
Abstract
In systemic sclerosis (SSc), the dysregulation of several molecular pathways seem to have a role in the disease pathogenesis. Either angiogenesis and vasculogenesis are disturbed and impaired, and an imbalance between angiogenic and angiostatic factors may be involved in the genesis and maintenance of vasculopathy. Aberrant immune system activation and function involves both B and T cells, as well as many different chemokines and cytokines. Particularly, chemokines are central to the initiation and maintenance of inflammatory responses as well as angiogenesis and fibrosis. Increased expression of several chemokines as CXCL4 (platelet factor 4), CXCL8 (IL8), CXCL5 (ENA-78), CCL5 (RANTS), CXCL9 (MIG), CCL24, CXCL10 IP-10), CXCL12, CXCL16 (SRPSDX), CCL2 (MCP-1), CCL19 (MIP-3β/ELC), CCL24 (Eotaxin 2), suggests a complex mechanism by which many immune cell types, including T cells, macrophages and neutrophils are recruited to the skin in SSc patients. Many of these chemokines have redundant roles, possibly to ensure recruitment of specific cell types. Several studies have shown a synergistic effect of combinations of these chemokines in cell recruitment, emphasizing the importance of understanding global chemokine expressions. urthermore, chemokines can be detected in peripheral blood compared with cytokines or growth factors. The utility of cytokines as biomarkers has been investigated but longitudinal studies are necessary to clarify their clinical utility for the evaluation of disease activity, therapeutic effects on skin sclerosis or interstitial lung disease and risk stratification of SSc patients. An effective therapeutic agent, able to interfere with complex chemokine networks, is warranted to attenuate perivascular inflammation, dysregulated angiogenesis and the evolution of skin and internal organ fibrosis, is the most ambitious goal for the scientific research of the future.
Introduction
Cell to cell and cell to extracellular matrix (EMC) communication are regulated by several molecules such as cytokines, chemokines and adhesion molecules that guarantee a dynamic equilibrium in physiological state. Dysregulation of this tightly controlled system could be responsible for inflammatory (arthritis), proliferative (cancers) and anti-proliferative (autoimmune) diseases (1, 2).
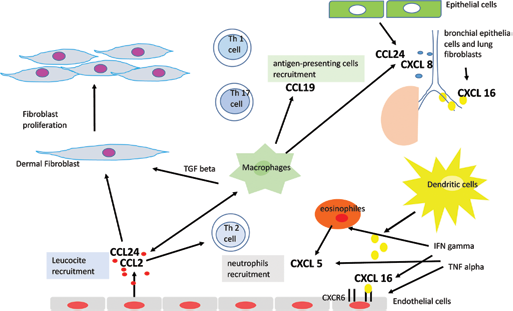
Angiogenic chemokines.
Systemic sclerosis (SSc) is a connective tissue disease characterized by fibrosis of the skin and of internal organs, cellular and humoral immunity abnormalities and pronounced alterations in the microvasculature (3). In SSc, many molecular pathways are dysregulated and have a role in its complex pathogenesis (4, 5). Either angiogenesis and vasculogenesis are disturbed and impaired, and an imbalance between angiogenic and angiostatic factors may be involved in the pathogenetic mechanisms leading to SSc vasculopathy (4, 5). In SSc, the vascular injury occurs very early in the disease, even before the onset of fibrosis (4, 5). In the early phases of the disease when tissue inflammation is predominant, it has been suggested that vessel permeability and injury coincide with the infiltration of immune cells, mainly consisting of T cells and macrophages localized at perivascular regions (6–7–8). The immune cell recruitment is facilitated by vascular endothelial cells which, upon injury, upregulate adhesion molecules such as vascular cell adhesion molecule 1 (VCAM-1), intercellular adhesion molecule 1 (ICAM-1) and E-selectin, ultimately leading to immune activation, and tissue damage (9). Aberrant immune system activation and function involves both B and T cells, as well as many different chemokines and cytokines. Excessive activation and polarization to Th2 cells is observed with consequent cytokine overproduction (interleukin [IL] 4, IL13, IL6) that promote fibroblast proliferation, collagen deposition and increase profibrotic transforming growth factor beta (TGFβ) (10–11–12). Additional, pro-inflammatory (IL1, tumor necrosis factor alpha [TNFα], IL 17A and IL 10) (13, 14) and pro-fibrotic (IL13, IL25, IL33) cytokines are upregulated in SSc patients (15–16–17). Chemokines, defined as cytokines inducing chemotaxis in nearby responsive cells, are central to the initiation and maintenance of inflammatory responses as well as angiogenesis (13). Consequently, the fine tuning of their expression at inflamed sites contributes to the resolution of the process, by promoting wound healing and tissue repair (13).
In SSc skin, several chemokines show upregulated expression: CXCL9 (MIG) and CXCL10 (IP-10), which recruit T cells (18–19–20); CCL2 (MCP-1), which recruit macrophages (21–22–23) and other chemokines such as CCL5 (RANTS), CXCL8 (IL8), CXCL5 (ENA-78) and CCL24 (Eotaxin 2), which recruit many immune cell types, including T cells, macrophages and neutrophils and up-regulate adhesion molecules expression, thus allowing diapedesis (9, 21, 24, 25). In this review, we report the literature on chemokines found in SSc and their position in the disease pathogenesis.
Chemokines and Systemic Sclerosis
Chemokines are small soluble proteins, not bigger than 8-10 kDa, that induce chemotactic activity in various cells, especially cells of the immune system, organizing T cell motility and migration in physiological immune responses as well as in pathological processes such as in chronic inflammatory and autoimmune conditions. This action is evident after binding to their specific receptors constitutively expressed on cells belonging to the immune system. Their target cells express respective and sometimes overlapping receptors (26). Inflammatory chemokines can be induced by cells exposition to pro-inflammatory cytokines or tissue damage or infection. In inflamed contexts, interaction of inflammatory chemokines with their receptors on leucocytes leads to inflammatory cell influx and accumulation at damaged or infected sites. Membership in family of chemokines is classified on the basis of the presence of variations of a conserved cysteine motif in the mature proteins. According to this classification, chemokines can be divided into subfamilies: C (one cysteine residue is missing), CC (first two cysteine residues are immediately adjacent in the primary structure), CXC (first two cysteine residues are separated by an amino acid) and CX3C (separation involves three amino acids) chemokines. This system is crucial in several biological functions, although its key role is in regulating tissue-specific leucocyte migration under both inflammatory and homeostatic conditions. Functionally, chemokines attract leukocytes into tissues and contribute to regulate angiogenesis, vascular proliferation and fibrosis contributing to manifestations of scleroderma. Together, the CC and CXC subfamilies constitute the majority of chemokines, whereas the C and CX3C chemokines represent a limited subpopulation of unconventional chemokines (27). The CXC chemokines can be subdivided into two groups according to the presence of the enzyme-linked receptor (ELR) motif or Glu-Leu-Arg. The ELR+ CXC chemokines such as CXCL16 and CXCL5 are potent promoters of angiogenesis (Tab. I), by binding the unique receptor CXCR6, as well as potent activators and chemoattractants for neutrophils, whereas the ELR-chemokines, such as CXCL4, CXCL9 and CXCL10, have angiostatic (Tab. II) properties by binding the CXCR3 receptor without exhibiting neutrophil chemoattractant activities (28). These different functions appear to be due to the presence of three different splice variants of CXCR3: CXCR3-A, CXCR3-B, and CXCR3-alt (29) and, in particular, the angiostatic role is mediated solely by CXCR3-B isoform (30, 31) (Tab. I, Fig. 1).
Chemokines with angiogenic activity in the pathogenesis of systemic sclerosis
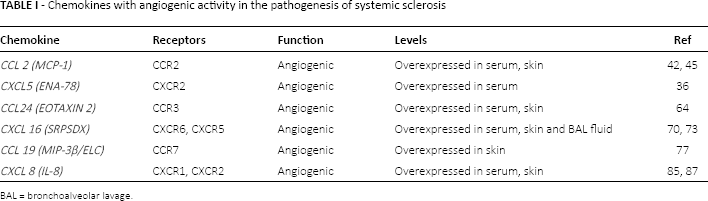
BAL = bronchoalveolar lavage.
Chemokines with angiostatic activity in the pathogenesis of systemic sclerosis

BAL = bronchoalveolar lavage.
In SSc, the inflammatory response is activated and chemokine alterations have been described and have been implicated through human, animal and in vitro studies in its pathogenesis. Endothelial cell injury, caused by ischemia-reperfusion, may induce inflammatory cell infiltration and subsequent cytokine production leading to tissue fibrosis. During this process, chemokines mediate leukocyte and mononuclear cell chemotaxis, cell activation and induce an interaction between leukocytes and fibroblasts with release of profibrotic growth factors (32, 33), engage in the perpetuation of the chronic damage and in the induction of fibrosis (21).
In particular, chemokines such as CCL2, CCL3, CCL4, CCL5, CCL7, CXCL8, CXCL12 initiate and maintain aberrant fibroblast function, which eventually leads to excessive extracellular matrix deposition (13, 15) and compound polymorphisms in CCL5 and CXCL8 seem to increase the risk of SSc (34).
Furthermore, in the course of SSc, abnormal expression of some chemokines could also influence the pathologic angiogenesis and vasculogenesis (35). Recent data suggest that upregulation of some chemokines such as CXCL9, CXCL10, CXCL16, and downregulation of others as CXCL5 may influence vasculopathy in SSc (19, 36) as well as the increase or downregulation of their receptor (19).
Angiogenic Chemokines
CCL 2 (MCP-1)
Among CC chemokines that are angiogenic, monocyte chemoattractant protein-1 (MCP-1/CCL2 and MIP-1a/CCL) that binds to CCR2, is overexpressed in SSc skin and serum of SSc patients (21, 37, 38). CCL2 is produced by macrophages, fibroblasts, endothelial cells and other cells, and attracts and activates monocytes and T cells, inducing T-helper 2 cell polarization (39) and stimulates collagen production by fibroblasts via specific receptors and endogenous upregulation of TGFβ expression (21, 22, 40, 41). In fact, circulating levels of CCL2 correlate in SSc with the extent of skin and pulmonary fibrosis, likely playing a role in tissue fibrosis (21, 42–43–44). SSc fibroblasts express CCR2 (45), and mice that do not express CCL2 receptor CCR2 are protected from fluorescein isothiocyanate-induced and bleomycin-induced lung fibrosis (46, 47). It has been reported also that cultured SSc dermal fibroblasts display augmented expressions of CCL2 mRNA and protein (22, 48) compared to healthy controls. Furthermore, stimulation with platelet-derived growth factor (PDGF) significantly enhanced CCL2 expression in dermal SSc fibroblasts (21, 22) as well as exogenously administered CCL2 stimulated auto-induction of CCL2 mRNA by scleroderma, but not normal, dermal fibroblasts (22). Anti-CCL2 antibody therapy reduces mononuclear phagocyte accumulation in bleomycin-challenged mice (49). Further work suggests a role for CCL2 in matrix turnover by upregulating matrix-degrading enzymes and their inhibitors such as tissue inhibitor of matrix metalloproteinase-1 (TIMP-1) (50).
Hasegawa et al (51) reported that serum CCL2 levels declined with disease progression, along with improvement of skin thickness assessed by modified Rodnan Skin Score (mRSS). They confirmed the correlation between CCL2 and lung fibrosis: moreover, they found that when the progression of interstitial fibrosis was evaluated by variations of % of vital capacity (VC), the variations of %VC during the 3 years were inversely associated with the changes of CCL 2 circulating levels (51).
Luzina et al (52) reported that expression of CCL2 mRNA was most augmented among 4507 genes in bronchoalveolar lavage (BAL) cells from SSc inflammatory lungs as well as protein levels of CCL2 that was also increased and associated with the presence of interstitial lung disease (ILD), correlating negatively with lung function parameters and positively with computed tomography scores (52).
Interestingly, CCL2 inhibits alveolar epithelial cell-derived prostaglandin E2 (PGE2) synthesis and a reduction of PGE2 levels enhanced proliferation (53). Thus, an important profibrotic mechanism of MCP-1 interactions with its main receptor, CCR2, is to limit prostaglandin E2 production in alveolar epithelial cells after injury, thus promoting fibrosis (53).
High CCL2 levels have also been described in early phases of SSc and in patients with higher frequencies of cardiac, vascular or kidney involvement (13, 37, 45, 54), and an up-regulation of CCL2 production has been demonstrated in animal models of crescentic nephritis (55). In a rat model of monocrotaline-induced pulmonary hypertension, gene transfer of a dominant negative inhibitor of the CCL2 gene inhibited the progression of CCL2-induced pulmonary hypertension and mononuclear cell infiltration into the lung (56). This suggests that early inhibition of MCP-1 signaling may prevent the recruitment of monocytes/macrophages into perivascular tissues, preventing the immune-mediated development of pulmonary hypertension in this model (56).
In the early phase of SSc, in animal models, all CCL5, CCL2 CCL3 molecules are up-regulated (57), but while MCP1/CCL2 and MIP1α/CCL3 remained increased during the time, RANTES/CCL5, after an initial marked peak, rapidly decreased, suggesting a more relevant role for MCP1/CCL2 and MIP1α/CCL3 (57).
It was thus suggested that CCL2 levels could be a useful biomarker to follow-up skin and lung fibrosis and to assess therapeutic efficacy.
CCL5 (RANTES)
Levels of “regulated upon activation, normal T cell expressed and secreted” (RANTES/CCL5) and macrophage inflammatory protein-1α (MIP-1 α/CCL3) are increased in serum and BAL fluids from SSc patients (38, 58). Increased expression of RANTES and MIP-1α precedes the development of dermal and pulmonary fibrosis in a murine model of scleroderma (59). Expression of abundant CCL5 mRNA has been demonstrated in situ in the skin of SSc patients, but not in the skin of healthy control (60) and also in lung samples from patients with severe pulmonary arterial hypertension (PAH), in comparison with controls (61). Endothelial cells seem to be the major source of CCL5 within the pulmonary artery walls of SSc patients and associated with CD45+ inflammatory cell infiltrates (61).
CCL5 levels may be a potential useful biomarker of PAH in SSc patients.
CCL24 (Eotaxin 2)
The chemokine CCL24 is a potent chemoattractant for inflammatory cells, predominantly eosinophils, basophils and T helper type 2 (Th2) lymphocytes through its receptor CCR3. It is produced in various cell types including: activated T lymphocytes, macrophages, endothelial cell, epithelial cells and dermal fibroblasts (62–63–64).
Additionally, CCL24 was shown to stimulate human lung fibroblast proliferation and collagen synthesis (65). Knock out mice to CCL24 revealed significantly reduced inflammatory cell infiltration into BAL fluid in OVA-induced model of pulmonary inflammation (66). Recently, it was discovered that high CCL24 sera levels were described in limited and diffused phases of SSc and that SSc skin showed significant overexpression of both CCL24 and its cognate receptor, CCR3 (67). A blockade of CCL24, using a specific monoclonal antibody, was described in the past to result in anti-inflammatory and anti-atherosclerotic features, that are significantly relevant in the SSc pathogenesis (68, 69). Recently, it was revealed that treatment with anti-CCL24 monoclonal antibody in bleomycin-induced scleroderma mice model, exhibited reduced symptoms of the disease, including dermal thickness, immune cell infiltration to the lung and collagen deposition in the skin (70).
Over-expression of CCL24 in the skin and sera of SSc patients may suggest involvement of CCL24 in the pathogenesis of SSc.
CXCL 16 (SRPSDX)
CXCL16 is a CXC chemokine expressed on a number of different cell types, including leukocytes, endothelial cells (ECs) and keratinocytes that bind to CXCR6 (71). CXCL16 is present in membrane-bound receptors CXCR6 and CXCR5 that facilitate firm adhesion of emigrating leukocytes, and soluble forms that mediate chemotactic properties (72, 73). It is well known that this angiogenic chemokine is overexpressed in SSc patients, both in skin and serum and its receptor CXCR6 is overexpressed on ECs of the skin (19). Elevation of serum CXCL16 levels were found in SSc patients and associated with greater extent of skin fibrosis but not with the severity of pulmonary fibrosis (71). Rabquer et al (19), analyzing skin biopsies and serum samples of SSc patients, found that CXCL16 was significantly upregulated in early SSc or with PAH. CXCL16 was actively expressed in bronchial epithelial cells and lung fibroblasts, and increased concentrations can be found in BAL fluids from patients with interstitial lung disease (ILD) (74).
CXCL16 seems to play an important role in the development of skin sclerosis in patients with SSc but it is not useful as a clinical marker of pulmonary fibrosis.
CXCL5 (ENA-78)
CXCL5 is a member of CXC chemokines with glutamic acid-leucine-arginine (ELR) motif and it is also known as epithelial-derived neutrophil activating peptide 78 or ENA-78, produced by epithelial cells following stimulation by IL1 or TNFα (75). It is also expressed in eosinophil, which supports the notion that eosinophils are not mere targets for chemokines, but also have immunoregulatory properties (76). CXCL5 has been found elevated in BAL fluid from patients with idiopathic pulmonary fibrosis (IPF) (77). In SSc, Ichimura et al (36) reported that serum CXCL5 levels were significantly decreased in diffuse SSc (dcSSc) and limited SSc (lcSSc) patients compared with healthy controls, suggesting a negligible role for CXCL5 in the fibrotic process underlying SSc. Furthermore, serum CXCL5 levels were reduced in early SSc (<1 year), characterized by abnormal vascular activation, correlated with disease duration and decreased during the progression of the disease, amplifying the risk for the development of digital ulcers. This suggested that CXCL5 is associated with disease onset and with vascular aspects of disease progression (36).
Serum CXCL5 levels may serve as an inverse marker for the severity of SSc vasculopathy.
CCL19 (MIP-3β/ELC)
CCL19 is upregulated in SSc and its expression correlates strongly with markers of vascular inflammation (angiopoietin 2, Von Willebrand factor [vWF] and junctional adhesion protein 2 [JAM2]) and with macrophage markers: CD163, Siglec-1 and CCL2 The immunofluorescence and in vitro data identify macrophages as an important source of CCL19 and as macrophages are present in the skin under non-inflammatory conditions, resident as well as recruited macrophages might be an important source of CCL19. A strong correlation of CCL19 with vascular inflammation and macrophage recruitment has been demonstrated (78).
Furthermore, CCL19 is a ligand for CCR7, which is expressed on many antigen-presenting cells and is important for their homing to lymph nodes from peripheral tissues during early- and late-stage inflammation (79, 80). This suggests that CCL19 plays a role in the recruitment of cells to the skin during SSc rather than emigration of antigen-presenting cells after activation. This is supported by the correlation of CCL19 with vascular inflammation expressed by infiltrating immune cells in SSc skin (21–22–23, 78). Overall, this study suggests that CCL19 plays a role in cutaneous inflammation in dcSSc (78).
CCL19 seems to be the only chemokine upregulated in SSc skin, associates with vascular injury and may be a sensitive marker for macrophage activity.
CXCL8 (IL-8)
CXCL8 (IL-8) is a chemokine often produced by macrophages and epithelial cells by mediating the inflammatory response and an angiogenic pathway (81). CXCL8 is a neutrophil chemoattractant factor and a potent proangiogenic chemokine overexpressed both in SSc serum and skin (82–83–84) that binds two receptors CXCR1 and CXCR2 even if its angiogenic properties are mediated primarily by CXCR2, and two polymorphisms of its gene have been associated with scleroderma (85). To date, CXCR2 has been shown to be upregulated on SSc dermal fibroblasts (86, 87), though no studies have investigated its expression on SSc ECs. Expression levels of CXCL8 are increased in SSc serum and skin, particularly in early SSc patients with disease of less than 1 year duration (84). A recent study demonstrated that plasma CXCL8 levels are elevated in anti-topoisomerase I-positive SSc (88). CXCL8 is also increased in BAL fluid from SSc patients, suggesting a critical role for CXCL8 in SSc-ILD (58, 89). Compound polymorphisms in CCL5 and CXCL8 appear to increase the risk of SSc (34). Finally, Hasegawa et al (90) reported that baseline serum CXCL8 levels seem significantly associated with severity of physical dysfunction at baseline and in subsequent years in SSc patients.
Literature data about the potential role of IL-8 in fibrosis are mainly centered on IPF showing greater IL-8 levels in lung (91). When IL-8 is depleted from IPF tissue specimens, tissue-derived angiogenic activity is markedly reduced (91). Studies in animal models of pulmonary fibrosis have shown the importance of chemokines in promoting angiogenesis, which is necessary for the development of pulmonary fibrosis (92).
Serum CXCL8 levels may be useful for predicting the decline in physical function in patients with progressive SSc.
The Angiostatic Chemokines
CXCL4 (platelet factor 4; PF-4)
CXCL4, also known as platelet factor-4, is a 70-amino acid, lysine-rich, 7.8-kDa protein that was first identified as a product of megakaryocytes and is chemotactic for neutrophils, monocytes and fibroblasts, and may have important roles in inflammation and wound repair (93).
CXCL4 is generally considered one of the most potent anti-angiogenic chemokines, influencing angiogenesis through an integrin-dependent mechanism (94). Romagnani et al (95) reported that increased circulating levels of CXCL4, found in the serum of SSc patients, showed anti-angiogenic activity, and the disease was characterized by rarefaction of vessels, despite the presence of multiple proangiogenic factors.
Increased levels of CXCL4 were also found in patients with Raynaud's phenomenon, most of whom did not progress to SSc. This finding could support the hypothesis that CXCL4 may sensitize several cellular components, including fibroblasts but the presence of CXCL4 alone is not sufficient, but play an additional role in the initiation and maintenance of cellular activation.
In addition, CXCL4 induces the expression of thrombospondin 1 and attenuates the effects of vascular endothelial growth factor A (VEFG-A), one of the major proangiogenic regulators that largely mediates angiogenesis by primarily binding vascular endothelial growth factor receptor 2 (VEGFR2) and that is markedly increased in serum and different cell types both in the epidermis and dermis of patients with SSc (96–97–98).
These observations might explain the absence of ‘appropriate’ neovascularization in SSc, despite the presence of high levels of VEGF and VEGFR2 which is actually upregulated on SSc ECs (99). Interestingly, Manetti et al (100) reported further data to explain this paradox. They demonstrated that endogenous antiangiogenic VEGF165b splice variant is selectively upregulated in SSc skin and circulation and provided evidence that profibrotic TGFβ 1 and SRp55 splicing factor may contribute to the switch from proangiogenic to antiangiogenic VEGF isoforms and that constitutive overexpression of antiangiogenic VEGF165b leads to impaired VEGFR-2 downstream signaling and capillary morphogenesis in SSc microvascular endothelial cells (MVECs) (100).
CXCL4 is also a profibrotic chemokine, inhibiting the expression of the antifibrotic cytokine interferon-γ (from type 1 helper T cells), up-regulating profibrotic cytokines interleukin-4 and interleukin-13 (from type 2 helper T cells) and stimulating the proliferation of regulatory T cells with impaired suppressive function (101).
A recent multicenter study used proteome-wide analysis to demonstrate that CXCL4 was the predominant protein secreted by plasmacytoid dendritic cells in SSc skin and plasma (99). Furthermore, levels of CXCL4 seem to correlate with the development of skin and pulmonary fibrosis (PF) and PAH (99), but not with extent and progression of PF as measured by change in diffusing capacity of the lungs for carbon monoxide (DLCO) (102). CXCL4 levels have also been found elevated in the BAL fluid of patients with SSc-ILD (103).
The identification of CXCL4 as a marker for fibrosis and PAH may be helpful in early diagnosis and risk assessment of disease progression and an important guide in patients who require a more aggressive treatment since the earliest phase of disease.
CXCL 10 (IP-10)
Interferon-γ-inducible protein 10 (CXCL10, also called IP-10) is a 10 kDa protein and it is functionally categorized as an ‘inflammatory’ chemokine. It was initially identified as a chemokine that is induced by IFN-γ and, under the influence of cytokines, is secreted by various cell type, such as neutrophils, keratinocyte, endothelial cells, fibroblasts, monocytes, mesenchymal cells, dendritic cells, and astrocytes (104). CXCL10 lacking its ELR motif, suppresses neovascularization and functions as an ‘angiostatic’ chemokine (105) and upon binding the receptor CXCR3, it regulates immune responses by promoting recruitment of leukocytes, such as T cells, eosinophils, monocytes, and natural killer (NK) cells (106). In bleomycin-induced pulmonary fibrosis, levels of the CXCL10 are decreased. This chemokine inhibits fibroblast proliferation and deposition of extracellular matrix by regulating angiogenesis (107).
The presence of high CXCL10 levels in peripheral liquids could thus stand as a marker of the host's immune response, especially T helper (Th) 1-orientated T-cells (108).
A recent study proposed a composite chemokine score of plasma levels of CXCL10 (IFN-γ-inducible protein 10; IP-10) and CXCL11 (IFN-γ-inducible T-cell-a chemoattractant; I-TAC) was elevated in SSc patients and correlated with the Medsger severity index, particularly with the severity of lung, skin and muscle involvement (109).
It has been observed that increased CXCL10 levels were associated with cardiac involvement, signs of PAH, elevated ESR, and although only present in few patients, with digital loss and renal crises (44). Moreover, CXCL10 is specifically increased in SSc complicated by ILD (110), and could serve as a disease activity marker (51).
IFN-γ-inducible chemokine score that may be a promising stable serologic marker of a more severe form of SSc and may be useful for risk stratification of patients.
CXCL 9 (MIG)
Chemokine (C-X-C motif) ligand 9 (CXCL9) is a small cytokine belonging to the CXC chemokine family that is also known as monokine, induced by gamma interferon (MIG). CXCL9 is a T-cell chemoattractant, which is induced by IFN-γ and has antiangiogenic effects binding CXCR3 receptor. CXCL9 was found significantly elevated in SSc serum compared to normal controls, regardless the subset of disease. (19, 51, 111) even if their levels are not likely as useful as CCL2 for evaluating disease activity. Until today no relationship between CXL9 levels, variation and change in clinical parameters in SSc have been reported.
CXCL9 has antiangiogenic effects binding CXCR3 receptors, but further studies are needed to elucidate its role in SSc pathogenesis.
Chemokines and Cytokines Cross Talk in Systemic Sclerosis
In SSc, abnormal immunological regulation is considered to be a key stimulus in promoting vascular abnormalities, starting with the earliest phase of SSc (112). This applies to CD3+ T lymphocytes, with CD4+T cells predominating over CD8+T cells.
Mononuclear cell infiltrate is predominantly perivascular suggesting that the microvascular endothelium is one of the major targets of the immune reaction and a trigger for subsequent inflammatory changes and development of fibrosis (113).
B cells, NK cells and increased numbers of mast cells, particularly degranulated, have been detected in the skin of SSc patients in the early stages (114). Th cells differentiate from naïve cells into various subsets (Th1, Th2, Th17 and Th22) associated with a set of inducer and effector cytokines. Previously, it was thought that this differentiation was an irreversible event but recent evidence suggests that differentiated Th cells retain the flexibility to transform from one lineage to another (115–116–117). The pattern of cytokine production of skin-infiltrating T cells from SSc patients demonstrated a high concentration of interleukin-4 (IL-4) and high IL-4 mRNA expression in skin perivascular mononuclear cell infiltrates by in situ hybridization (118). IL-4 drives the production of IgE as Th2-cell-derived IL-9 contributes to the regulation of IgE production and to mast cell growth (119). T cells from peripheral blood, skin, BAL fluid, and several other tissues obtained from SSc patients have undergone oligoclonal expansion driven by one or more unidentified specific antigens (8).
In 2004, Fujii et al (111) showed that Th2 chemokines (TARC/CCL17 and MDC/CCL22) and serum CXCL10 levels were elevated in patients with diffuse and limited SSc. A few years later, Rabquer et al (19) examined CXCL9, CXCL10, and CXCL16 levels in SSc serum and skin, and their receptors in SSc skin and on ECs derived from the skin of patients with SSc. They found that while the expression of CXCL9 and CXCL10 were elevated in SSc sera, the expression of CXCR3 was reduced on SSc dermal ECs. In contrast, CXCL16 and CXCR6 were elevated in SSc sera and in SSc dermal ECs, respectively (19). These findings suggest that angiogenic chemokine receptor expression is important to promote angiogenesis in SSc since both pro- and anti-angiogenic chemokines are elevated systemically in SSc, and for this reason it is important to determine the expression of both angiogenic mediators and their receptors.
A longitudinal study demonstrated high serum levels of CXCL10 (Th1) and CCL2 (Th2) chemokines in newly diagnosed SSc. High values of CXCL10 were associated with a more severe clinical phenotype (lung and kidney involvement) and appeared to decline during the follow-up, while CCL2 levels remained unmodified, suggesting that the disease progresses from the early Th1 inflammatory condition to the advanced Th2-like stage (120).
CXCL8 and CCL2 are the most critical chemokines for the recruitment of neutrophils and monocytes, respectively. Further, the transition from neutrophil to monocyte accumulation during inflammation might be linked to a shift in chemokine synthesis from CXCL8 to CCL2, probably via IL-6 signaling (121). Therefore, increased CXCL8 production may trigger initial inflammation while CCL2 may contribute to the subsequent chronic inflammation and fibrosis in SSc. Thus, CXCL8 levels, rather than CCL2 levels, can likely predict the progression of SSc, despite the finding that serum levels of CXCL8 and CCL2 were associated with each other (90).
Conclusions
Increased expression of several chemokines as CXCL4, CXCL8, CXCL5, CCL5, CXCL9, CCL24, CXCL10, CXCL12, CXCL16, CCL2, CCL2, CCL19, suggests a complex mechanism by which immune cells are recruited into SSc skin. Many of these chemokines have redundant roles, possibly to ensure recruitment of specific cell types. Several studies have shown a synergistic effect of combinations of these chemokines in cell recruitment, emphasizing the importance of understanding global chemokine expressions (122, 123). In fact, chemokines act through numerous pathways and mechanisms, including attraction of inflammatory cells; direct action on target cells, such as fibroblasts and endotheliocytes; stimulation of production and activation of TGFβ, including autocrine mechanism; and imbalance intracellular signaling activated by these molecules. The effects of chemokines on endothelial cells allow angiogenesis, a process that is necessary for the influx of fibroblasts and inflammatory cells as well as epithelial cell alterations that are required for the development of fibrosis (43).
Furthermore, because chemokines can be detected easier in peripheral blood compared with cytokines or growth factors, their utility as biomarkers of SSc have been investigated. It is interesting to note that the levels of inflammatory chemokines are increased in SSc independently of the disease phenotypes and clinimetric measures. This suggests that, while chemokines are clearly associated with the pathogenesis of SSc, other factors may be critical for determining the level of disease activity and severity even if some of them could be useful as predictive biomarkers, in particular CXCL4, CXL8, CXCL5 and CCL2 (124).
In conclusion, the clinical utility of chemokines as biomarkers is not yet clarified particularly because some factors may influence the levels (disease phase, treatment, etc.). Serial measurements or longitudinal study of serum chemokines levels could be useful for the evaluation of disease activity, therapeutic effects of skin sclerosis or interstitial pneumonia and risk stratification of SSc patients. An integrative and effective therapeutic agent able to interfere with complex chemokine networks, described above, so as to attenuate perivascular inflammation, dysregulated angiogenesis and skin and internal organ fibrosis, is the most ambitious goal for the scientific research of the future.
Footnotes
Financial support: No grants or funding have been received for this study.
Conflict of interest: None of the authors has financial interest related to this study to disclose.