
Abstract
Background
Epithelial-to-mesenchymal transition (EMT) of human peritoneal mesothelial cells (HPMCs) contributes to fibrotic thickening of the peritoneum that develops in patients on peritoneal dialysis (PD). The process is thought to be largely mediated by transforming growth factor-beta (TGF-β). As TGF-β has also been implicated in senescence of HPMCs, we have performed an exploratory study to examine if senescent HPMCs can undergo EMT.
Methods
Omentum-derived HPMCs were rendered senescent by repeated passages in culture. Features of EMT were assessed by immunostaining and quantitative polymerase chain reaction (qPCR) at various stages of the HPMC lifespan and after treatment with or without TGF-β. The motility of HPMCs was assessed in a scratch wound migration assay.
Results
Replicative senescence of HPMCs was associated with a gradual increase in the constitutive expression of EMT markers, including increased production of extracellular matrix proteins. However, senescent HPMCs also retained epithelial cell features such as cytokeratin, calretinin, and E-cadherin and showed decreased, rather than increased, motility. In contrast, exposure to TGF-β resulted in an up-regulation of mesenchymal markers and down-regulation of epithelial markers. Such effects of TGF-β occurred both in young and senescent cells, although they were less pronounced in senescence.
Conclusions
Senescence of HPMCs is associated with spontaneous development of several EMT features. At the same time, senescent HPMCs preserve epithelial cell-like characteristics and are less prone to develop a full EMT phenotype in response to TGF-β. These observations may support the concept of cellular senescence being antagonistically pleiotropic with regard to EMT.
Transforming growth factor-β induces EMT in HPMCs (4) by engaging various mitogen-activated protein kinases (MAPKs) (5,6), as well as the nuclear transcription factor-κB (NF-κB) (7) and heat shock proteins (8). Adenovirus-mediated overexpression of TGF-β in the rat peritoneum leads to the up-regulation of genes involved in EMT and migration of mesothelial cells from the peritoneal surface monolayer into the sub-mesothelial stroma (9).
The significance for PD of phenotypic conversion of HPMCs is that it is associated with an increase in the production of pro-fibrotic and pro-angiogenic mediators impinging on structure and function of the peritoneal membrane (1). However, the mechanism by which EMT is initiated during PD remains unclear. It has been demonstrated that PD fluids with high concentration of glucose degradation products can induce EMT both during short-term direct exposure in culture and, to a lesser extent, after chronic PD exposure in vivo (10,11).
In addition to being a mediator of EMT, TGF-β is critically involved in cellular senescence, which emerged as an important process of tissue remodeling (see [12] for a recent review). In this respect, TGF-β has been shown to mediate such aspects of HPMC senescence as growth arrest, cell hypertrophy, and expression of senescence-associated β–galactosidase (SA-β–Gal). Both HPMCs that senesce in vitro and HPMCs explanted from older donors secrete increased amounts of TGF-β, and the magnitude of TGF-β release correlates inversely with the lifespan of HPMCs in culture (13).
Given these roles of TGF-β, some kind of a relationship between cellular senescence and EMT is likely to exist. On the one hand, it is thought that by secreting increased amounts of bioactive molecules, senescent cells can promote EMT in neighboring cells (14). Indeed, it has been demonstrated that mesothelioma cells that entered senescence in response to chemotherapeutics became capable of inducing EMT in naïve non-senescent mesothelioma cells (15). On the other hand, the overexpression of EMT-controlling transcription factors inhibited growth suppressors and allowed the cells to bypass senescence (3,16). Conversely, inhibition of these factors resulted in cell cycle arrest, senescence (17), and insensitivity to TGF-β–driven EMT (18).
While it appears that TGF-β may act as a driver of both EMT and cellular senescence in HPMCs, it is not clear what determines the choice of response to TGF-β. To shed some light on this relationship, we have carried out experiments to compare the ability of pre-senescent and senescent HPMCs to undergo TGF-β–induced EMT in culture.
Methods
All chemicals, unless otherwise stated, were purchased from Sigma-Aldrich (St Louis, MO, USA). Tissue culture plastics were from Nunc (Roskilde, Denmark).
Cell Culture
Human peritoneal mesothelial cells were isolated by enzymatic digestion of the specimens of omentum removed from consenting patients undergoing elective abdominal surgery. The procedure was essentially as described elsewhere (19). Freshly isolated cells were identified as pure mesothelial by their typical cobblestone appearance at confluence and uniform positive staining for cytokeratin and calretinin, and negative staining for fibroblast-specific protein-1 (FSP-1) and von Willebrand factor. Immunostaining procedures were performed as previously described (20). Cells were maintained in the Earls'–buffered M199 medium supplemented with L-glutamine (2 mM), penicillin (100 U/mL), streptomycin (100 μg/mL), hydrocortisone (0.4 μg/mL), and 10% vol/vol fetal calf serum (Invitrogen, Karlsruhe, Germany).
Induction of Cell Senescence
Replicative senescence of HPMCs was induced by serial passages, as earlier described in detail (21). Cells were considered senescent when they stopped increasing in number over 4 weeks, showed distorted morphology, and stained positively for senescence-associated β–galactosidase (SA-β–Gal). Cytochemical assessment of SA-β–Gal was performed using the Senescence β–Galactosidase Staining Kit (Cell Signaling Technology, Beverly, MA, USA), according to the manufacturer's instructions. Human peritoneal mesothelial cells reach senescence in vitro typically after 6 – 10 passages (21). Thus, in order to monitor changes that occur in HPMCs with time in culture, the cells were assessed at their first passage (young cells), third to fifth passage (middle-passage cells), and last passage (senescent cells).
Induction of EMT
To induce EMT, HPMCs were incubated for 72 hours in normal culture medium supplemented with 1 ng/mL of human recombinant TGF-β1 (R&D Systems, Minneapolis, MN, USA). Such an exposure was previously shown to effectively induce EMT in HPMCs (4,22).
Scratch Wound Healing Assay
The assessment of cell wound closure was performed as described in detail elsewhere (23,24). Briefly, HPMC monolayers were scratch-wounded with a pipette tip, gently washed with medium to remove resulting debris and placed in an incubator coupled to an Axio Observer D1 inverted microscope (Zeiss, Oberkochen, Germany). Cells were maintained in standard culture medium until wound closure. Cell images were acquired by time-lapse microscopy at 60-minute intervals and analyzed using the AxioVision Rel. 4.6.3 image analysis software (Zeiss). The rate of remesothelialization was calculated by measuring reduction in the denuded area over time.
Immunostaining
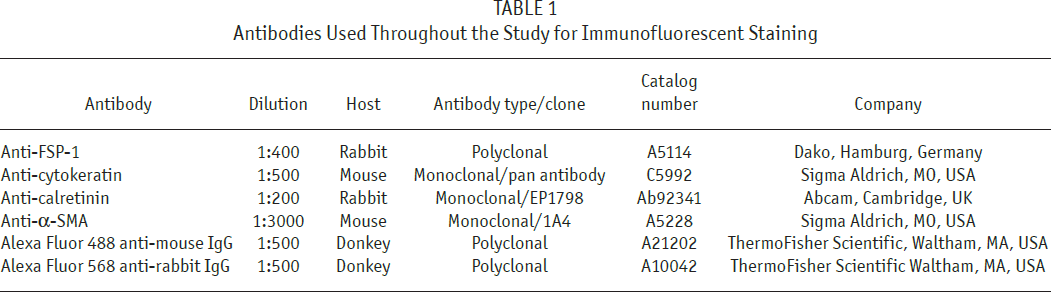
Cells were grown on Lab-Tek Chamber Slides (Nunc) and fixed with 3.7% paraformaldehyde. Then, cells were permeabilized with 0.1% Triton X-100, blocked with 5% bovine serum albumin (BSA), and incubated with appropriate primary antibodies for 1 hour at 37°C (Table 1). Antibodies were diluted in phosphate buffered saline (PBS) containing 0.1% BSA and 0.05% Tween 20. The slides were then incubated with appropriate fluorescent Alexa dye-labeled secondary antibodies in PBS and counter-stained with 4’,6-diamidino-2-phenylindole (DAPI) nuclear stain (all from Invitrogen/ThermoFisher Scientific, Waltham, MA, USA). Positively stained cells were counted by 2 independent observers in 5 randomly selected fields under a confocal microscope (LSM710 Zeiss Confocal Microscope) at 400x magnification, and expressed as a percentage of all DAPI-stained nuclei.
Antibodies Used Throughout the Study for Immunofluorescent Staining
Gene Expression Analysis
Expression of target genes was assessed with reverse transcription and quantitative polymerase chain reaction (qPCR). Total RNA was extracted with RNA Bee (Tel-Test, Friendswood, TX, USA), purified with the RNeasy kit (Qiagen, Hamburg, Germany), quantified, and reverse-transcribed into complementary DNA (cDNA) using RNA PCR Core Kit (Applied Biosystems, Foster City, CA, USA). Quantitative PCR was carried out on a Light Cycler 2.0 using a SYBR Green labeling kit (Roche, Mannheim, Germany) and specific primers for target genes (Table 2). The results were calculated by the cycle threshold method and normalized for the endogenous reference (histone H3).
Polymerase Chain Reaction Primer Pairs Used Throughout the Study

Statistical Analysis
All experiments were performed in triplicate using cells from separate donors. Data are presented as mean ± standard error (SE).
Results
As previously described (25), replicative senescence of HPMCs was accompanied by a gradual loss of their uniform cobblestone appearance. Instead, senescent cells became greatly enlarged, irregular and vacuolated, and stained extensively for SA-β–Gal (Figure 1). Despite these profound changes in morphology, senescent cells still expressed epithelial and mesothelial cell markers—cytokeratin and calretinin (Figure 2). However, they also began to express increasingly mesenchymal markers—α–smooth muscle actin (α–SMA) and fibroblast-specific protein (FSP)1. At the mRNA level, senescent HPMCs showed an increased expression of the transcription factor Snail, collagen 1, and fibronectin, and—interestingly—of E-cadherin (Figure 3A and B). The levels of TGF-β mRNA did not change considerably with senescence.
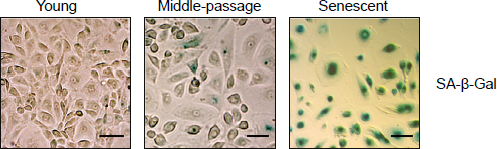
Expression of senescence-associated β–galactosidase SA-β–Gal) by HPMCs. Cells at different stages of their lifespan in culture were analyzed by cytochemistry for SA-β–Gal (magnification 40x, the scale bar corresponds to 100 μm). HPMC = human peritoneal mesothelial cell.
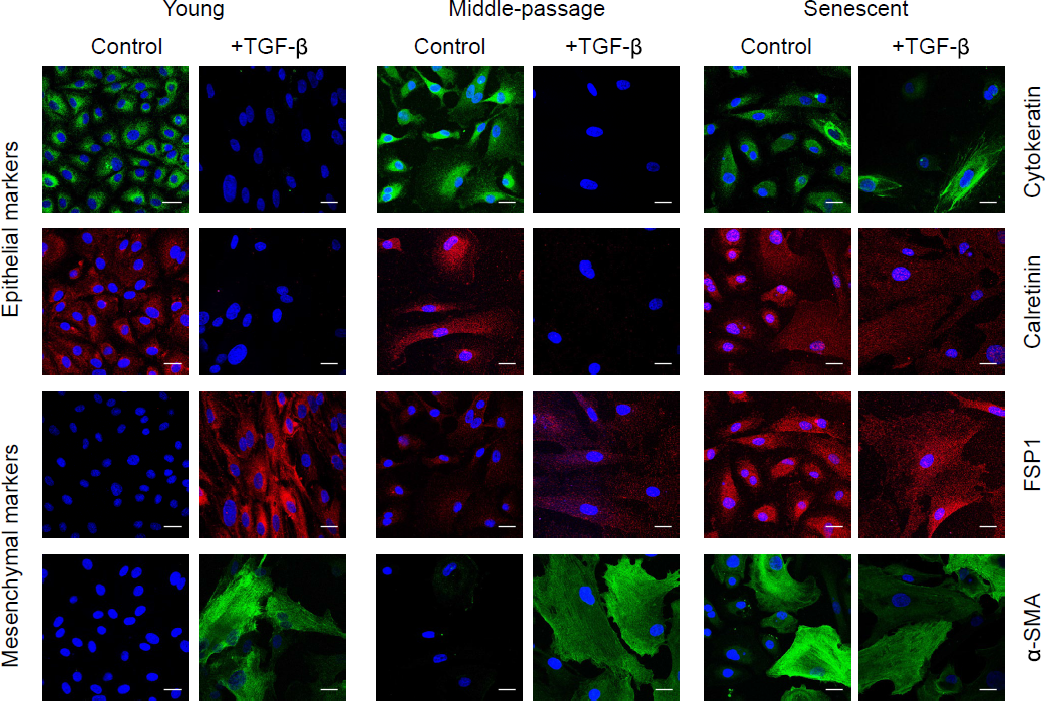
Expression of epithelial and mesenchymal markers by HPMCs. Exemplary images of young, middle-passaged, and senescent cells immunostained for a panel of biomarkers expressed either constitutively (Control) or after stimulation with TGF-β1 (+TGF-β), as described in Methods. Cell nuclei were counterstained with DAPI (blue). Magnification 400x, scale bar = 25 μm. HPMC = human peritoneal mesothelial cell; TGF = transforming growth factor.
In order to assess whether senescence of HPMCs impacts their ability to undergo full EMT in response to TGF-β, the cells were treated with TGF-β at different stages of their lifespan in culture (Figures 2 and 3). Young HPMCs responded to TGF-β by developing typical features of EMT – inducing Snail, down-regulating epithelial cell markers (cytokeratin, calretinin, E-cadherin), and up-regulating markers of myo-fibroblastic activity (FSP-1, α–SMA, collagen 1, fibronectin, TGF-β). A similar pattern of changes was also seen in middle-passage and senescent cells. However, there was a trend towards these effects of TGF-β becoming less pronounced, as expressed as a fold change in mRNA expression (Figure 3C). Notably, the induction of Snail and down-regulation of E-cadherin, cytokeratin, and calretinin was less evident in senescent cells.
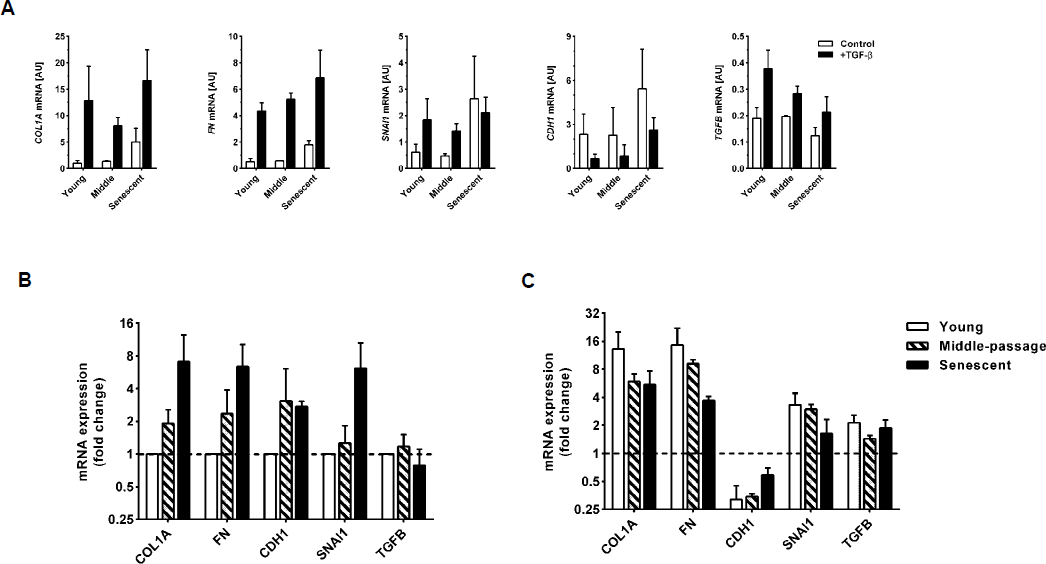
Expression of mRNAs for selected EMT markers in HPMCs undergoing senescence in the presence or absence of TGF-β. A) absolute expression levels as assessed by RT-qPCR; B) mRNAs expressed constitutively by HPMCs at different stages of senescence; the data are presented as a fold change from values seen in young cells; C) mRNAs expressed by HPMCs treated with TGF-β at different stages of senescence; the data are presented as a fold change from values recorded in the absence of TGF-β in young, middle-passaged, and senescent cells, respectively. All data were derived from 3 separate experiments with cells from different donors. EMT = epithelial-to-mesenchymal transition; HPMC = human peritoneal mesothelial cell; TGF = transforming growth factor; mRNA = messenger ribonucleic acid.
As EMT is a key step in mesothelial cell migration during wound healing, the rate of remesothelialization was assessed following scratch injury. These experiments demonstrated that the process of wound closure was delayed in cultures of senescent HPMCs (Figure 4).
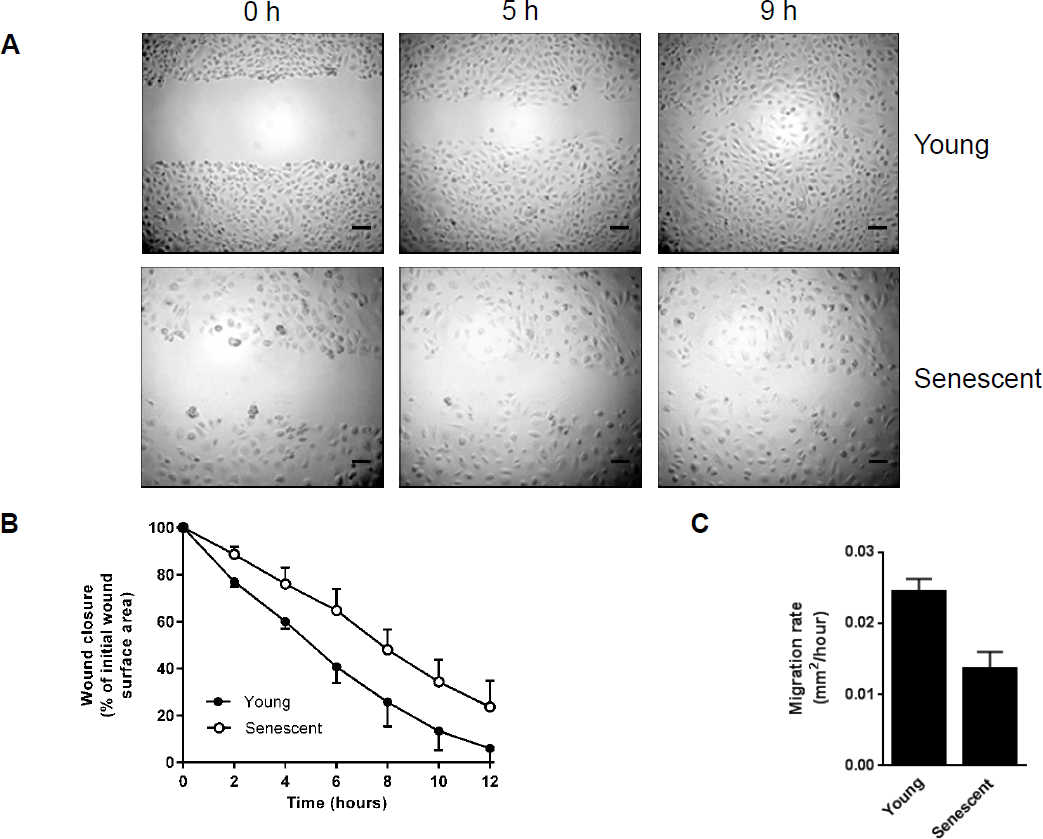
Wound healing in HPMCs undergoing senescence. A) Representative images of scratch wound healing in young (top) and senescent (bottom) HPMCs (magnification 10x, the scale bar corresponds to 100 μm); B) the rate of wound closure expressed as a percentage of the initial wound surface area; C) the rate of cell migration calculated as wound surface area (μm2) covered per hour. All data were derived from 3 separate experiments with cells from different donors. HPMC = human peritoneal mesothelial cell.
Discussion
The main observation of the present study is that HPMCs undergoing replicative senescence in culture acquire some phenotypic features consistent with EMT. In particular, they up-regulate Snail, a key transcriptional regulator of EMT. Moreover, they start expressing α–SMA, a marker of myofibroblasts, and FSP-1, a robust marker of fibroblasts and cells undergoing fibroblastic conversion (26). Accordingly, they also increase the expression of extracellular matrix proteins, collagen1, and fibronectin.
Intriguingly, however, having developed these EMT features, senescent HPMCs did not entirely lose an epithelial cell structure and still expressed typical epithelial cell proteins such as cytokeratin, calretinin, and E-cadherin. Expression of the latter was even increased, which could be attributed to great enlargement of senescent cells (21). Nevertheless, the observation is surprising given that it was associated with increased expression of Snail, a transcriptional repressor of E-cadherin (27). It may suggest that senescent HPMCs did not undergo full EMT and displayed an intermediate phenotype with mesenchymal markers being already expressed and epithelial markers not yet down-regulated (28).
As TGF-β is thought to be necessary for both the early and late phases of EMT (29), we assessed the effect of TGF-β on EMT at various stages of senescence. It turned out that the expression of genes involved in EMT displayed a very similar pattern in young, middle-passaged, and senescent cells. However, the effects of TGF-β in senescent cells appeared to be not so prominent as in young cells. Notably, both young and senescent cells responded to TGF-β by down-regulating E-cadherin and cytokeratin, albeit the effect seen in senescent cells was less evident than in young cells. These data may suggest that features of EMT present in senescent cells developed in a manner independent of TGF-β. Indeed, we did not observe an increase in TGF-β mRNA in senescent cells. Since earlier reports showed a significant increase in the release of active TGF-β by senescent HPMCs (13), our observations may point to a significant role of post-transcriptional mechanisms in increasing TGF-β release by senescent HPMCs.
We have also detected that senescent HPMCs displayed decreased motility and migratory capacity, which led to delayed wound healing. In this respect, previous observations in animals have shown that the development of “incomplete” EMT with reduced mesothelial cell mobilization and a non-invasive phenotype may be mediated by platelet-derived growth factor B (30). More recently, it has been shown that EMT-activated and dispersed cells migrate faster compared with epithelial biomarker-expressing cells with preserved contacts with neighboring cells (31).
Our data may thus suggest that cellular senescence hampers the development of a full EMT phenotype in HPMCs by retaining their epithelial features. The fact that high expression of E-cadherin and cytokeratin occurred in the presence of increased Snail expression indicates that senescence repressed the Snail-mediated effects. Thus, given the proposed role of EMT in peritoneal fibrosis, HPMC senescence could act as an anti-fibrotic mechanism. In this respect, it has been demonstrated that senescence of activated stellate cells can limit fibrosis of the liver (32). However, this concept may be at odds with the observation of increased expression of collagen and fibronectin by senescent HPMCs. These seemingly contradictory effects could be explained by antagonistic pleiotropy of senescent cells (33). On the one hand, by restraining EMT, cellular senescence of HPMCs can inhibit the development of peritoneal fibrosis. On the other hand, an increased production of extracellular matrix proteins may contribute to submesothelial fibrotic thickening. Another aspect of this apparent ambiguity could be an effect of TGF-β. By driving EMT, which is an essential part of wound healing, TGF-β promotes prompt wound closure. Indeed, it has been demonstrated that the treatment with TGF-β enhances cell motility and accelerates scratch wound healing in young mesothelial cells from rodents (34). However, by inducing cell senescence, TGF-β may indirectly delay tissue repair. Moreover, while small scratch wounds can close entirely by cell migration, the repopulation of larger injuries may require cell renewal. In this respect, mesothelial cell proliferation has been demonstrated to be clearly and dose-dependently inhibited by TGF-β (35). In addition, TGF-β displays seemingly paradoxical effects on the inflammatory response. On the one hand, TGF-β controls the course of acute bacterial peritonitis in rats by dampening pro-inflammatory cytokine release from mesothelial cells (36). On the other hand, by inducing cellular senescence, TGF-β contributes to the development of the senescence-associated secretory phenotype that is characterized by a chronic increase in pro-inflammatory mediators release (37).
The existence of senescent HPMCs in vivo is difficult to document. The presence of few SA-β–Gal-expressing mesothelial cells was detected in apparently normal human omentum (38). In the context of PD, it has been observed that some mesothelial cells shed from the peritoneal membrane into the dialysate may express senescent cell biomarkers SA-β–Gal and p16 (39). Mesothelial cells with senescence features have also been detected in peritoneal imprints taken from mice exposed to PD fluids and this effect has been attributed to osmotic agents in the dialysate (40). It has been hypothesized that dialysate glucose accelerates cell cycle progression and thus leads to earlier exhaustion of the growth capacity and/ or genotoxic stress that results in premature cell senescence so that potential defects are not passed to daughter cells (41). Although the regeneration of mesothelial cells is likely to involve several mechanisms (see [42,43] for review), a decrease in the proliferative potential of progenitor/parent cells may hamper mesothelial cell recovery from PD fluid-induced injuries and lead to depopulation of the mesothelial monolayer seen in peritoneal biopsies (44).
Conclusions
It is not clear to what extent the process of replicative HPMC senescence in vitro reflects the biological status of HPMCs in vivo. It is believed that the potential of explanted cells to grow in culture depends on the proliferative history of cells, which is influenced both by the calendar age of the donor and by the previous environmental stresses. This complexity may be reflected by large heterogeneity in telomere length that can be seen in freshly isolated HPMCs (25).
While the evidence of EMT was recorded in HPMCs isolated from the dialysis effluent (22), these cells were not analyzed for the senescence markers. Therefore, in future studies the simultaneous assessment of EMT and senescence in effluent-derived HPMCs and in peritoneal biopsies may help to define exact molecular mechanisms linking these 2 processes. Interestingly, recent lineage tracing experiments in rodents demonstrated that EMT of mesothelial cells did not contribute significantly to the accumulation of myofibroblasts following peritoneal injury (45). In view of our findings it would be interesting to see if mesothelial cells expressed senescence markers under these conditions.