of a stainless-steel tray with dry ice underneath and cut at midline. The right and left hemispheres were cut coronally into 4 mm blocks. The right hemispheres were snap frozen and left hemispheres were fixed in 10% formalin. Frozen pieces of caudate, motor cortex, and piriform cortex from 5-year-old sheep (n = 12 WT and 13 OVT73, all rams) were dissected and sent to the DiFiglia laboratory at MGH where they were stored at –80°C until use. Pieces of caudate, putamen, motor cortex, and piriform cortex ( 2 mm3) from 10-year-old sheep (n = 10 WT ewes, 5 WT rams, 10 OVT73 ewes, 5 OVT73 rams) were dissected from frozen blocks either at UMass Chan Medical School or from blocks sent to MGH. It should be noted that the caudate putamen is one structure in mouse and separate in sheep.
Preparation of synaptosome fractions
We used protocols reported previously for preparing synaptosomes from mouse brain [15, 23]. Brain tissues from caudate nucleus, putamen, motor cortex, and piriform cortex were homogenized on wet ice using a Dounce homogenizer (tight piston B, 8 strokes per sample) in 7 ml 0.32 M sucrose containing 10 mM DTT and protease inhibitors (Roche, #11836170001). After a centrifugation at 1000xg for 10 min at 4°C, the supernatant (post-nuclear fraction) was layered on top of an equivalent volume of sucrose (1.2 M) and further centrifuged at 160,000xg with a SW41 swinging bucket rotor in a Beckman L8-80M ultracentrifuge for 35 min at 4°C with low acceleration and no brakes. The synaptosome fraction corresponds to the visible white layer between the two sucrose fractions. Synaptosome enrichment was verified by western blot analysis of presynaptic markers SNAP 25 and the postsynaptic marker PSD95.
Western blotting
A final panel of 15 antibodies was selected for screening that have been reported to be changed in synaptosomes of HD mouse models and can detect the endogenous sheep proteins [15, 23]. Some of the proteins (PDE10A and SCN4B) were selected based on proteomics results in mice [15]. In some cases, additional proteins were tested (GFAP, alpha-actinin1, or synaptophysin). The protein concentration of synaptosome fractions was determined using the Bradford method (BioRad). 5μg of protein from each sample were run on Criterion XT 4–12% Bis-Tris gels (BioRad, #3450125). To separate WT and mutant HTT, a Criterion 3–8% Tris-acetate gel (BioRad, #3540131) was used and run until the 95kD marker was at the bottom of the gel. Proteins were transferred onto a nitrocellulose membrane using TransBlot Turbo apparatus (BioRad #170-4150). Membranes were cut horizontally to optimize the number of proteins analyzed on each blot. Membranes were blocked in 5% milk (BioRad, #170-6404) in TBST at room temperature for 1 h then incubated in primary antibodies diluted in blocking buffer overnight at 4°C. After washing in TBST, blots were incubated in secondary antibodies coupled with peroxidase (JacksonImmunoResearch, #711035152, #715035150, #713035147) for 1 h at room temperature. An additional washing step was performed before incubating membranes in SuperSignal West Pico Plus Chemiluminescent substrate (Pierce #34580) for 5 min at room temperature. Images were acquired with a CCD camera (AlphaInnotech) and exposure to Hyperfilm ECL (GE Healthcare #28906839). For re-probing, blots were washed in TBST, stripped with Restore Western Blot Stripping Buffer (ThermoScientific, 21059), washed again, then incubated with primary antibodies.
Antibodies
The following antibodies were used for western blotting: PDE10A (1 : 2000, Abcam #ab177933), GABA1 beta1 (1 : 500, Abcam #ab16703), DARPP32 (1 : 8000, Abcam #ab40801), VAMP1 (1 : 1000, Abcam #ab3346), PSD95 (1 : 1000, Cell Signaling #2507), VGLUT1 (1 : 10,000, Synaptic Systems #135302), SNAP25 (1 : 5000, BD Transduction Laboratories #610366), Calmodulin (1 : 2000, Abcam #ab45689), Na+/K+ ATPase (1 : 10,000, Affinity Bioreagents #MA3-915), VGLUT2 (1 : 2000, Synaptic Systems #135402), Syntaxin1 (1 : 8000, Millipore #AB5820), Synaptophysin (0.25 mg/ml, Boehinger Mannheim # 902314), Pan-Trk (1 : 2000, Abcam #ab76291), GFAP (1 : 8000, Millipore #AB5804), SCN4B (1 : 1000, Abcam #ab80539), VAMP2 (1 : 2000, Abcam #ab18014), HTT (1 : 2000, Ab1, aa1-17 [29]; 1 : 500, MAB2166 aa181-810, (EMD Millipore); PolyQ 3B5H10 (1 : 1000, Sigma #P1874), Vinculin (1 : 2000, Sigma #V9131), GAPDH (1 : 5000, Millipore #MAB374), Alpha-actinin1 (1 : 500, Abcam #10861), Alpha-actinin2 (1 : 500, Sigma #A7732), Actin (1 : 500, Sigma #A4700).
Mutant HTT detection in CSF
CSF samples from WT (n = 21 0.6-, 10 7- and 16 8-year-old) and OVT73 (n = 20 0.6-, 7 7- and 15 8-year-old) sheep were analyzed by Evotec (Hamburg, Germany) and were not available for NfL analysis. The samples were prediluted in aCSF+1% Tween-20 + PI with a dilution factor of 1 : 2.5 and tested in triplicate by Singulex Erenna detection assay using 2B7 and MW1 antibodies based on the method described by Wild et al. [30]. Statistical analysis was performed using a non-parametric Kruskal-Wallis with Dunn’s multiple comparisons test where appropriate.
Detection of NfL
NfL levels were assessed in CSF and plasma from 10-year-old sheep using the Simoa NF-light Advantage kit (Quanterix, Item 103186) and run on the HD-X Analyzer (Quanterix) in the MADRC/MIND Biomarker Core. Assays were performed according to manufacturer’s specifications. Samples of CSF or plasma were diluted for the assay either by the machine (dilutions lower than 1 : 12) or manually (dilutions higher than 1 : 12). Every sample was run in technical duplicate. Intra-assay reliability was assessed by calculating the coefficient of variation (<20%). Values below the lower limit of quantification and values detected as outliers by the ROUT method (Q = 1%), were excluded from the analysis. Results were analyzed with GraphPad Prism 9 software.
RESULTS
Detection of HTT and other proteins in synaptosomes of caudate, putamen, motor cortex and piriform cortex of WT and OVT73 sheep
The levels of HTT and other proteins were measured in synaptosomes from different brain regions of 5- and 10-year-old WT and OVT73 sheep. The quality of the subcellular fraction for synaptosomes was confirmed based on the enrichment of the presynaptic and postsynaptic proteins SNAP25 and PSD95, respectively compared to post-nuclear fractions (Supplementary Figure 1). A panel of antibodies was used that detected proteins known to be affected in HD patient brain or changed in synaptosomes of HD mice based on western blot and proteomics (Figs. 1 2). The presence of mHTT in synaptosomes of the OVT73 sheep was achieved using an antibody with a polyglutamine(Q)-dependent affinity (3B5H10) [31] or with MAB2166 that does not detect endogenous sheep HTT [8]. Antibody Ab1 which recognizes HTT 1-17aa, was used to probe for WT and mHTT, and revealed low levels of human mHTT protein in synaptosomes compared to endogenous sheep HTT (21.6–23.6% of sheep HTT, Fig. 3). The gel types used in Figs. 1 2 do not separate the transgenic mHTT from the endogenous WT HTT so in these figures, antibody Ab1 detects both as total HTT. The levels of mHTT remained constant with increase in age. In addition, analysis of mHTT levels in CSF by an ELISA-based assay (Singulex assay) indicated no difference between samples from 0.6-, 7-, and 8-year-old sheep (Fig. 3D).
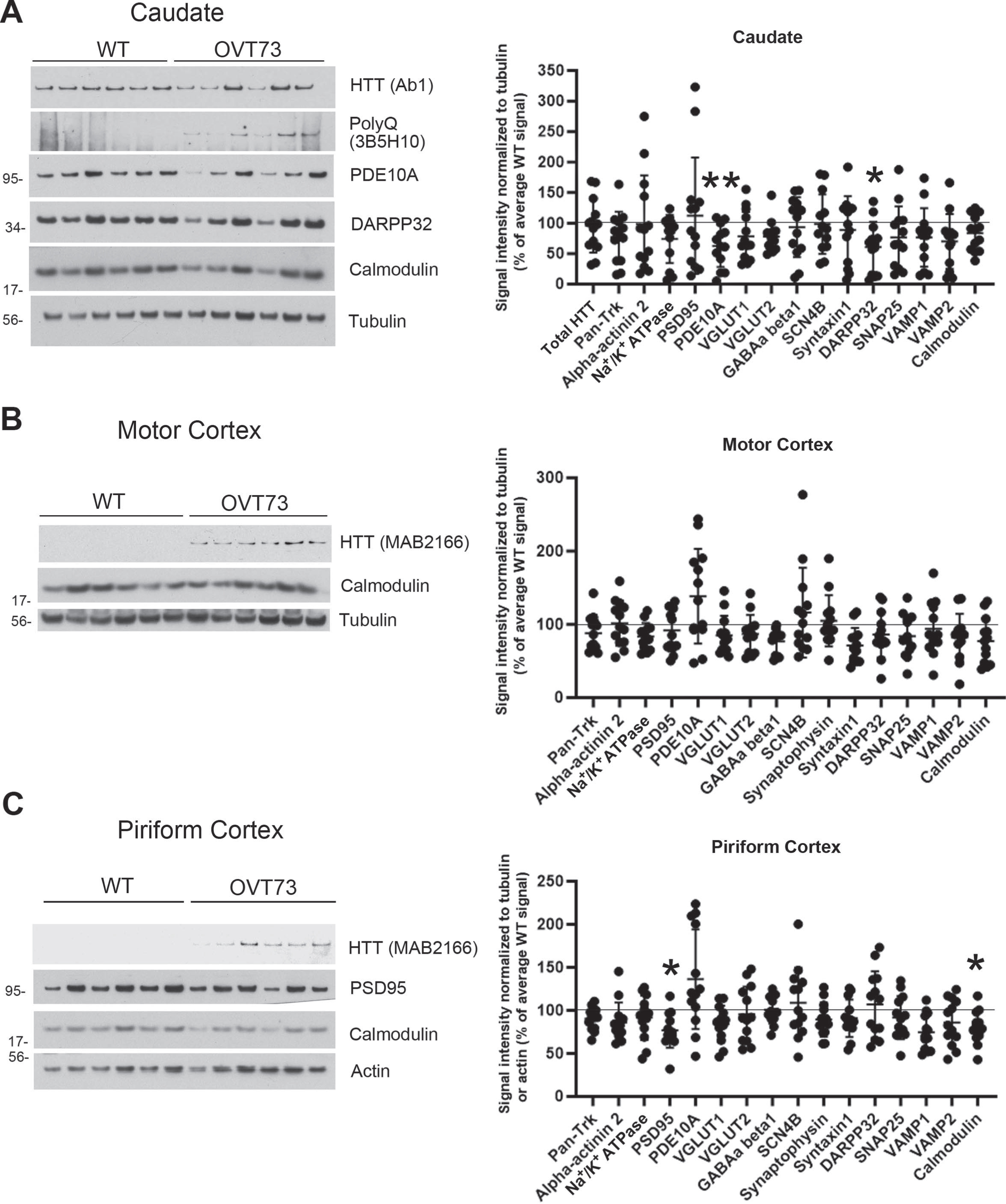
Protein changes in synaptosomes of 5-year-old OVT73 sheep. Representative western blot images of synaptic proteins that were found changed on 2 separate gels are shown for caudate (A) and piriform cortex (C). Total HTT or mHTT and calmodulin western blot images are shown for all brain regions (A, B, C). Significant reductions in levels of PDE10A (–37%) and DARPP32 (–37%) were found in the caudate and PSD95 (–23%) and calmodulin (–20%) in the piriform cortex in OVT73 sheep compared to WT. No significant changes were found in the motor cortex. Graphs (A, B, C) show mean total pixel intensity±SD for each protein normalized to tubulin or actin loading controls as a percent of the average signal obtained in the WT sheep. Statistics are unpaired t tests with N = 12 WT and 13 OVT73 sheep (*p < 0.05, **p < 0.01 compared to WT sheep). Molecular mass markers are shown to the left of the western blots. HTT migrates at around 350kD. Western blots of all samples are shown in Supplementary Figure 2.
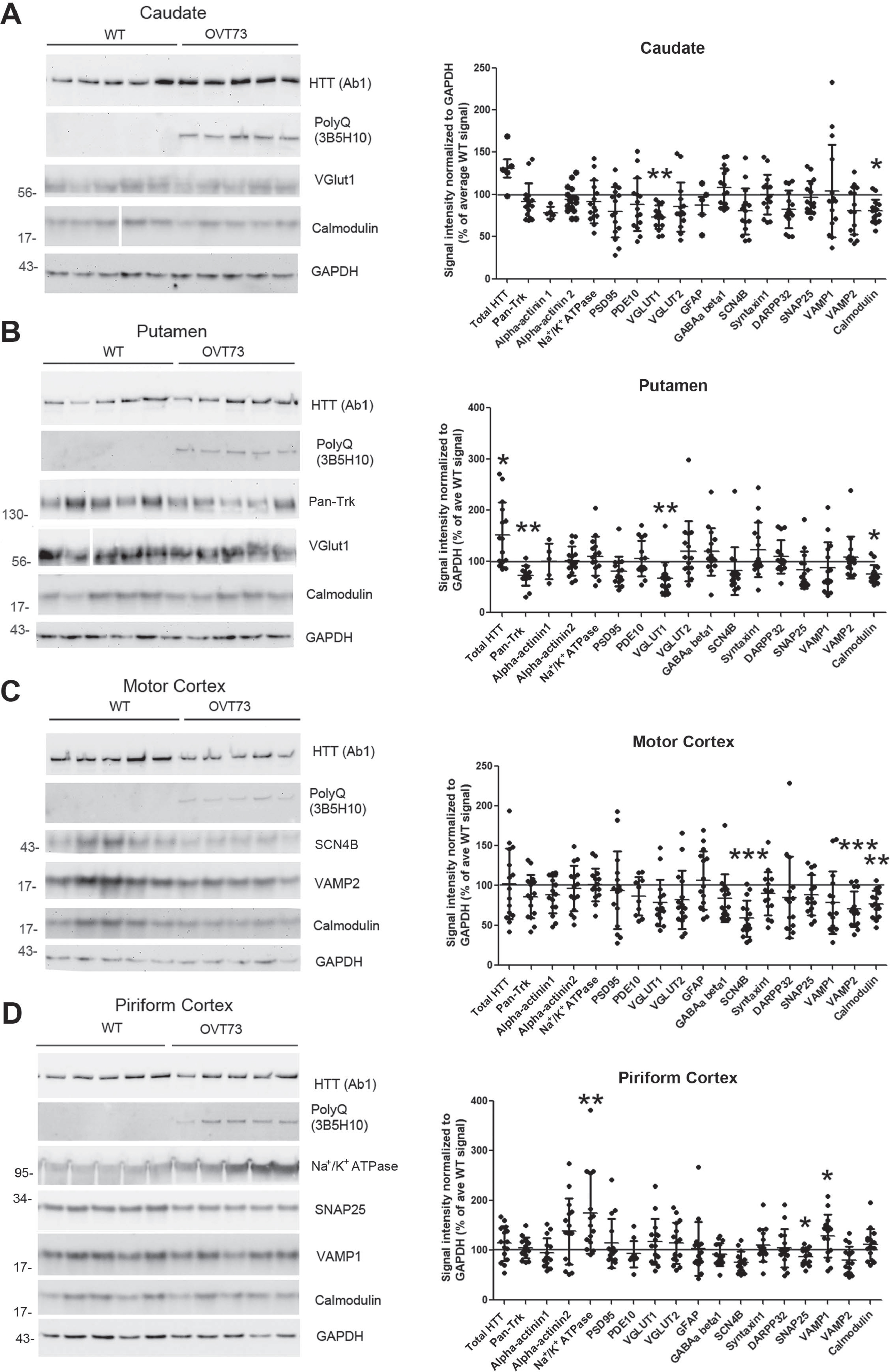
Protein changes in synaptosomes from caudate, putamen, motor cortex and piriform cortex of 10-year-old WT and OVT73 sheep. Representative western blot images detecting synaptic proteins that were found changed in the caudate (A), putamen (B), motor cortex (C), and piriform cortex (D) and their corresponding quantification. Total HTT, mHTT and calmodulin western blot images are shown for all brain regions (A, B, C, D). A) In the caudate, calmodulin protein levels were reduced by 20% and VGLUT1 by 27%. B) In the putamen, calmodulin levels decreased by 24%, VGLUT1 by 34%, and Pan-Trk by 28%, while HTT levels were increased by 52%. C) In the motor cortex, calmodulin levels were reduced by 23%, SCN4B by 41% and VAMP2 by 30%. D) In the piriform cortex, SNAP25 levels were reduced by 13% and Na+/K+ ATPase and VAMP1 levels were increased by 74% and 29% respectively. Graphs show mean signal intensity±SD measured in ImageJ and normalized to GAPDH. Each dot represents one animal. For most of the protein tested, intensity was assessed from 14 WT samples and 15 OVT73, except in caudate (A), HTT, GFAP and alpha-actinin1: n = 7 for WT and n = 5 for OVT73; in putamen (B), GFAP and alpha-actinin1: n = 7 for WT and n = 5 for OVT73; in motor cortex (C), PDE10A: n = 8 for WT and n = 10 for OVT73. All statistics are by default unpaired t-test between WT and OVT73 for each protein. Welch’s correction was applied if variance between WT and OVT73 was not equal; non-parametric test was applied if Gaussian distribution was not verified. (*p < 0.05, **p < 0.01, ***p < 0.001). Values are reported as percentage of WT for graph convenience. Spaces in a and b indicate removal of lanes. Molecular mass markers are shown to the left of the western blots. HTT migrates at around 350kD. Western blots of all samples are shown in Supplementary Figure 3.
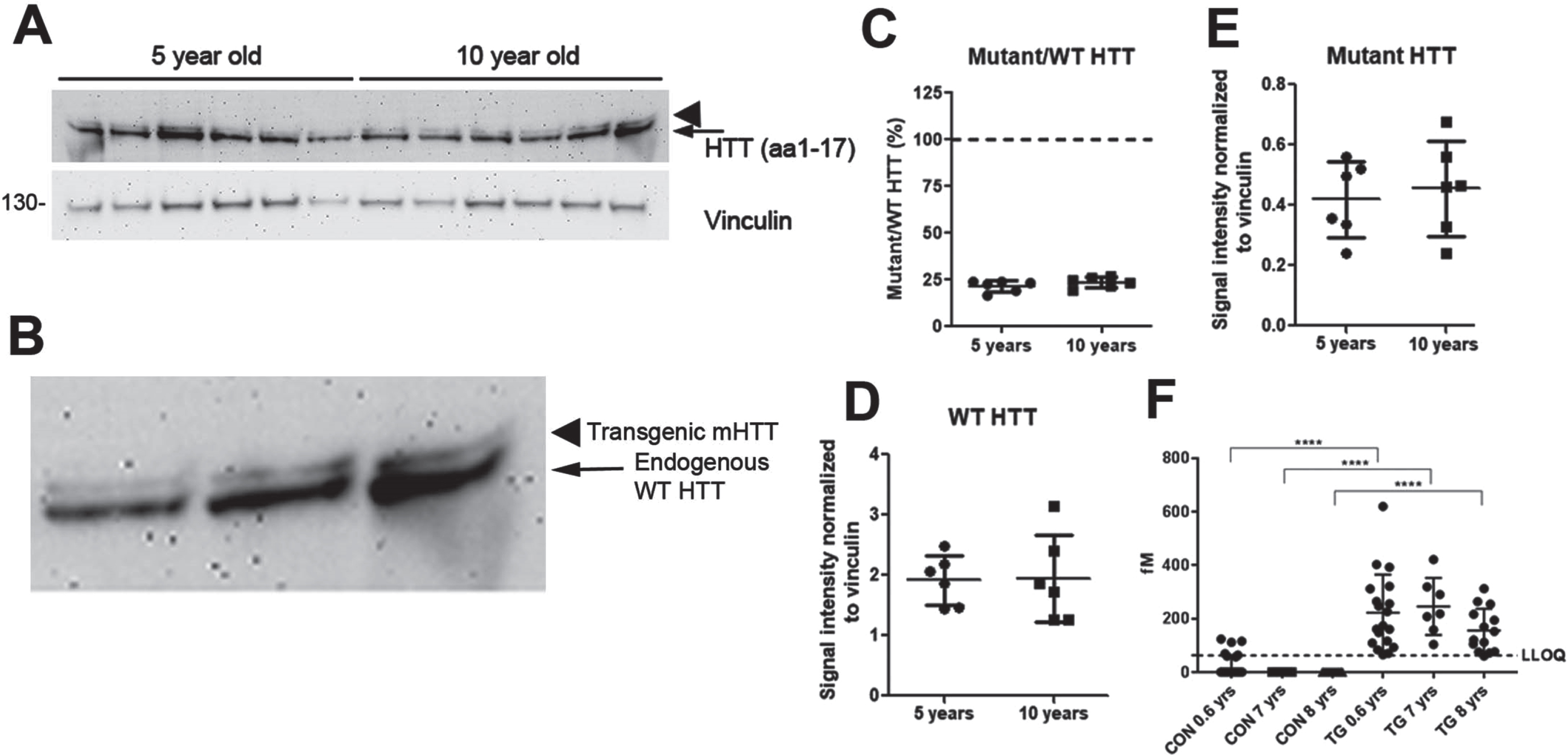
SDS soluble transgenic human mHTT is low compared to endogenous sheep HTT. A) Western blot assay of synaptosomes from caudate of some of the 5 and 10-year-old sheep probed with anti-HTT Ab1 (aa1-17) antibody with an enlargement of the last 3 lanes shown in B. Molecular mass markers are shown to the left of the western blots. HTT migrates at around 350 kD. C) Low levels of human mHTT are detected relative to endogenous WT HTT in 5- and 10-year-old OVT73 HD sheep (21.6% and 23.6% respectively). D, E) There is no significant difference in the levels of endogenous WT HTT (arrow in A and B) or mHTT (arrowhead in A and B) between 5- and 10-year-old OVT73 sheep (C, D, E, mean±SD, unpaired t-test, n = 6 5- and 10-year-old sheep). F) Expanded mHTT levels in groups of control sheep (CON) and transgenic OVT73 sheep (TG) at different ages (0.6 years; 7 years; 8 years). Data are shown as median±interquartile range. Statistical analysis was performed using a non-parametric K Kruskal-Wallis with Dunn’s multiple comparisons (****p < 0.0001, n = 21 0.6-, 10 7-, and 16 8-year-old WT sheep and n = 20 0.6-, 7 7-, and 15 8-year-old OVT73 sheep). For control groups, HTT levels were set arbitrary to 0.00001 for presentation. CON, Control (WT) sheep; TG, Transgenic (OVT73) sheep.
There were changes in levels of 11 proteins in the OVT73 sheep brain, but there was no consistent pattern by age or brain region. For instance, synaptosomes from 5-year-old sheep had lower levels of PDE10A and DARPP32 in the caudate; while PSD95 and calmodulin levels were decreased in the piriform cortex (Fig. 1). At 10 years of age, OVT73 HD sheep caudate had lower levels of VGLUT1; putamen had altered levels of Pan-Trk and VGLUT1; in the piriform cortex Na+/K+ ATPase, SNAP25 and VAMP1 were changed, and in motor cortex there were differences in SCN4B and VAMP2 (Fig. 2). There was an increase in levels of total HTT in the putamen of the 10-year-old OVT73 sheep which is largely due to an increase in endogenous sheep HTT (see Fig. 3). Of interest, calmodulin was reduced in the caudate, putamen, and motor cortex of OVT73 HD sheep. The percent changes in these proteins in OVT73 sheep compared to WT are summarized in Table 1. To determine if there was a sex effect in protein levels between 10-year-old OVT73 and WT sheep, the data for ewes and rams were separated (Supplementary Table 1). In the caudate and putamen, there were no protein changes detected in the rams but several in the ewes. In the motor cortex, there were more protein changes in rams than in the ewes. The proteins changed in OVT73 HD sheep partially overlapped with those affected in HD knock-in mouse models previously investigated (Supplementary Table 2). More proteins were changed in previous analysis of caudate putamen synaptosomes of 12-month-old HDQ140/Q140 mice than in 10-year-old OVT73 HD sheep brain. It is important to note that the analysis of proteins changed in OVT73 HD sheep brain synaptosomes was limited to antibodies that detect sheep.
Summary of protein changes in synaptosome fractions of 5- and 10-year-old OVT73 sheep (Unpaired t-test)
NS, not significant.
NfL measurements in CSF and plasma of WT and OVT73 sheep
Elements present in a tissue or biofluid can interfere with the accurate evaluation of an analyte of interest [32, 33]. This phenomenon, known as the matrix effect, may result in an under- or over-estimation of the analyte concentration. To assess the matrix effect in CSF and plasma, NfL was measured at different dilutions of these biofluids to determine the minimum required dilution (MRD). A difference higher than 20% indicates a matrix effect and is reported in Tables 2 3. The analysis revealed that the MRD for CSF is 100x (Table 2), and for plasma is 16x (Table 3) for samples from 10-year-old sheep.
Determination of minimal required dilution (MRD) to measure NfL levels in 10-year-old sheep CSF and avoid the matrix effect using the Simoa NF-light Advantage kit
Determination of minimal required dilution (MRD) to measure NfL levels in 10-year-old sheep plasma and avoid the matrix effect using the Simoa NF-light Advantage kit
Using the MRDs obtained above, CSF and plasma from WT and OVT73 sheep were subjected to NfL detection using the Simoa assay. As indicated in Fig. 4, the results indicate no significant difference in NfL levels between WT and OVT73 HD sheep.
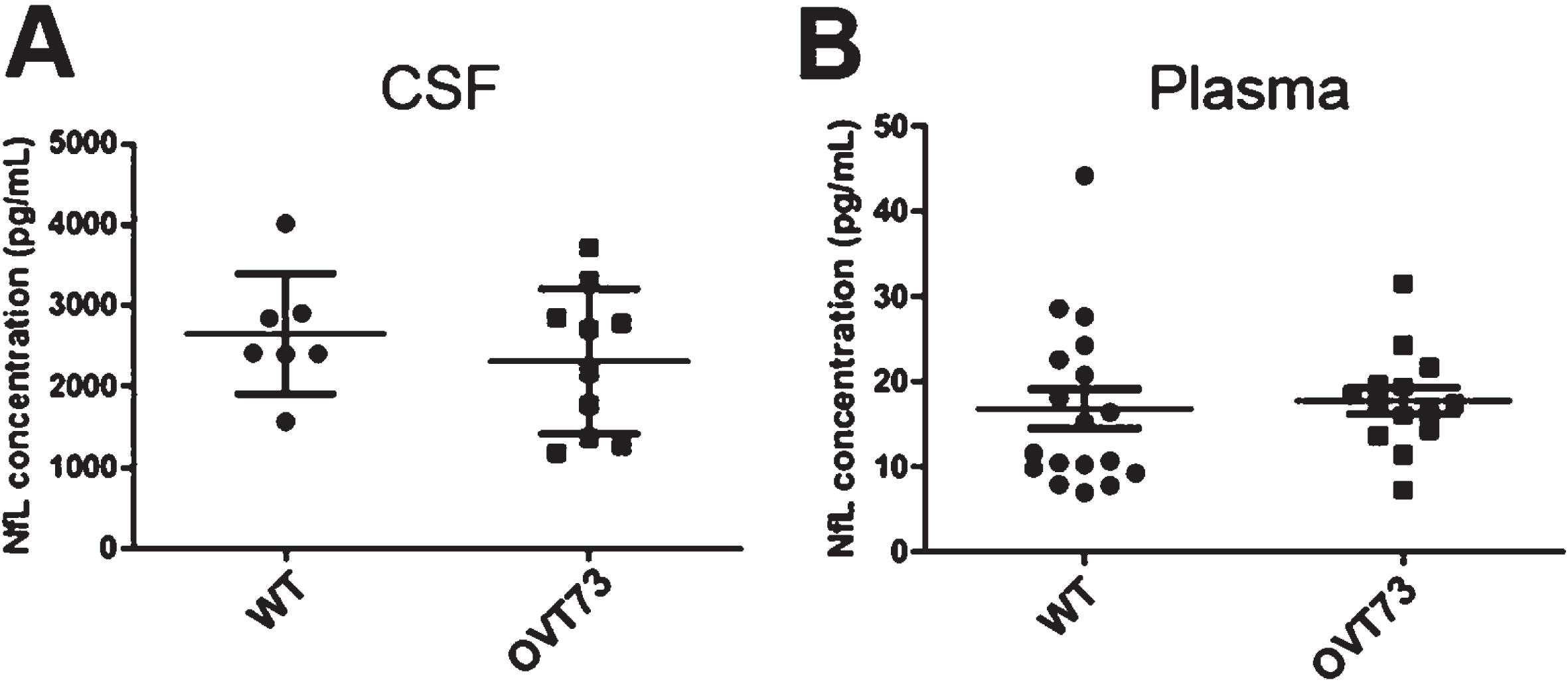
NfL levels in CSF and plasma are not significantly different between 10-year-old WT and OVT73 HD sheep. A) CSF samples were diluted 100 times, and NfL levels were assessed in the HD-X analyzer. No difference was observed between WT (n = 7) and OVT73 HD (n = 10) (p = 0.4244). Comparable results were obtained in plasma (B) (p = 0.92). 16x dilution was performed in WT (n = 18) and OVT73 HD (n = 16). An unpaired t-test was applied after verifying for normal distribution and similar variance.
DISCUSSION
In this study we sought to establish if OVT73 HD transgenic sheep expressing the full-length human HD gene exhibit synaptic changes in brain or altered levels of NfL in plasma and CSF that would indicate axonal pathology. Our western blot results revealed low expression of mHTT in all regions examined. Despite changes in levels of synaptosomal proteins, including some that function presynaptically, axonal dysfunction was not sufficient to trigger detection of altered NfL levels in biofluids suggesting the OVT73 HD sheep remain in a prodromal stage at 10 years of age.
Most proteins changed in OVT73 sheep synaptosomes were reduced, and some of them such as DARPP32, SCN4B, PDE10A, and calmodulin, were identified to be lower in mouse models of HD [14, 17–23]. Transcripts encoding PDE10A, SCN4B, and calmodulin are known to be downregulated in HD mice and may account for lower protein levels seen in synaptosomes [14, 20]. Calmodulin is also sequestered by cross-linking to transglutaminase in nuclear aggregates of HD patient postmortem brain [34] and interacts more with mHTT than with WT HTT [35], pointing to additional factors that could affect calmodulin levels and function in HD synaptosomes. Calmodulin has a role in exocytosis [36, 37]. Interestingly, studies in HD R6/1 mice revealed that several calcium/calmodulin-dependent protein kinases (CaMK) are hypophosphorylated, suggesting their decreased activity is linked to calmodulin [38].
Proteins changed in OVT73 HD sheep were affected by age and brain regions. For example, PDE10A and DARPP32 were significantly reduced in the caudate of OVT73 sheep at 5 years, similar to HD mice at 6 months. However, SCN4B, a protein reduced in caudate putamen of 6-month-old HDQ140/Q7 mice, was only significantly reduced in the motor cortex of 10-year-old OVT73 sheep. A few proteins (Na+/K+ ATPase and VAMP1) were also increased in piriform cortex of 10-year-old OVT73 HD sheep but not in 5-year-old sheep or in caudate or putamen. In 12-month-old HDQ140/Q140 mice, increases in levels of VAMP2 have been noted in caudate putamen [23]. Increases in synaptosomal proteins, particularly in older animals, may be a compensatory response or could result from decreased turnover due to impaired autophagy or proteosomal degradation. Sapp et al. found that protein changes in synaptosomes of HD mouse cortex differed from those in caudate putamen [15]. Our results in OVT73 HD sheep and those in mouse suggest that analysis of synaptosomes derived from regions of cortex as well as caudate and putamen may reveal more and different protein changes.
Mouse models of HD that develop molecular, cellular, and behavioral phenotypes vary in levels of mHtt expression. In knock-in mouse models of HD, the levels of HD transcript expression are equal to WT, and in the most widely studied full-length HD transgenic mouse models (YAC128 and BACHD), levels of mHTT are like the endogenous HTT [13, 39]. In contrast, OVT73 HD sheep have markedly lower levels of the human HD transgene than the endogenous sheep HTT, resulting in only 21.6–23.6% mHTT compared to endogenous sheep HTT protein in the synaptosome fraction of the 5- and 10-year-old OVT73 HD sheep. This may partly explain the mild phenotypes described in this model [8]. Nevertheless, some protein changes do occur in OVT73 HD sheep synaptosomes. Also, mHTT protein levels are sufficient to form some cytoplasmic and nuclear inclusions in the brain of 18-months and 3-year-old OVT73 HD sheep [8]. Importantly, the OVT73 sheep transgene lacks the human DNA sequence necessary to generate HTT1a which is thought to be the main aggregate-forming mHTT species [40, 41]. Although OVT73 HD sheep have mild molecular changes and express low levels of mHTT, they do develop a progressive circadian abnormality [2] and some neurologic and white matter changes [11]. Moreover, HD sheep were useful for testing lowering of mHTT as a therapeutic strategy. AAV delivery of miRNA targeting human HTT into OVT73 sheep caudate putamen was found to be safe and effective for reducing mHTT levels in the caudate putamen [42].
NfL is a cytoskeletal protein localized to myelinated axons, and its elevation in biofluids of HD patients is thought to arise from axonal pathology in brain and therefore serve as a biomarker for disease [43, 44]. Although white matter changes in 10-year-old OVT73 HD sheep have been reported [11], we found no change in NfL levels in the plasma or CSF of OVT73 HD sheep compared to WT even though some axonal proteins were changed such as VGLUT1, calmodulin, and SCN4B. VGLUT1, which was reduced in caudate and putamen, is known to reside on terminals of cortical afferents [45] suggesting corticostriatal projections are affected in the OVT73 HD sheep. Synaptic proteins could be investigated as potential biomarkers for pathology progression in sheep biofluids as they are in AD patients [46].
Altogether our results support a prodromal state for OVT73 sheep up to 10 years of age, associated with modest synaptic protein changes and unaltered NfL levels. Multiple factors may contribute to slow disease progression. These include low levels of mHTT, a CAG repeat number (69 CAGs) that may not undergo somatic expansion, the presence of CAA motifs at the end of the CAG repeat that hinder somatic expansion [13, 48], and the absence of human intronic sequence in the transgene that prevent formation of a potentially toxic HTT1a fragment [40, 41]. Understanding the role of these factors in disease progression beyond the prodromal state will be important for guiding HD therapeutics.
Footnotes
ACKNOWLEDGMENTS
This work was supported by CHDI Foundation, Inc (JSC6367) and the Dake family fund to MD.
CONFLICT OF INTEREST
KBK-G spouse owns less than 0.1% stock in the following companies: Advanced Microdevices, Aveo Pharmaceuticals, Inc, Boston Scientific Corporation, Bristol-Myers Squibb Company, Cisco Systems, Inc., Fate Therapeutics, GE Healthcare Life Sciences, Generex Biotechnology Corporation, Idera Pharmaceuticals, Inc., Nante Health, Neurometrics, Inc., NuGenerex, Repligen Corporation, Sesen Bio, Inc., T2 Biosystems, and Vericel Corporation.
Other authors have no declarations of interest to report.
DATA AVAILABILITY
Data sharing is not applicable to this article as no datasets were generated or analyzed during this study.