
Abstract
Neuroplasticity is a fundamental yet relatively unexplored process that can impact rehabilitation of lower extremity (LE) movements. Transcranial magnetic stimulation (TMS) has gained widespread application as a non-invasive brain stimulation technique for evaluating neuroplasticity of the corticospinal pathway. However, a majority of TMS studies have been performed on hand muscles, with a paucity of TMS investigations focused on LE muscles. This perspective review paper proposes that there are unique methodological challenges associated with using TMS to evaluate corticospinal excitability of lower limb muscles. The challenges include: (1) the deeper location of the LE motor homunculus; (2) difficulty with targeting individual LE muscles during TMS; and (3) differences in corticospinal circuity controlling upper and lower limb muscles. We encourage future investigations that modify traditional methodological approaches to help address these challenges. Systematic TMS investigations are needed to determine the extent of overlap in corticomotor maps for different LE muscles. A simple, yet informative methodological solution involves simultaneous recordings from multiple LE muscles, which will provide the added benefit of observing how other relevant muscles co-vary in their responses during targeted TMS assessment directed toward a specific muscle. Furthermore, conventionally used TMS methods (e.g., determination of hot spot location and motor threshold) may need to be modified for TMS studies involving LE muscles. Additional investigations are necessary to determine the influence of testing posture as well as activation state of adjacent and distant LE muscles on TMS-elicited responses. An understanding of these challenges and solutions specific to LE TMS will improve the ability of neurorehabilitation clinicians to interpret TMS literature, and forge novel future directions for neuroscience research focused on elucidating neuroplasticity processes underlying locomotion and gait training.
Introduction
Neuroplasticity, the capacity of the nervous system to adapt in response to experience, is a fundamental yet relatively unexplored process underlying rehabilitation of gait and lower limb movements (Kleim & Jones, 2008). Gait rehabilitation treatments intended to alter movement patterns are more likely to be effective if they are based on an understanding of and induce a change in the relevant brain circuitry (Belda-Lois et al., 2011). However, neuroplasticity principles typically applied during gait rehabilitation are derived largely from experiments involving hand muscles (Butler, Kahn, Wolf, & Weiss, 2005; Butler & Wolf, 2003; Cohen et al., 1998; Kleim & Jones, 2008; Rossi, Hallett, Rossini, & Pascual-Leone, 2009; Rossini et al., 2015). Upper limb tasks require greater dexterity, fractionation of movements, and a fine gradation of muscle force. In contrast, lower limbs engage in tasks requiring relatively gross control of muscle force, emphasis on joint stabilization and postural support, as well as inter-limb and inter-joint coordination during gait (Joseph, 1985; Mille, Simoneau, & Rogers, 2014). There is a paucity of research investigating neural correlates underlying rehabilitation of lower extremity (LE) movements and rehabilitation. Without an in-depth understanding of how cortical and spinal circuitry is modulated following gait retraining, developing evidence-based gait rehabilitation treatments that induce long-lasting improvements in walking function will remain challenging. This perspective review summarizes challenges and opportunities associated with evaluation of neuroplasticity in corticospinal circuitry controlling LE muscles. We focus on transcranial magnetic stimulation (TMS), a popular non-invasive brain stimulation technique for probing the excitability of descending motor pathways.
The role of the corticospinal tract in lower extremity motor control
Multiple structures throughout the neuraxis are implicated in the control and coordination of LE movements, posture, and locomotion (Charalambous, Bowden, & Adkins, 2016b; Joseph, 1985; Lacquaniti, Ivanenko, & Zago, 2012). Although circuitry within the brain stem and spinal cord can maintain rhythmic EMG activity during gait, cortical sensorimotor areas play a significant role in locomotor control (Barthelemy, Grey, Nielsen, & Bouyer, 2011). The primary motor cortex (M1) influences ongoing EMG activity during gait (Barthelemy et al., 2011; Bonnard, Camus, Coyle, & Pailhous, 2002). Cortical centers help modify the motor plan to avoid obstacles, change direction and speed, respond to perturbations, and enable cognitive or motor dual-tasks during locomotion (Barthelemy et al., 2011; Charalambous, Bowden, & Adkins, 2016a; Rothwell, 1997). In addition to the increasing evidence for the role of M1 in the control of posture and locomotion (Papegaaij, Baudry, Negyesi, Taube, & Hortobagyi, 2016; Papegaaij, Taube, Baudry, Otten, & Hortobagyi, 2014; Papegaaij, Taube, Hogenhout, Baudry, & Hortobagyi, 2014; Papegaaij, Taube, et al., 2016), other descending pathways such as reticulo-spinal and vestibulo-spinal tracts are relevant to lower limb motor control, and merit more investigation (Prentice & Drew, 2001; Ting et al., 2015). Additionally, fast and slow conducting pyramidal neurons may play a differential task-specific role during LE movements (Stout & Beloozerova, 2013). In light of the multiplicity and complexity of neural structures implicated in the control of LE movements and locomotion, recognizing the strengths, limitations, and extent of specificity of the measurement technique used to evaluate this circuitry becomes an important consideration.
TMS, a non-invasive technique to evaluate the excitability of the corticospinal pathway
TMS has made substantial contributions to our understanding of the human motor system (Groppa, Oliviero, et al., 2012; Hallett, 2007; Reis et al., 2008; Rossini et al., 2015). During TMS, a magnetic field generated by a coil positioned over the head induces an electric field in cortical neural elements located underneath the coil (Klooster et al., 2016; Rossi et al., 2009; Rossini et al., 2015). During TMS targeting M1, which is the focus of the present manuscript, TMS-induced motor-evoked potentials (MEPs) can be recorded using surface electromyography (EMG) sensors attached on the skin overlying the muscle of interest (Groppa, Oliviero, et al., 2012; Rothwell, 1997) as well as other muscles not directly targeted (Wolf, Butler, Alberts, & Kim, 2005). Two main factors influence how TMS acts upon a neural element (axon, dendrite, or cell body) within M1 - depth of the neural element from the surface of the head, and orientation of the neural element within the induced electric field (Wassermann, Epstein, & Ziemann, 2008). The strength of the induced electric field diminishes markedly with depth, limiting the use of TMS to stimulating superficial cortical targets (Deng, Lisanby, & Peterchev, 2013; Groppa, Oliviero, et al., 2012; E. Wassermann et al., 2008). The orientation of the neural element with respect to the direction of induced current influences both the efficacy of stimulation (stimulus intensity required to bring corticospinal neurons to firing threshold) and the combination of cortical elements that are stimulated (direct axonal stimulation versus trans-synaptic indirect stimulation) (Brasil-Neto et al., 1992; Di Lazzaro et al., 1998; Groppa, Oliviero, et al., 2012; Terao et al., 2000). In response to stimulation of M1, the earliest wave originates from direct activation of the axons of fast pyramidal tract neurons and is termed the “D” wave (Di Lazzaro & Rothwell, 2014; Groppa, Oliviero, et al., 2012; Kernell & Chien-Ping, 1967; Terao et al., 2000). The later waves require integrity of cortical grey matter, and are thought to originate from indirect, trans-synaptic activation of pyramidal tract neurons, termed “I” waves (Di Lazzaro & Rothwell, 2014; Groppa, Oliviero, et al., 2012; Kernell & Chien-Ping, 1967; Terao et al., 2000). Utilizing an optimum TMS coil orientation as well as increasing stimulation intensity can increase the probability of generating D-waves (Priori et al., 1993; Terao et al., 2000; Terao et al., 1994). Given these basic principles underlying TMS, we focus on the applicability of TMS to target M1 corticomotor neurons that project to LE muscles. In the context of the complex neural circuitry implicated in the control of LE movements, TMS probes the excitability of the corticospinal pathway (single-pulse TMS), facilitatory and inhibitory drive within the cortex (paired-pulse TMS), and can be paired with other electrophysiological techniques to evaluate motor control circuitry.
The use of TMS for studying lower limb musculature presents unique methodological challenges
There are methodological challenges associated with using TMS to evaluate corticomotor excitability of LE muscles. Additionally, for each challenge, potential methodological solutions and avenues for future research directions are suggested (Groppa, Oliviero, et al., 2012; Rossini et al., 2015).
Challenge associated with location of the LE motor homunculus deep within the inter-hemispheric fissure
During LE TMS, one obvious challenge is that the M1 cortical representation of most LE muscles (i.e. the LE homunculus) is located at the midline of the brain within the depths of the inter-hemispheric fissure (Smith, Stinear, Alan Barber, & Stinear, 2017; Terao et al., 2000; Terao et al., 1994). Compared to M1 representation of upper limb muscles, representations for muscles surrounding the foot (e.g. abductor hallucis), ankle (e.g. tibialis anterior, soleus, gastrocnemius), and knee (e.g. rectus femoris, vastus lateralis) joints are located at a greater depth with respect to the TMS coil. Because the strength of the TMS-induced magnetic field attenuates rapidly with distance from the surface of the scalp (Deng et al., 2013), the TMS-induced electric field may not reach adequate depth to activate cortical motoneuronal cells controlling ankle and foot muscles. Additionally, in contrast to the upper limb, LE muscles are involved in more gross control, with a greater proportion of motor units driven by larger motor neurons with higher activation thresholds (Smith et al., 2017). Additionally, while the deeper location of the LE M1 representation is a challenge related to LE TMS (Priori et al., 1993; Terao et al., 2000; Terao et al., 1994; Weiss et al., 2013), differences in the orientation of corticospinal axons within M1 for UE compared to LE muscles may also contribute to the higher TMS intensities required to elicit MEPs from LE muscles. Inter-individual variability in the shape and size of gyri and sulci within LE M1 may further contribute to the variability and challenge in finding motor threshold and hotspot locations for LE muscles (Inoue, Shimizu, Nakasato, Kumabe, & Yoshimoto, 1999; Sobel et al., 1993; Thielscher, Opitz, & Windhoff, 2011; Weiss et al., 2013). Thus, compared to upper limb muscles, higher TMS intensities may be required to elicit MEPs from the LE, further reducing the probability of eliciting measurable MEPs during LE TMS (Chen et al., 1998; Deng et al., 2013; Smith et al., 2017). These challenges are compounded further for individuals with neurological lesions, such as stroke and spinal cord injury, because of elevated motor thresholds (McDonnell & Stinear, 2017; McKay, Lee, Lim, Holmes, & Sherwood, 2005).
Solution 1: Utilization of specialized coils for improved access to deep-seated LE M1 representations
Previous TMS investigations focusing on LE muscles have attempted to overcome the challenge imposed by the deeper location of LE M1 by applying specialized TMS coils. The double-cone coil has been used most frequently (Jayaram & Stinear, 2009; Madhavan, Rogers, & Stinear, 2010; Papegaaij, Baudry, et al., 2016; Papegaaij, Taube, Hogenhout, et al., 2014; Papegaaij, Taube, et al., 2016). Other studies have used the batwing coil (Barthelemy et al., 2011; Needle, Palmer, Kesar, Binder-Macleod, & Swanik, 2013; Palmer, Hsiao, Awad, & Binder-Macleod, 2016; Palmer, Hsiao, Wright, & Binder-Macleod, 2017; Palmer, Needle, Pohlig, & Binder-Macleod, 2016; Palmer, Zarzycki, Morton, Kesar, & Binder-Macleod, 2017) and flat figure of eight coil (Smith et al., 2017). The angulation in the batwing and double cone coils helps to increase the depth of penetration of the induced electric field (Deng et al., 2013; Klooster et al., 2016). Coil shape and size influence the depth and focality of cortical tissue targeted during TMS (approximately 5 cm2 for figure of eight coils), and merit more careful investigation for LE muscles (Deng et al., 2013; Klooster et al., 2016). The limited magnetic field available in current commercial stimulators is another limiting factor, and a provision to enhance magnetic field strength would be advantageous for LE TMS studies (Kammer, Beck, Thielscher, Laubis-Herrmann, & Topka, 2001). Of course, similar to upper limb muscles, the probability of eliciting a measurable MEP from LE muscles increases by instructing the participant to maintain low level background activation (e.g. 10% of the EMG during a maximal voluntary contraction) in the target muscle (Rossini et al., 2015; Rossini, Zarola, Stalberg, & Caramia, 1988). Because the influence of increasing background activation on TMS-evoked MEP amplitude may differ for leg muscles versus hand muscles, monitoring and accurately reporting the background EMG within and across sessions will improve the rigor of LE TMS studies.
Differences in optimal coil orientation for eliciting MEPs when stimulating upper and lower limb muscles need more consideration (Volz, Hamada, Rothwell, & Grefkes, 2015). For the upper limb, posterior to anterior cortical current flow perpendicular to the central sulcus produces the lowest motor thresholds (Brasil-Neto et al., 1992). In contrast, for LE muscles, medial to lateral cortical current flow (away from the inter-hemispheric fissure) may produce a lower motor threshold and shorter latency (Rossini et al., 2015; Smith et al., 2017; Terao et al., 1994, 2000) (Fig. 1). Recent LE TMS investigations reported successful elicitation of MEPs from leg muscles using the conventional flat figure-of-eight coil placed in the medio-lateral orientation (Smith et al., 2017). Furthermore, the relative composition of D-waves and I-waves in the TMS-induced volley may vary with coil shape and orientation, stimulation intensity, and across different lower limb muscles (Rossini et al., 2015; Smith et al., 2017; Terao et al., 1994, 2000; Volz et al., 2015). The composition of D- and I-waves in the TMS-induced volley is important due to their influences on motor threshold as well as the susceptibility of MEP amplitude to conditioning using paired-pulse protocols (Wassermann et al., 2008).
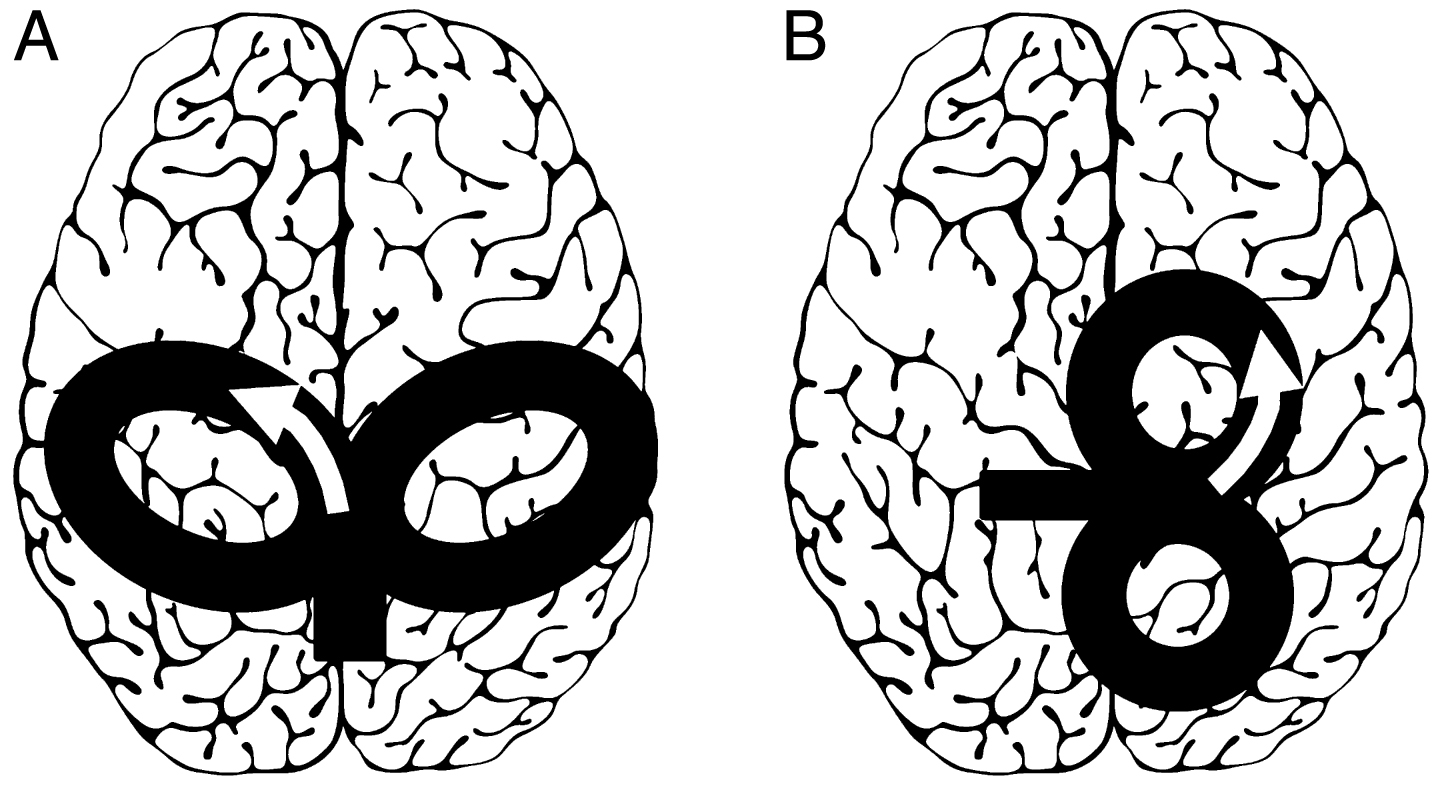
Schematic showing TMS coil orientations for targeting LE muscles (figure of eight coils). The schematics demonstrate coil position for stimulation of the right hemisphere leg area (to elicit MEPs from the ankle or foot muscle of the left leg). The arrows designate direction of induced current within the cortex (A) Several LE TMS studies have utilized specialized coils such as the double-cone or batwing coil positioned 0–2 cm posterior to the vertex and oriented such that the cortical current direction is posterio-anterior. (B) Recent studies and TMS consensus papers suggest that the optimal coil orientation for LE TMS may involve rotating the coil 90 degrees such that the cortical current direction is medio-lateral (B). Adapted from Rossini et al. (2015) and Groppa et al. (2012).
Challenges encountered with targeting individual lower limb muscles during TMS
The M1 representations of LE muscles spanning the same lower extremity joint (e.g. tibialis anterior and soleus) and adjacent joints (e.g. soleus and rectus femoris) are located in close proximity within the LE homunculus. During LE TMS, the TMS-induced cortical currents may ‘spill over’ to adjacent muscle representations of the ipsilateral limb. Due to the lack of focality of the TMS-induced magnetic field, TMS causes simultaneous activation of corticospinal neurons innervating agonist, synergist, and antagonist LE muscles. Therefore, during TMS, isolating cortical neurons that control distinct LE muscles may not be possible. For instance, given the limited focality of the TMS-induced magnetic field, when stimulating the M1 hotspot of the right tibialis anterior at supra-threshold intensities, both the tibialis anterior and soleus cortical neurons for the right ankle would likely be activated (Fig. 2, upper left). Similarly, due to their geometrical proximity within the homunculus, cortical neurons controlling muscles around the knee joint (e.g. rectus femoris, biceps femoris) could be activated simultaneously when targeting the tibialis anterior M1 hotspot (Fig. 2, lower left). A majority of TMS studies record MEPs from one or two targeted muscles, providing little information about the extent of concurrent activation of synergist and antagonist muscles during TMS. The lack of focality, although a challenge, can also provide a methodological opportunity for the design of TMS studies evaluating training-induced neuroplasticity in LE muscles. For example, when using TMS as a probe to evaluate the corticomotor excitability of ankle muscles, simultaneous TMS-induced descending volleys are generated in upper motor neurons controlling knee and hip muscles, and MEPs will likely be elicited from proximal LE muscles (e.g. rectus femoris) during TMS targeting the ankle. Thus, if EMG recordings are made from multiple LE muscles (those targeted and not targeted by the intervention), the use of LE muscles not directly targeted by the intervention as a ‘control’ muscle may be accomplished relatively easily during LE intervention studies.
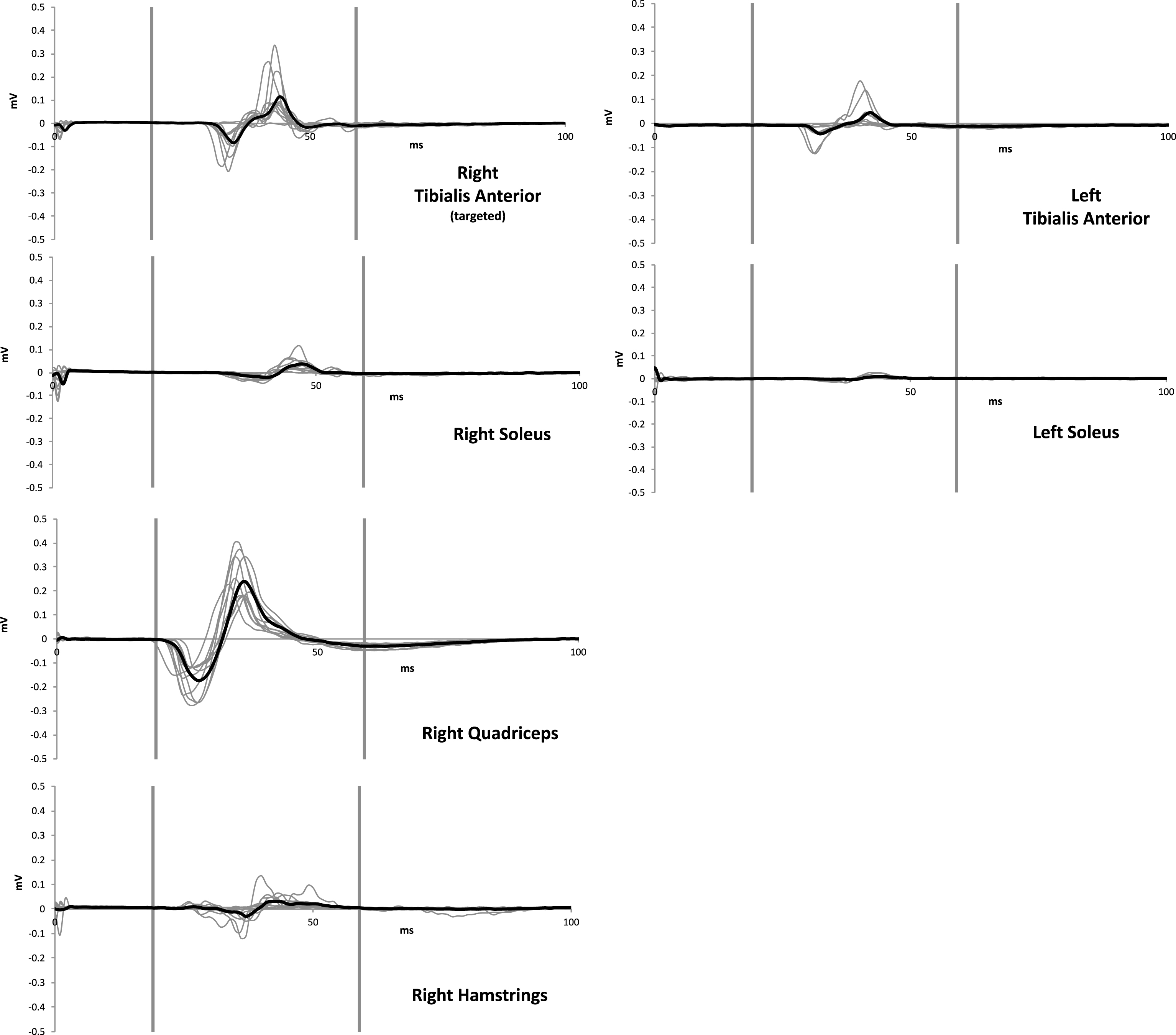
TMS to LE M1 may elicit MEPs from multiple leg muscles. Data from a representative neurologically unimpaired participant demonstrating MEPs from multiple LE muscles elicited by a supra-threshold TMS pulse (120% resting motor threshold) delivered using Magstim stimulator and custom batwing coil (oriented to induce postero-anterior cortical current flow) over the hotspot of the right tibialis anterior muscle. TMS elicited measurable MEPs from adjacent muscles of the right lower extremity ipsilateral to the right tibialis anterior (right quadriceps, right hamstrings, right soleus) as well as homologous muscles of the contralateral lower extremity (left tibialis anterior, left soleus). For each graph, the two vertical lines delineate the MEP window. The bold line represents the average of 10 individual MEPs (gray lines). Data from the left quadriceps and hamstrings were not collected due to constraints in the number of EMG channels available. These data showcase problem 2 (challenge with targeting individual muscles during LE TMS). These data also illustrate solution 2b (concurrent measurement from multiple LE muscles).
When applying TMS to evaluate cortical excitability of upper limb muscles, there is a comparatively greater distance between the targeted M1 and contralateral M1 representations. In contrast, cortical neurons projecting to homologous muscles for the ipsi- and contra-lateral LE (e.g. for the left and right tibialis anterior) are located in close proximity on the adjoining banks of the inter-hemispheric fissure. Due to this geometric proximity of the bilateral LE M1 motor representations, when attempting to target preferentially the M1 area controlling the contralateral leg, the non-focal TMS-induced magnetic field likely activates motor cortex in both hemispheres simultaneously (Fig. 2, upper row). During LE TMS targeting one leg, these bilateral MEP responses are observed often, but rarely systematically documented (Ackermann, Scholz, Koehler, & Dichgans, 1991; Smith et al., 2017). Proximity of the left and right M1 representations for LE muscles may impose a challenge to the study of neural processes mediating cross-limb transfer of training- or neuromodulation-related plasticity (Ruddy & Carson, 2013; Stockel, Carroll, Summers, & Hinder, 2016), and neural mechanisms of bilateral LE training (Hinder, Carroll, & Summers, 2013).
Solution 2a. Systematic investigation regarding cortical motor maps and hotspots for LE muscles
Very little information is available regarding the extent of overlap between hotspot locations of adjacent (e.g. tibialis anterior and soleus) and distant (e.g. soleus and quadriceps) lower limb muscles. Theoretically, based on the diameter and shape of conventionally used TMS coils (e.g. flat figure of eight, double cone, batwing), distribution of the TMS-induced electric field within the cortex, limited focality of the TMS-induced electric field, and proximity of different movement representations within the LE homunculus, adjacent LE muscles probably have comparable hotspots. However, previous studies often use procedures that assumed distinct hotspot locations for different lower limb muscles, such as tibialis anterior and soleus (Ackermann et al., 1991; Obata, Sekiguchi, Nakazawa, & Ohtsuki, 2009). There is a need for experimental studies to determine whether the same hotspot can be used for obtaining TMS-evoked MEPs from multiple LE muscles simultaneously. Other questions requiring further study relate to whether optimal coil orientation should be individualized for each participant as part of the hotspot determination procedure (Smith et al., 2017).
Previous studies, predominantly on upper limb muscles (Byrnes et al., 1998; Corneal, Butler, & Wolf, 2005; Ngomo, Mercier, & Roy, 2013; Pearce, Thickbroom, Byrnes, & Mastaglia, 2000; Wolf et al., 2004), report reorganization of cortical motor maps following immobilization (Liepert, Tegenthoff, & Malin, 1995), amputation (Schwenkreis et al., 2001), musculoskeletal injuries (Ward et al., 2015), and stroke rehabilitation (Sawaki et al., 2014). Similar to findings in upper limb, corticospinal plasticity associated with LE rehabilitation may be accompanied by changes in the size or location of motor maps. Recent LE TMS mapping studies showed symmetrical maps between left and right hemispheres for the vastus lateralis (Al Sawah et al., 2014) and rectus femoris muscles (Ward et al., 2016) in able-bodied individuals. Another LE mapping study showed a more dispersed pattern of tiblialis anterior hotspot locations in individuals post-stroke compared to healthy individuals (Sivaramakrishnan, Tahara-Eckl, & Madhavan, 2016). More comprehensive LE mapping studies are required to determine the extent to which TMS-derived corticomotor maps for LE muscles overlap between muscles spanning the same joint (tibialis anterior and soleus), muscles spanning adjacent joints (ankle and knee), and homologous muscles of the contralateral limb (left and right tibialis anterior). Additionally, more clarification is needed regarding the interpretation and limitations of motor maps and TMS hotspot location (Wassermann, McShane, Hallett, & Cohen, 1992). The hotspot is the coil position and orientation on the scalp that allows the TMS-induced magnetic field to best activate the targeted corticospinal neurons (e.g. pyramidal tract neurons controlling the right tibialis anterior muscle), providing the lowest threshold and shortest latency of TMS-evoked responses (Malcolm et al., 2006; Meincke, Hewitt, Batsikadze, & Liebetanz, 2016; Rossini et al., 1994). An increase in TMS-derived motor map area may signify greater cortical excitability, a larger area of functionally-relevant muscle representation, or that the targeted muscle representation can be activated from more remote scalp locations (Butler et al., 2005; Wassermann et al., 1992; Wolf et al., 2004).
Solution 2b. Simultaneous recordings from multiple lower extremity muscles
A simple, innovative, and informative methodological modification for LE TMS studies involves obtaining simultaneous EMG recordings from multiple LE muscles. For example, studies focused on evaluating changes in corticomotor excitability of the right tibialis anterior should also consider obtaining concurrent recordings from the right soleus, rectus femoris, gastrocnemius, biceps femoris, as well as similar muscles in the left (contralateral) lower limb (Fig. 2). EMG measurement from multiple muscles, instead of the commonly used approach of recording EMG from one target muscle, may allow us to capitalize on the proximity of LE muscle M1 representations. A majority of previous LE studies have used TMS to evaluate cortical excitability from one or two muscles (usually tibialis anterior or quadriceps) (Ackermann et al., 1991; Jayaram et al., 2012; Luft et al., 2005; Madhavan et al., 2010; Obata et al., 2009; Smith et al., 2017). Very few LE TMS investigations have recorded from more than 2 muscles or muscles spanning multiple LE joints (Chen et al., 1998; Smith et al., 2017; Tan, Shemmell, & Dhaher, 2016). While we may not anticipate a change in corticomotor excitability of other LE muscles not directly targeted by an LE rehabilitation intervention, the anatomical organization of LE M1 homunculus invites the opportunity to record from and evaluate multiple muscles. Future studies comprising simultaneous recordings from multiple LE muscles may reveal new insights about the organization and plasticity of circuitry controlling LE movements. For example, if a study’s primary focus is to evaluate whether an intervention comprising functional electrical stimulation (FES) delivered to ankle dorsiflexor muscles increases corticomotor excitability of the stimulated (tibialis anterior) muscle representation, the conventional TMS methodological approach would entail recording EMG from only the paretic tibialis anterior muscle. In contrast, we posit that simultaneous recordings from multiple LE muscles may provide additional insights about FES-induced corticomotor plasticity. Recordings from the contralateral or non-paretic tibialis anterior can determine whether the FES-intervention modulates the excitability of the non-lesioned corticospinal pathway. Concurrent recordings from the antagonist (soleus) muscle of the paretic leg may help determine whether FES-induced corticomotor plasticity is specific to the targeted muscle or it also induces modulation of corticomotor excitability of the antagonist. Further, inclusion of recordings from proximal muscles (e.g. rectus femoris) of the paretic leg may help determine whether FES induces a generic upregulation of corticomotor excitability in the paretic limb cortical representations. Recording from multiple muscles may be particularly advantageous for LE TMS studies because of the proximity of different LE representations, justifying the additional time required to setup EMG and determine threshold for more than one muscle.
Solution 2c. Methodological modifications to separate contra- and ipsi-lateral responses
A few studies have used clever methodological protocols to overcome the challenge associated with the close proximity of the left and right lower limb M1 representations within the inter-hemispheric fissure. For example, TMS stimuli systematically delivered at the vertex compared to 1–2 cm on either side (left and right) of the vertex may influence the probability of preferential activation of the contralateral leg muscles (Jayaram et al., 2012; Madhavan et al., 2010; Tan et al., 2016). Using this approach in individuals post-stroke, Madhavan et al. deduced stronger ipsilateral conductivity for the paretic ankle muscle if the slope of the TMS-elicited recruitment curve for the paretic ankle muscle was steeper during TMS delivered to the non-lesioned hemisphere (ipsilateral to paretic ankle) compared to the lesioned hemisphere (contralateral to paretic ankle) (Madhavan et al., 2010). In conjunction with the use of TMS to measure corticospinal tract conductivity, the study also used neuro-imaging (diffusion-tensor imaging) to evaluate corticospinal tract structural integrity on the lesioned and non-lesioned hemisphere (Madhavan et al., 2010). However, more studies should validate such indices of ipsilateral conductivity in able-bodied individuals. Studying the effects of rehabilitation programs on neurophysiologic and neuro-imaging metrics post-stroke may provide innovative insights about LE rehabilitation mechanisms. Interesting perspectives about LE TMS may also be gained by evaluating how the suppression of ongoing EMG in response to sub-threshold TMS (Papegaaij, Taube, et al., 2016) or activation-related modulation of intra-cortical inhibition (Papegaaij, Baudry, et al., 2016) vary across different LE muscles of the contra- and ipsilateral leg.
Challenges encountered due to differences in motor control circuitry and function between upper and lower limb musculature
In addition to factors related to geometrical proximity of muscle representations within LE M1, the unique organization of LE motor control circuitry limits the specificity of TMS-induced responses. Consistent with LE tasks involving gross or non-fractionated motor control, there may be considerable overlap in cortical circuits controlling adjacent leg muscles. Similarly, descending projections onto the spinal lower motoneuron pools of adjacent LE muscles also overlap (Jankowska, Padel, & Tanaka, 1975; Machii et al., 1999; Mang, Clair, & Collins, 2011). Furthermore, monosynaptic excitatory cortical projections, presumably common in upper limb muscles, may be fewer or weaker (have reduced synaptic strength) in LE muscles, such as the soleus (Bawa, Chalmers, Stewart, & Eisen, 2002; Brouwer & Ashby, 1990, 1992; Geertsen, Zuur, & Nielsen, 2010). Lower limb TMS studies have largely focused on the tibialis anterior (Beaulieu, Masse-Alarie, Ribot-Ciscar, & Schneider, 2017; Jayaram et al., 2012; Jayaram & Stinear, 2009; Sivaramakrishnan et al., 2016; Smith et al., 2017; Stinear & Hornby, 2005) and knee extensor muscles (Al Sawah et al., 2014; O’Leary, Morris, Collett, & Howells, 2015; Ward et al., 2016); with a few studies on the abductor hallucis muscle (Yen, Wang, Liao, Huang, & Yang, 2008). Variability in the strength of corticospinal connections across different lower limb muscles can markedly affect TMS-induced responses, and merits more systematic study.
As another example, due to reciprocal inhibition, during and prior to dorsiflexion of the ankle, soleus H-reflexes are depressed (Crone, Hultborn, Jespersen, & Nielsen, 1987). In contrast, during voluntary dorsiflexion, TMS-induced soleus MEPs are facilitated (Geertsen et al., 2010). For LE muscles, voluntary contraction of an agonist may be accompanied by preceding facilitation of antagonists by a subcortical motor program, enabling a rapid change in direction of movement during functional motor tasks (Geertsen et al., 2010). The effects of activating the antagonist on the TMS-evoked MEP recorded from the agonist, may be pertinent for TMS studies focused on both LE and UE muscles. Future LE TMS investigations on the influence of activation of synergist, antagonist, proximal joint, and contralateral leg muscles on TMS-evoked MEPs may provide insights about circuitry controlling LE movements as well yield more clearly defined modes of delivery for LE TMS.
Solution 3a. Commonly used TMS outcomes may need to be modified for LE musculature
Most TMS methodology and outcome measures were developed in upper limb muscles (Groppa, Oliviero, et al., 2012; Hallett, 2007; Rossini et al., 2015). There is a need to modify and validate TMS methodological protocols (e.g. hotspot determination, threshold determination, number of target muscles recorded) for LE muscles. Similarly, existing TMS outcome measures (e.g. motor threshold, MEP amplitude, recruitment curves) (Groppa, Oliviero, et al., 2012; Hallett, 2007; Kesar et al., 2017; Rossini et al., 2015) may need to be modified for use in lower limb muscles. For example, related to our recommendation to include concurrent recordings from multiple muscles during LE TMS studies, procedures used to determine motor threshold could be revisited when multiple muscle recordings are being made and/or variations in background EMG activation are observed. Due to greater depth of the LE M1, greater magnitude of background activation (e.g. greater than 10% maximal voluntary activation) may be required to elicit measurable MEPs from LE muscles. Also, surface EMG amplitude is influenced by multiple factors other than muscle activation, such as muscle depth from surface, skin impedance, thickness of subcutaneous tissues, etc. and these factors may vary across muscles more so for the LE (e.g. gluteus maximus and tibialis anterior) than UE (biceps femoris and flexor digitorum indicis). Thus, perhaps there is a need to revisit the practice of using an absolute MEP amplitude (e.g. 100μV) as an active motor threshold criterion for different muscles (e.g. tibialis anterior and gluteus maximus) (Groppa, Oliviero, et al., 2012; Rossini et al., 2015). For instance, active motor threshold criterion could be referenced to background EMG during the pre-TMS period, or as a percentage of the maximal muscle response (M-wave) elicited by peripheral nerve stimulation (Urbin, Ozdemir, Tazoe, & Perez, 2017b). Furthermore, during TMS experiments involving simultaneous recordings from multiple LE muscles, additional time may be required to estimate threshold for individual muscles as a basis for interpreting MEP data recorded from multiple muscles with respect to the threshold intensity for each muscle. Paired pulse techniques, commonly used to quantify intra-cortical facilitation (ICF) and short-interval intracortical inhibition (SICI), are used for hand muscles (Groppa, Oliviero, et al., 2012; Hallett, 2007; Kujirai et al., 1993; Rossini et al., 2015). During paired pulse TMS, the modulation of the test MEP amplitude by a conditioning pulse is purported to be mediated by intra-cortical mechanisms for hand and forearm muscles (Kujirai et al., 1993). In a comparative study of proximal and distal muscles of upper and lower limbs, Chen et al. demonstrated that even for LE muscles with a higher motor threshold, ICF and ICI are mediated by cortical mechanisms (Chen et al., 1998). However, there is a need to determine whether the unique geometry of the LE M1 representation influences paired-pulse measurements. Again, recordings from multiple muscles during LE paired-pulse experiments may prove advantageous. Additionally, to elicit measurable control MEPs, paired-pulse methodologies for LE muscles may need to be implemented with the target muscle maintaining a low-level background EMG activation (paired-pulse measurements in the active versus resting state).
Solution 3b. Systematic evaluation of the influence of testing posture and muscle activation on TMS-evoked responses
More studies should elucidate factors influencing TMS-evoked responses from LE musculature. For instance, the influence of testing posture (sitting, standing, tandem stance), and muscle activation state on TMS-evoked MEPs of LE muscles requires more investigation. When considering the effect of muscle activation on LE cortical excitability, little is known about the extent to which MEPs obtained from the right tibialis anterior are modulated by activation of the antagonist (right soleus), synergist (right fibularis longus), a proximal muscle (e.g., right rectus femoris), contralateral homologous (left tibialis anterior), or contralateral non-homologous muscles (left soleus, left quadriceps). When studying of the role of posture on cortical excitability, there are few lower limb TMS studies that have recorded TMS-evoked MEPs in the standing posture. Previous findings lack consensus about whether TMS-evoked MEPs of LE muscles increase during standing (Ackermann et al., 1991; Baudry, Collignon, & Duchateau, 2015; Lavoie, Cody, & Capaday, 1995; Obata et al., 2009; Papegaaij, Baudry, et al., 2016; Papegaaij, Taube, et al., 2016; Soto, Valls-Sole, Shanahan, & Rothwell, 2006).
Modulation of corticomotor excitability of LE muscles with variations in postural challenge or task difficulty may differ with age and neurologic pathology. These factors could influence design and interpretation of TMS studies focused on LE muscles and tasks. For example, short-interval intra-cortical inhibition of the soleus muscle was greater during supported standing (Papegaaij, Baudry, et al., 2016) and during standing on a foam rather than a rigid surface (Papegaaij, Taube, Hogenhout, et al., 2014). Subthreshold TMS-induced EMG suppression of ongoing soleus EMG activity, purported to be mediated through modulation of intra-cortical circuitry, was greater during an unsupported forward leaning postural task (Papegaaij, Taube, et al., 2016). When compared to TMS recordings in the seated posture, LE TMS data recorded during standing, challenging postural tasks, and walking may provide more sensitive indices of neuroplasticity. More studies investigating the effects of posture (standing, seated, supine) and trunk muscle activation on LE muscle MEPs are needed. Similarly, the test-retest reliability of TMS-evoked MEPs recorded during standing, dynamic postural tasks, and walking need to be demonstrated.
M1 may be involved in the control of both voluntary (trunk flexion) and postural tasks (postural stabilization during voluntary shoulder flexion). Thus, the use of TMS-derived measures of corticospinal excitability of trunk, UE, and LE muscles during postural control tasks is another area of interest (Chiou, Hurry, Reed, Quek, & Strutton, 2018; Fujiwara, Tomita, & Kunita, 2009; Kantak et al., 2013; Masse-Alarie, Neige, Bouyer, & Mercier, 2018). By measuring TMS-evoked MEPs in trunk muscles during a range of tasks, Chiou et al. showed that corticospinal excitability of the erector spinae muscle was greater during dynamic anticipatory posture-related adjustments than during static tasks requiring postural and goal-directed voluntary activity (Chiou, Gottardi, Hodges, & Strutton, 2016). A recent study used TMS to evaluate corticomotor excitability of trunk muscles (erector spinae and rectus abdominis) during anticipatory postural adjustments, which involve activation of trunk muscles in association with volitional UE activity (deltoid), and suggested that postural adjustments may have a cortical origin (Chiou et al., 2018).
Solution 3c. Evaluation of the reliability of TMS-evoked responses for LE muscles
TMS-evoked responses show a high degree of variability from trial to trial (within a session) as well as across multiple sessions (Darling, Wolf, & Butler, 2006; Julkunen et al., 2009; Jung et al., 2010; Kiers, Cros, Chiappa, & Fang, 1993; Malcolm et al., 2006; Ngomo, Leonard, Moffet, & Mercier, 2012). Numerous factors contribute to experimental variability of TMS-evoked responses, such as background activation, muscle mechanics, coil-positioning error, visual and cognitive attention, EMG sensor placement, oscillatory activity in neural networks, and habituation (Darling et al., 2006; Julkunen et al., 2009; Jung et al., 2010; Kiers et al., 1993; Malcolm et al., 2006; Ngomo et al., 2012; Tallent et al., 2012b). Although factors contributing to the variability of TMS-evoked MEPs merit more study in both upper limb and LE muscles, the research gap related to test-retest variability of TMS-derived outcomes is more marked for LE muscles. Tallent et al. conducted a rigorous evaluation of test-retest reliability of TMS-derived measures during dynamic contractions of the tibialis anterior, and showed good reliability for a majority of TMS-derived measures (Tallent et al., 2012b). However, interestingly, the coefficient of variation decreased from the 2nd to 3rd day of testing, suggesting that a familiarization session may improve repeatability (Tallent et al., 2012b). Recent evaluations of the reliability of TMS outcomes for the tibialis anterior muscle in the ipsi- and contralesional hemispheres of post-stroke individuals support the use of active motor threshold and MEP latency for diagnostic and prognostic applications, while emphasizing the need to evaluate test-retest reliability of TMS outcomes in large sample sizes (Beaulieu et al., 2017; Cacchio et al., 2011). Testing procedures and activation history can also influence TMS outcomes. Repetitive motor activity induces transient increases in corticomotor excitability in hand muscles (Classen, Liepert, Wise, Hallett, & Cohen, 1998). In lower limb muscles, undertaking a systematic evaluation of the magnitude and recovery time course of post-contraction MEP facilitation would help with designing improved experimental protocols that minimize the influence of testing procedures on MEP amplitude and variability (Tallent et al., 2012a). Early and long-term neural responses to strength training can coexist with the neural plasticity induced by motor skill or gait training (Aagaard, Simonsen, Andersen, Magnusson, & Dyhre-Poulsen, 2002; Selvanayagam, Riek, & Carroll, 2011). With the increasing prevalence of longitudinal studies evaluating rehabilitation-induced neuroplasticity, there is a need to document physiological or test-retest variability of TMS-derived outcome measures from LE muscles (Tallent et al., 2012a, 2012b). Similar to other LE clinical outcomes, such as over ground gait speed and timed up and go test score, documentation of the minimal detectable change for TMS-derived outcome measures is warranted (Beaulieu et al., 2017; Tallent et al., 2012a, 2012b).
Solution 3d. Pairing TMS with other electrophysiological techniques
Experimental approaches that pair TMS with other electrophysiological techniques, such as peripheral nerve stimulation, can provide innovative insights about activity in specific descending pathways implicated in LE motor control. For example, pairing TMS with peripheral nerve stimulation at varying latencies can elucidate how descending circuits influence the spinally-mediated Hoffman’s reflex (H-reflex) response. TMS-conditioning of the H-reflex has been used to evaluate short-latency facilitation (SLF) and long-latency facilitation (LLF) (Geertsen, van de Ruit, Grey, & Nielsen, 2011; Gray, Sabatier, Kesar, & Borich, 2017). SLF is purported to reflect the strength of fast-conducting monosynaptic descending projections, while LLF evaluates the strength of the indirect, polysynaptic descending projections (e.g. cortico-reticular, cortico-vestibular) onto the lower motor neuron pool (Geertsen et al., 2011; Gray, Sabatier, et al., 2017). Given the potentially greater role played by polysynaptic descending pathways in LE motor control, measures such as LLF may be particularly valuable for LE studies. Paired-associative stimulation (PAS), a technique based on principles of Hebbian spike timing-dependent plasticity, can be used both for evaluation as well as neuro-modulation (Stefan, Kunesch, Cohen, Benecke, & Classen, 2000). During PAS, repeated pairing or association of a TMS pulse with a peripheral nerve stimulus may induce short-term changes in corticospinal excitability (Stefan et al., 2000). A few studies have demonstrated that facilitatory and inhibitory PAS can be induced in LE muscles during seated and walking conditions (Stinear & Hornby, 2005; Urbin, Ozdemir, Tazoe, & Perez, 2017a; Uy, Ridding, Hillier, Thompson, & Miles, 2003). The challenges and opportunities for further inquiry related to the use of PAS and TMS-conditioning of H-reflexes in LE muscles are consistent with those described earlier – the TMS-induced electric current may activate bilateral motor cortices (and other muscle representations of the ipsilateral hemisphere) simultaneously, leading to bilateral descending volleys influencing spinal motor neuron pools, thus confounding interpretation of study findings. Accordingly, techniques such as PAS, SLF, and LLF are particularly valuable for LE muscles because the pairing of the TMS pulse (which has limited focality for LE muscles) with a peripheral nerve stimulation pulse (which targets a specific muscle/nerve) may help partially overcome the disadvantage related to lack of specificity during LE TMS. Additionally, when TMS-elicited MEPs cannot be elicited from LE muscles in individuals with elevated motor thresholds due to neurological insults (e.g. stroke, cerebral palsy), the combination of TMS with electroencephalography (EEG) may be a valuable technique to study direct cortical responses to TMS (Gray, Palmer, Wolf, & Borich, 2017; Sato, Bergmann, & Borich, 2015). Finally, evaluation of the role of cortico-cortical connections and excitability of areas other than M1 (such as the dorsal premotor cortex, parietal cortex) in LE motor control is needed (Groppa, Schlaak, et al., 2012; Koch & Rothwell, 2009; Parmigiani, Zattera, Barchiesi, & Cattaneo, 2018). Similarly, the role of the cerebellum, which is critical for both motor and cognitive control as well as adaptation and motor learning, in training-induced neuroplasticity of LE muscles is warranted (Grimaldi et al., 2016; Jayaram, Galea, Bastian, & Celnik, 2011; Spampinato, Block, & Celnik, 2017).
The fundamental methodological challenges associated with the use of TMS to evaluate corticospinal excitability of LE musculature have been summarized. We discuss methodological solutions that can help alleviate these challenges. There are several gaps in our knowledge related to organization of corticospinal circuitry controlling LE movements and locomotion. TMS experimental protocols and outcome measures may need to be modified for use in LE muscles. A simple yet informative methodological modification for LE TMS studies, involving concurrent recordings from multiple LE muscles, should be considered. Additionally, systematic TMS investigations focused on LE muscles are needed to determine how the testing posture and activation state influence TMS-elicited MEPs from LE muscles. TMS studies focused on the LE are important for elucidating the neuroplasticity correlates underlying rehabilitation of gait and posture. Information gained using TMS can be markedly enhanced by the use of complimentary electrophysiological (peripheral nerve stimulation, electroencephalography) and neuro-imaging techniques. An understanding of the challenges and solutions specific to LE TMS will improve the ability of neurorehabilitation clinicians to interpret the TMS literature, and forge novel future directions for neuroscience research focused on elucidating neuroplasticity processes underlying locomotion and gait training. Better comprehension of neuroplasticity processes underlying LE motor control can inform the development of innovative rehabilitative, pharmacological, and neuro-modulation treatments to maximize LE function in individuals with neurological impairments.
Conflict of interest statement
The Authors declare that there are no conflicts of interest.