
Abstract
Purpose
The gut is in direct contact with BDE-99 (2,2′,4,4′,5-pentabromodiphenyl ether), one of the most abundant PBDE congeners in the environment and in human tissues. The objective of the present study was to investigate the effects of BDE-99 on colorectal cancer (CRC) cells.
Methods
The effects of BDE-99 on cell proliferation were measured by CCK-8 assay in the CRC cell line HCT-116. Wound healing and transwell migration/invasion assays were used to test the migration and invasion of CRC cells. Factors related to epithelial-to-mesenchymal transition (EMT) were measured by real-time PCR and Western blot analysis for mRNA and protein levels, respectively.
Results
BDE-99 was found to increase migration and invasion and trigger EMT in HCT-116 cells; EMT was characterized by cells acquiring mesenchymal spindle-like morphology and by increased expression of N-cadherin with a concomitant decrease in E-cadherin. BDE-99 treatment also increased the protein and mRNA levels of the transcription factor Snail, but not Slug, Twist, and ZEB1. Knockdown of Snail by siRNA significantly attenuated BDE-99-induced EMT in HCT-116 cells, suggesting that Snail plays a crucial role in BDE-99-induced EMT. The PI3K/Akt inhibitor LY294002 completely blocked BDE-99-induced Snail and invasion of HCT-116 cells.
Conclusions
Our results revealed that BDE-99 can trigger the EMT of colon cancer cells via the PI3K/AKT/Snail signaling pathway. This study provides new insight into the tumorigenesis and metastasis of CRC stimulated by BDE-99 and possibly other PBDE congeners.
Introduction
As flame retardants, polybrominated diphenyl ethers (PBDEs) have been widely used in various products and processes such as electronics, furnishings, polyurethane foams, and plastics (1). PBDEs are highly resistant to degradation and therefore can enter the food chain and threaten human health (2). They are found in various environmental media (3, 4) and are considered as health hazards (5). With regard to PBDE accumulation in the human body, BDE-99 (2,2′,4,4′,5-pentabromodiphenyl ether) has shown the highest concentrations and is one of the most abundant PBDE congeners in the environment and in human tissues (6).
Although BDE-99 can be easily accumulated in human tissues, few experimental studies on BDE-99 toxicity have been reported, while mechanistic information is also lacking. BDE-99 has been found to elicit various adverse effects including neurobehavioral toxicity (7), developmental and reproductive toxicity (8), and endocrine disruption (9). Recent studies suggested that acute oral BDE-99 exposure can cause renal and liver impairment, with oxidative damage being a potential mechanism for nephrotoxicity and hepatotoxicity (10). Exposure to low doses of BDE-99 has been suggested to cause biochemical and hormonal alterations (10). Although the gut is in direct contact with PBDEs consumed orally, there are very limited data regarding the effects of BDE-99 on the development and progression of colorectal diseases.
Colorectal cancer (CRC) is the third most common form of cancer and the second leading cause of cancer-related death in the Western world (11). Recent studies suggested that more than 80% of CRC cases and deaths are attributable to diet and environmental factors (12). Most studies of the carcinogenic characteristics of PBDEs were focused on breast cancer. The risk of developing breast cancer has been suggested to be associated with adipose concentrations of PBDE among women undergoing surgical breast biopsies (13). Barber et al (14) revealed that very low PBDE doses (10−12 to 10−9 M) can trigger the formation of micronuclei in MCF-7 breast cancer cells. Further studies also suggested that PBDEs can promote the proliferation of MCF-7 cells at very low doses (15, 16). However, there is no study concerning the effects of BDE-99 on tumorigenesis and progression of CRC.
Therefore, the present study was designed to investigate the effects of BDE-99 on the progression of colon cancer. We found that BDE-99 can promote epithelial to mesenchymal transition (EMT) in HCT-116 CRC cells via the transcription factor Snail. To our knowledge, this is the first study illustrating the effects of BDE-99 on CRC invasion.
Materials and Methods
Reagents
All chemicals were of reagent grade or better and were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA) unless otherwise noted. BDE-99 was dissolved in dimethyl sulfoxide (DMSO) to prepare a 10 mM stock solution, and stored at −20°C. Primary antibodies against E-cadherin (E-Cad), Zona occludin-1 (ZO-1), N-cadherin (N-Cad), fibronectin (FN), vimentin (Vim), Snail, Slug, ZEB, Twist, p-AKT (Ser473), AKT, and β-actin were purchased from Cell Signaling Technology (Beverly, MA, USA). Horseradish peroxidase-conjugated secondary antibody was from Santa Cruz Biotechnology (Santa Cruz, CA, USA). All compounds were solubilized in DMSO. The PrimeScript® RT reagent kit and SYBR® Premix Ex Taq™ were products of Ta KaRa (Takara Shuzo, Japan). E.Z.N.A.® HP Total RNA kit was purchased from Omega Bio-Tek (Doraville, GA, USA). Medium containing 0.5% DMSO was used as the control.
Cell line and cell culture
The HCT-116 cell line (passage 12~15) conserved in our own laboratory was grown in Dulbecco's Modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 IU/mL penicillin and 100 μg streptomycin. The cells were incubated at 37°C in a 5% CO2 atmosphere. Twenty-four hours before the experiments, the medium was removed and replaced with DMEM without phenol red supplemented with 5% dextran-coated, charcoal-treated FBS (DCC-FBS) to exclude estrogenic effects caused by the medium. Cells were then plated in the same medium and allowed to attach overnight.
Cell viability assays
The proliferation and viability of cells were assessed by means of the Cell-Counting Kit-8 (CCK-8, Dojindo Laboratories, Kumamoto, Japan) assay according to the manufacturer's instructions. Briefly, HCT-116 cells were seeded in 96-well plates at a cell density of 1 × 10 4 per well. After treatment with increasing concentrations of BDE-99 for 48 hours, 10 μL of CCK-8 solution was added to each well. The plates were incubated for another 2 hours, and the absorbance was measured at 450 nm using a microplate reader. Experiments were repeated 3 times.
Wound healing and transwell migration/invasion assay
For the in vitro wound healing assay, confluent monolayers of HCT-116 cells were scratched, and the migration distance of the cells into the scratched area was measured in 5 randomly chosen fields. Migration and invasion assays were performed in Boyden chambers according to previous studies (17, 18). Briefly, polycarbonate filters (8-μm pore size; Corning Costar, Rochester, NY, USA) pre-coated with Matrigel matrix (BD Bio-sciences, Franklin Lakes, NJ, USA) were used for the invasion assay and uncoated filters were used for the migration assay. Cells (1 × 10 5 ) in 300 μL medium (containing 0.1% FBS) with or without bisphenol A (BPA) were seeded in the upper chamber. Then 600 μL medium with 10% FBS was added to the lower chamber and served as a chemotactic agent. After 48 hours' incubation, the cells that had migrated and adhered to the lower chamber were fixed in 4% paraformaldehyde for 20 minutes, stained with hematoxylin, and counted under an upright microscope (5 fields per chamber). To quantify invasion, cells in the upper chamber were fixed in 4% paraformaldehyde for 20 minutes, then the Matrigel was mechanically removed from the filter with a cotton swab. Cells adhering to the bottom of the filter were stained with hematoxylin and counted under an upright microscope (5 fields per chamber). Each migration and invasion assay was repeated in 3 independent experiments.
Quantitative real-time PCR
Quantitative real-time PCR was performed as previously described (19). Briefly, HCT-116 cells (1 × 10 6 ) were plated onto 6-well plates. After exposure to BDE-99 for 24 hours, cells were washed twice with ice-cold phosphate-buffered saline. Total mRNA was extracted with TRIZOL reagent. The first strand of cDNA was generated from 2 μg total RNA using oligo-dT primer and SuperScript II Reverse Transcriptase (GIB-CO BRL, Rockville, MD, USA). Quantitative real-time PCR was run on an iCycler (Bio-Rad, Hercules, CA, USA) using validated primers and SYBR Premix Ex Taq II (TaKaRa) for detection. The cycle number when the fluorescence first reached a preset threshold (Ct) was used to quantify the initial concentration of individual templates for expression of mRNA of the genes of interest. The primer pairs were as follows: GAPDH, forward 5′-GCA CCG TCA AGG CTG AGA AC-3′ and reverse 5′-TGG TGA AGA CGC CAG TGG A-3′; E-Cad, forward 5′-TAC ACT GCC CAG GAG CCA GA-3′ and reverse 5′-TGG CAC CAG TGT CCG GATTA-3′; Vim, forward 5′-TGA GTA CCG GAG ACA GGT GCA G-3′ and reverse 5′-TAG CAG CTT CAA CGG CAA AGT TC-3′; Snail, forward 5′-GAC CAC TAT GCC GCG CTC TT-3′ and reverse 5′-TCG CTG TAG TTA GGC TTC CGA TT-3′; ZEB1, forward 5′-TAC AGA ACC CAA CTT GAA CGT CAC A-3′ and reverse 5′-GAT TAC ACC CAG ACT GCG TCA CA-3′; Twist, forward 5′-GGA GTC CGC AGT CTT ACG AG-3′ and reverse 5′- TCT GGA GGA CCT GGT AGA GG-3′; Slug, forward 5′-TTC GGA CCC ACA CAT TAC CT-3′ and reverse 5′-GCA GTG AGG GCA AGA AAA AG-3′. Transcripts of the housekeeping gene GAPDH in the same incubations were used for internal normalization.
Western blot analysis
Western blot analysis was performed as previously described (19). Briefly, cells were treated with BDE-99 before being lysed in cell lysis buffer, then lysates were cleared by centrifugation and denatured by boiling in Laemmli buffer. Aliquots of protein were separated on 10% sodium dodecyl sulfate (SDS)–polyacrylamide gels and electrophoretically transferred to nitrocellulose membranes. Following blocking with 5% nonfat milk at room temperature for 2 hours, membranes were incubated with the primary antibody at 1:1000 dilution overnight at 4°C and then incubated with a horseradish peroxidase-conjugated secondary antibody at 1:1000 dilution for 2 hours at room temperature. The expression of EMT-related markers was detected with the Western Lightning chemiluminescence detection reagent (Perkin-Elmer Life Sciences, Wellesley, MA, USA).
RNA interference
HCT-116 cells were plated on 6-well plates (2 × 10 5 cells per well) and cultured for 24 hours. They were then transfected with 2 μg plasmid vector or 100 pmol siRNA oligomer mixed with Lipofectamine 2000 reagent (Invitrogen, Carlsbad, California, USA) in serum-reduced medium according to the manufacturer's instructions. The target sequence for Snail siRNAs was 5′-UGCAGUUGAAGAUCUUCCGCGACUG-3′. Both si-Snail and control siRNAs (sequence 5′-CAGCUUUGGCUGAGCGUAU-3′) were obtained from Ambion (Grand Island, NY, USA).
Statistical analysis
All values were reported as means ± SD of 3 independent experiments unless otherwise specified. Data were analyzed by 2-tailed unpaired Student's t-test for comparisons between 2 groups and by 1-way ANOVA followed by the Bonferroni test for multiple comparisons. For the statistical analyses we used SPSS 13.0 for Windows. A p value <0.05 was considered statistically significant.
Results
Effects of BDE-99 on proliferation and invasion of HCT-116 cells
To determine the cytotoxicity of BPA in colon cancer cells, HCT-116 cells were treated with concentrations of BDE-99 ranging from 10−10 to 10 4 M for 24, 48, and 72 hours, and then cell viability was assessed by the CCK-8 assay. The results showed that BDE-99 had limited effects on the proliferation of HCT-116 cells (Fig. 1A). The effects of BDE-99 on HCT-116 cells were further investigated by use of wound healing and transwell migration/invasion assays. Interestingly, treatment with 10−6 M BDE-99 for either 48 or 72 hours significantly increased wound closure compared with the control group (p<0.05) (Fig. 1B). Furthermore, exposure to 10−6 M BDE-99 resulted in significantly more migration and invasion by cells (Fig. 1C). Compared with the untreated cells, the numbers of migrated and invaded cells treated with BDE-99 were significantly greater (p<0.05) than those of control cells at 48 hours and 72 hours. These results suggested that BDE-99 has limited effects on proliferation but can significantly promote the invasion of HCT-116 cells.
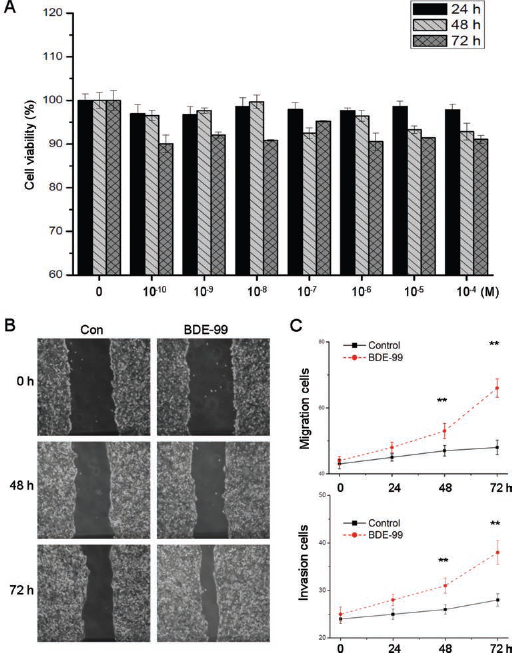
Effect of BDE-99 on proliferation and invasion of HCT-116 cells. (A) Cells were treated with various concentrations (10−10 to 10−4 M) of BDE-99 for 24, 48, and 72 hours and then cell viability was assessed by the CCK-8 assay. (B) Representative images of wounds at 0, 48, and 72 hours in the presence or absence of BDE-99 (10−6 M). Confluent monolayers of HCT-116 cells were scraped by a pipette tip to generate wounds and were then cultured in the presence or absence of BDE-99 for the indicated times. (C) HCT-116 cells were allowed to migrate transwell chambers or spread through the matrix gel and into the bottom side of the filter for different times in the presence or absence of BDE-99 (10−6 M). The migrated or invasive cells were fixed, stained, and photographed. The numbers of migrated and invasive cells were compared with control cells. Data are presented as means ± SD of 3 independent experiments. **p<0.05 compared with control.
BDE-99 induces an EMT phenotype in HCT-116 cells
The increased migration and invasion of HCT-116 cells suggests that BDE-99 might be able to induce the EMT process, in which the epithelial markers E-Cad and ZO-1 were decreased while the mesenchymal markers N-cad and FN were increased (20). Considering the relationship between EMT and cell metastasis, we further measured the effects of BDE-99 on the occurrence of EMT in HCT-116 cells. The morphology of HCT-116 cells treated with or without 10−6 M BDE-99 was observed by phase-contrast microscopy. We found that HCT-116 cells treated with BDE-99 decreased cell-to-cell contacts and the induction of a fibroblast-like state (Fig. 2A). Therefore, we measured the protein expression of EMT-related biomarkers. Western blot analysis showed that this morphological change was associated with downregulation of the epithelial markers E-Cad and ZO-1 and upregulation of the mesenchymal markers Vim, FN and N-Cad (Fig. 2B). qRT-PCR analysis further confirmed the increasing expression of Vim, FN and N-Cad, and the decreasing expression of E-cad and ZO-1 at the mRNA level (Fig. 2C). Collectively, these results indicated that BDE-99 plays an important role in promoting an EMT phenotype in HCT-116 cells.
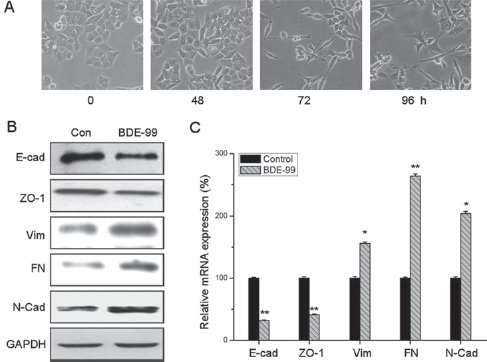
BDE-99 induces an EMT phenotype in HCT-116 cells. (A) Cells were treated with 10−6 M BDE-99 for 0 to 96 hours and then morphological cell changes were studied by phase-contrast microscopy. (B) Western blot analysis showed that the epithelial markers E-Cad and ZO-1 were downregulated while the mesenchymal markers Vim, FN, and N-Cad were upregulated in HCT-116 cells treated with 10−6 M BDE-99 for 96 hours. (C) HCT-116 cells were treated with or without BDE-99 (10−6 M) for 96 hours. The mRNA levels of E-Cad, ZO-1, Vim, FN, and N-Cad were analyzed by qRT-PCR. *p<0.05 compared with control; **p<0.01 compared with control.
Snail plays an essential role in BDE-99-induced EMT
Recent studies indicated that transcription factors such as Snail, ZEB1, Twist and Slug play essential roles in regulating EMT (21), so their roles in BDE-99-induced EMT of HCT-116 cells were investigated. Compared with untreated cells, BDE-99 treatment significantly increased both the protein and mRNA levels of Snail, but did not alter the mRNA and protein levels of Slug, Twist and ZEB1 (Fig. 3A and B).
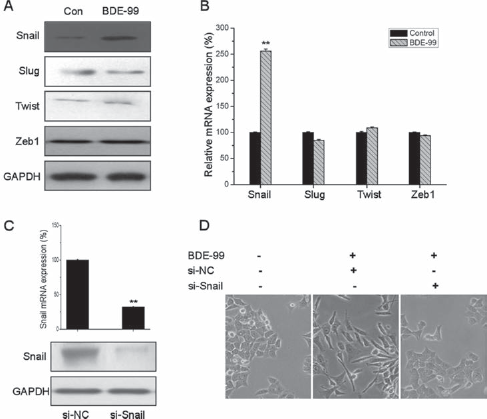
Snail is essential in BDE-99-induced EMT of HCT-116 cells. Cells were treated with 10−6 M BDE-99 for 96 hours, then protein and mRNA expression of the transcription factors Snail, Slug, Twist, and ZEB1 was analyzed by Western blot (A) and qRT-PCR (B), respectively. (C) HCT-116 cells were transfected with Snail-specific si-RNA (si-Snail) or negative control si-RNA (si-NC) for 24 hours, then the mRNA and protein expression of Snail was analyzed by qRT-PCR and Western blot, respectively. (D) HCT-116 cells transfected with si-Snail or si-NC were stimulated with or without BDE-99 (10−6 M) for 96 hours, and morphological changes were examned using phase-contrast microscopy. **p<0.01 compared with control.
To verify the role of Snail in BDE-99-induced EMT of HCT-116 cells, cells were transfected with non-targeting control si-RNA or si-Snail for 24 hours, and then treated with BDE-99 for 96 hours. Both qRT-PCR and Western blot analysis showed that Snail was effectively silenced by siRNA (Fig. 3C). Morphological changes were observed under a phase-contrast microscope. Compared to control cells, spindle-like morphological changes were observed upon BDE-99 addition in control si-RNA-transfected cells but not in si-Snail-transfected cells (Fig. 3D). Generally, these observations demonstrated that Snail is essential for BDE-99-induced EMT in HCT-116 cells.
The PI3K/AKT signaling pathway is involved in BDE-99-induced EMT
Our results indicated that BDE-99 can induce EMT and upregulate Snail in HCT-116 cells. To investigate the underlying molecular mechanisms, inhibitors of PI3K/AKT (LY294002), MAPK (PD98059), and p38 (SB-203580) were used, as previous studies reported that activation of these pathways modulate the EMT progress and Snail expression (22). HCT-116 cells were pretreated with inhibitors for 1 hour before BDE-99 stimulation, then the expression of Snail was detected by Western blot. The results indicated that the AKT inhibitor (LY294002), but not the other inhibitors, completely blocked BDE-99-induced Snail (Fig. 4A), suggesting that the activation of the PI3K/AKT pathway is responsible for BDE-99-induced Snail upregulation. Subsequently, the expression of biomarkers of EMT was measured in HCT-116 cells treated with BDE-99 with or without the AKT inhibitor. The BDE-99-mediated downregulation of E-cad and upregulation of Vim and Snail were clearly attenuated by the AKT inhibitor (Fig. 4B). Furthermore, BDE-99-induced migration (Fig. 4C) and invasion (Fig. 4D) were significantly inhibited when AKT was blocked by the inhibitor LY294002. These results suggested that the AKT signaling pathway was involved in BDE-99-mediated EMT in HCT-116 cells.
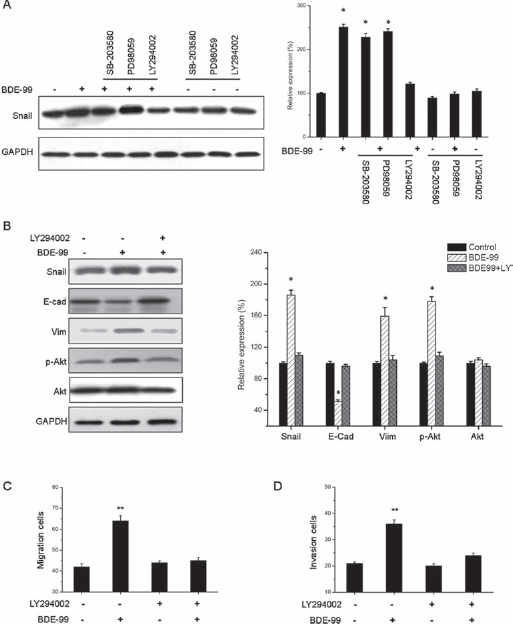
The PI3K/AKT signaling pathway is involved in BDE-99-induced EMT of HCT-116 cells. (A) Cells were pretreated with SB203580 (20 mM), PD98059 (20 mM), and LY294002 (20 mM) for 1 hour, respectively, followed by stimulation with 10−6 M BDE-99 for 48 hours. The expression of Snail was examined by Western blot. (B) HCT-116 cells were pretreated with or without LY294002 (20 mM) for 1 hour, followed by stimulation with or without 10−6 M BDE-99 for 96 hours. The expression of Snail, E-Cad, Vim, p-Akt, and Akt was examined by Western blot. (C) HCT-116 cells were allowed to migrate transwell chambers after pretreatment with or without LY294002 (20 mM) for 1 hour, followed by stimulation with or without 10−6 M BDE-99 for 72 hours. (D) HCT-116 cells were allowed to spread through the matrix gel and into the bottom of the filter after pretreatment with or without LY294002 (20 mM) for 1 hour, followed by stimulation with or without 10−6 M BDE-99 for 72 hours. Bands from 3 independent experiments quantified by densitometry, with results (mean ± SD) normalized to GAPDH expression in each sample. The control value was set as 100%. *p<0.05 compared with control; **p<0.01 compared with control.
Discussion
PBDEs have a similar structure to other persistent organic pollutants such as polychlorinated biphenyls (PCBs). Reports of the International Agency for Research on Cancer suggested that PBDEs are potential carcinogens (23). It is therefore reasonable to hypothesize that PBDEs could induce intracellular and organism toxicity (24). However, there are limited data concerning the effects of BDE-99 related to toxicity and carcinogenicity, although BDE-99 is one of the PBDE congeners with the highest concentrations in the human body (25). This means there is an urgent need to study the toxicity and related mechanisms of BDE-99 in human health.
In the present study, we demonstrated that 10−10 to 10−4 M BDE-99 has limited effects on the proliferation of colorectal HCT-116 cells. This finding is consistent with a previous study showing that the frequency of structural chromosome aberrations was not significantly higher than the control and no sign of cytotoxicity was observed in BDE-99-treated Allium cepa (26). The study of Slotkin et al also showed that growth of PC12 cells was unimpaired by BDE-99 at 50 μM (27). However, our data contradicted the data of Souza et al (28), which showed that BDE-99 can cause death of the hepatoblastoma cell line HepG2 by apoptosis at higher concentrations (greater than 10−5 M). The difference between our study and theirs may be due to the different cell lines used.
Interestingly, we found that 10−6 M BDE-99 can significantly promote the migration and invasion of HCT-116 cells. We used phenol–red-free growth medium and DCC-FBS to eliminate the effects of estrogen, and found that treatment with 10−6 M BDE-99 for 72 hours can induce 1.5-fold greater cell migration and 1.7-fold greater cell invasion than observed in the control group. There has been very little research into the effects of PBDEs on the invasion of cancer cells. One proteomic study revealed that 32 of 111 identified proteins in the neonatal cortex treated with BDE-99 for 24 hours were proteins related to the structure motility of cells such as cofilin and β-actin (29). It was also reported that 3 weeks of PBDE-47 exposure significantly decreased the microtubule subunit β-tubulin in the mussel Mytilus edulis (30). Considering that the cytoskeleton and its associated proteins play a major role in cell migration, our results are consistent with those of previous studies and suggest that BDE-99 can promote tumorigenesis and tumor progression by triggering metastasis.
Furthermore, BDE-99 treatment for 96 hours resulted in an EMT phenotype change combined with the upregulation of epithelial markers and downregulation of mesenchymal markers via the Snail transcription factor. As the first step of tumor invasion and metastasis, EMT has been suggested to be significantly correlated with the progression and metastasis of CRC (31). Transcription factors such as Snail are important contributors to the origin and process of EMT (32). Clinical data revealed that upregulation of Snail in CRC tissues was significantly correlated with lymph node metastasis (33). The present study investigated the mRNA and protein levels of Snail, Slug, Twist and ZEB1 to verify whether these transcription factors were involved in BDE-99-induced EMT. The protein and mRNA levels of Snail exhibited rapid upregulation upon BDE-99 stimulation. However, there was no significant change in either mRNA or protein levels of the other 3 transcription factors. When we silenced Snail by means of si-Snail, it turned out that si-Snail significantly blocked the BDE-99-induced morphological changes. There results suggested that Snail plays an essential role in the BDE-99-induced EMT of CRC cells. Other transcription factors such as ZEB2 and E47 were also found to regulate the EMT processes in many cancers (21). Whether ZEB2 and E47 are involved in the regulation of BDE-99-mediated EMT of HCT-116 cells needs further study.
Previous studies reported that multiple signaling transduction pathways, including the PI3K/Akt, MEK/ERK, and p38 MAPK pathways, can modulate the EMT process and Snail expression (34). In the present study, PI3K/AKT was mediated by BDE-99 among these complicated signaling pathways. When blocking the PI3K/AKT signaling pathway by LY294002 inhibitor, we first observed that the overexpression of Snail and the mesenchymal marker Vim and the downregulation of the epithelial marker E-Cad caused by BDE-99 were abolished. Subsequently, LY294002 also inhibited the BDE-99-induced migration and invasion of HCT-116 cells. Our data demonstrated that AKT inhibition abrogated BDE-99-induced EMT and reversed the BDE-99-mediated increase in the invasive capability of HCT-116 cells in vitro. These results indicate that the PI3K/AKT signaling pathway is involved in BDE-99-mediated EMT and invasion of HCT-116 cells.
The current study established that BDE-99 has limited effects on cell viability but can significantly promote invasion and induce EMT phenotype changes in HCT-116 cells. Furthermore, BDE-99 can trigger the EMT of colon cancer cells via the PI3K/AKT/Snail signaling pathway. This study provides new insight into the tumorigenesis and metastasis of CRC stimulated by BDE-99 and possibly other PBDE congeners. The effects of BDE-99 on normal and other cancer cells remains to be determined.
Footnotes
Financial support: No foundation supports.
Conflict of interest: The authors declare no conflict of interest.