
Abstract
The worldwide annual incidence of oral squamous cell carcinoma (OSCC) is over 300,000 cases with a mortality rate of 48%. This cancer type accounts for 90% of all oral cancers, with the highest incidence in men over 50 years of age. A significantly increased risk of developing OSCC exists among smokers and people who consume alcohol daily. OSCC is an aggressive cancer that metastasizes rapidly. Despite the development of new therapies in the treatment of OSCC, no significant increase in 5-year survival has been recorded in the past decades. The latest research suggests focus should be put on examining tumor stroma activation within OSCC, as the stroma may contain cells that can produce signal molecules and a microenvironment crucial for the development of metastases. The aim of this review is to provide an insight into the factors that activate OSCC stroma and hence faciliate neoplastic progression. It is based on the currently available data on the role and interaction between metalloproteinases, cytokines, growth factors, hypoxia factor and extracellular adhesion proteins in the stroma of OSCC and neoplastic cells. Their interplay is additionally presented using the Systems Biology Graphical Notation in order to sublimate the collected knowledge and enable the more efficient recognition of possible new biomarkers in the diagnostics and follow-up of OSCC or in finding new therapeutic targets.
Introduction
Oral cancer is the sixth most common cancer in the world, constituting 3%-5% of all malignant diseases in both sexes. Over 90% of oral cancers are oral squamous cell carcinomas (OSCC) (1-3), with an annual worldwide incidence of over 300,000 cases and a mortality rate of 48% due to its metastatic potential and limited treatment options (4). The incidence of OSCC has significantly increased during the past 2 decades, with a 0.7% and 1.8% increase per year in men and women, respectively (5).
OSCC primarily affects women after menopause (6), smokers, people who consume alcohol and red meat on a daily basis, and people infected with human papillomaviruses (7, 8). There is a significantly higher incidence of OSCC in men than women (9, 10), which may be the result of greater consumption of tobacco and alcohol by men (11). However, both alcohol and tobacco modify the levels of estrogen and testosterone within gender-specific frames (12), which yields possible additional xenohormonal mechanisms involved in the etiology of OSCC. Oral cancer in women occurs almost exclusively after menopause, which suggests that a decrease in estrogen levels may play an important role in the initiation of oral cancer. In female patients with oral cancer, there is a significantly higher prevalence of OSCC compared with control subjects among hysterectomized women and those whose menopause occurred at a significantly younger age (13).
The OSCC microenvironment consists of tumor cells, different stromal cells, and the extracellular matrix (ECM). Based on the latest research, the activated tumor environment in OSCC has a significantly greater effect on metastases than had previously been assumed (14, 15). However, it is still unknown whether the mechanisms in OSCC-activated stromal cells that promote metastases are similar to those identified in the stroma of other cancers (16-18).
Cancer-associated fibroblasts (CAFs) are major promoters of metastasis progression in cancer tissue and the dominant cell population in tumor stroma (19, 20). They may have their origin in different cell types such as mesenchymal stem cells, bone marrow stem cells, endothelial cells and even cancer cells (21). CAFs produce growth factors, cytokines, chemokines and metalloproteinases that stimulate cancer growth (22). Reprogrammed CAFs resembling myofibroblasts that aggregate around neoplastic tissue have been reported to invade normal tissue (23). These spindle-shaped cells in the stroma of OSCC play an active role in producing ECM components. Their increased presence significantly correlates with stage, regional recurrence and distant metastasis (15).
It has been suggested that CAFs induced in stroma mimic a wound-healing microenvironment and stimulate the activation of multiple active molecules that promote angiogenesis, desmoplasia and paracrine activities (19). CAFs initiate vascularization of the tumor edge through the production of vascular endothelial growth factor (VEGF) (24). Together with CAFs, tumor-associated mast cells (TAMCs) also promote angiogenesis and discharge signal molecules such as tumor necrosis factor alpha (TNF-α) and hypoxia-inducing factor 1 alpha (HIF-1α) as well as proinflammatory cytokines such as interleukin 6 (IL-6) (25, 26), which orchestrate cancer growth and influence the dynamics of metastasis.
It is interesting to mention that miR-21, which regulates cell proliferation, apoptosis and epithelial-to-mesenchymal transition during neoplastic progression, is increased in OSCC stroma and its stromal expression has a particularly negative prognostic value (27, 28), showing the potency of this finding for future investigation.
The aim of this review is to provide insight into the complex multifactorial biology of OSCC stroma and the tumor microenvironment based on the currently available data on the role and interaction between stromal metalloproteinases, cytokines, growth factors and extracellular stromal proteins, all related to the mechanisms involved in the biology of OSCC and especially its progression. Additionally, using the systems biology tool “Systems Biology Graphical Notation” (SBGN), a presentation of the interaction between stromal active factors and tumor mechanisms is provided so as to suggest new approaches in the investigation of new biomarkers for diagnostics and therapy targets.
Impact of stromal cytokines on the aggressiveness of OSCC
IL-6 is produced by a variety of cells, primarily monocytes, macrophages and tumor cells, and plays a central role in the regulation of inflammatory and immune responses. It has an important role in cancer progression via effects on proliferation, migration and angiogenesis in several cancers. Oral cancer cells induce CAFs to produce IL-6 (29-31). Additionally, IL-6 produced by stromal cells plays a role in the osteoclast formation induced by OSCC (32).
Interleukin-1 (IL-1) beta mediates the promotion of inflammatory cell migration from blood into inflamed tissue, and regulates the synthesis and decomposition of the ECM (33, 34). It can thus be used as a marker of disease progression (35). IL-1 alpha produced by OSCC cells has a dual effect on CAFs: promotion of CAF proliferation and upregulation of the secretion of cytokines in CAFs such as CCL7, CXCL1 and IL-8 (36).
Studies have suggested that TAMs produce IL-10, which points to a stromal contribution in immunosuppression mechanisms (37).
The influence of gender on the levels of stromal cytokines in the progression of OSCC has been poorly investigated, although cytokine levels are known to be estrogen and testosterone dependent (38, 39). Such information would be of significant value in designing personalized therapy, especially in OSCC, where significant gender differences in incidence and survival have been described.
OSCC stromal growth factors and hypoxia factor
The proliferation and spread of tumor cells depend on their angiogenesis. Tumors that grow quickly lead to hypoxia, which regulates tumor cell survival factors such as HIF-1α and VEGF, leading to even stronger tumor proliferation, angiogenesis and increased radioresistance (40-42). Thus, VEGF interacts with mast cells, especially in the early stages of OSCC carcinogenesis, by promoting epithelial growth and angiogenesis (43, 44). Over 90% of OSCCs have elevated levels of VEGF and HIF-1α (45), both being related to poorer survival (46). These findings support the hypothesis that tumor angiogenesis is closely related to – but not strictly dependent on – the hypoxic conditions of a tumor’s microenvironment (47).
VEGF-A and VEGF-C protein expression is significantly higher in primary OSCC than in normal oral mucosa. Lymph node metastases in patients with oral cancer are closely correlated with positive VEGF and matrix metalloproteinase 9 (MMP-9) expression (25). Besides, VEGF has been shown to be a predictive marker of malignant potential, as increased levels were found in precancerous oral lesions and oral cancer in comparison with controls (48). VEGF acts as an activator of tumor lymphangiogenesis within metastases also via CD163-positive stromal tumor-associated macrophages (44, 49).
Transforming growth factor beta (TGF-β) family members show different expression in OSCC subtypes. CAFs from genetically unstable OSCC, which promote the malignant phenotype by weakening intercellular epithelial adhesion, have shown increased levels of active TGF-β1 and TGF-β2. (50). Additionally, TGF-β1 secreted by stromal fibroblasts induces expression of PDPN (the gene encoding podoplanin) in OSCC cells, which significantly correlates with invadopodia formation and lymph node metastasis (51).
Platelet-derived growth factor receptor beta is exclusively expressed in CAFs of OSCC but not in tumor cells, which gives this biomarker particular significance in diagnostics (52).
Role of stroma in the formation of ECM and desmoplasia
Cancer cells degrade the ECM, thus facilitating migration and metastasis by mechanisms similar to wound healing. As described previously, the OSCC stroma secretes several growth factors and inflammatory mediators that play a crucial role in cancer progression. On the other hand, cancer stroma also expresses altered levels of collagens, elastin, hyaluronic acid, fibronectin, proteoglycans and glycoproteins, leading to the loss of tissue architecture (53).
ECM degradation is caused by proteinases. Matrix metalloproteinases (MMPs) are involved in the initiation, invasion and metastasis of cancer. In oral cancers, MMPs, especially MMP-9, are associated with ECM degradation, allowing the cancer to spread (54).
Disruption in the expression of MMP-1 promotes OSCC progression via increased levels of hyaluronan synthase 2, which is associated with alpha smooth muscle actin (α-SMA)-positive myofibroblasts in the stroma (55). It has been shown that a stromal environment that promotes cancer cell migration is accomplished via stimulated expression of MMPs. This type of expression is the responsibility of thrombospondin-1 produced by stromal cells (56).
In OSCC, retrodifferentiation is demonstrated by the expression of oncofetal fibronectins in the stroma. Both tumor cells and recruited stromal myofibroblasts take part in the formation of the fetal ECM milieu. As shown in one study (57), the oncofetal type III extra-domain (ED-B) fibronectin matrix is produced by recruited stromal cells, which enables the remodeling of the carcinoma invasion front.
A migration-promoting microenvironment in OSCC is also mediated by myofibroblast progression with stromal upregulation of laminin isoforms, which leads to vascular basement membrane reorganization (alpha-3- and gamma-2-chain laminins) (58). Myofibroblasts are α-SMA and vimentin positive, by which they also contribute to OSCC invasion (59). The Gas 6/Axl axis, which takes part in cadherin and vimentin expression and promotes cell invasion, migration and the triggering of NF-κB signaling, has been reported to be active in OSCC and to induce cancer to become apoptosis resistant (37). Additionally, the OSCC stroma seems to have a desmoplastic response that has been described in highly invasive tumors (60).
Versican is overexpressed in different cancer types. It plays a significant role in tumor growth by repressing cell adhesion, stimulating cell proliferation and migration, and regulating angiogenesis. OSCC has strong stromal versican expression, which has been shown to be an independent predictor of unfavorable prognosis (61). Invasion in OSCC is associated with the increased deposition of laminin in stroma, beneath the invading carcinoma cells. Increased expression of laminin-5 has been shown to occur in the areas of direct tumor-stroma interaction (57, 62).
Galectin 1 increases CAF-conditioned-medium-induced tumor cell migration and invasion, possibly by production of monocyte chemotactic protein-1 (MCP-1/CCL2) (63), which has been reported to be associated with decreased disease-free survival in OSCC (64).
A significant inverse correlation has been reported between the Ki-67 level and the stroma/tumor proportion and degree of keratinization (65).
Biology of OSCC presented by SBGN
Systems biology has become an inevitable part of data interpretation and presentation in research based on complexity. In the near future, it will undoubtedly become a potent tool in communicating the achievements of clinical research and will probably very soon take part in decision-making processes regarding personalized medicine. In this paper, the collected data on the interactions between stromal biomarkers associated with OSCC progression are presented using SBGN, whose main advantages are its clarity and wide applicability (66). This set of 3 languages was developed a few years ago and is accompanied by detailed technical specifications and a software application that makes it simple to use. This paper uses the Activity Flow language of SBGN, the simplest of the 3 and perfectly suited to the “bird view” description of disease mechanisms. Figure 1 shows OSCC in its microenvironment as well as the interaction of the molecules described in this paper, known to play a significant role in the etiology and progression of this cancer type. This is the first time that SBGN has been applied for clinical purposes and we believe it has shown a unique capacity not only to summarize data but also to examine biological pathways in the contexts of the living environment and gender on the cellular, stromal and matrix level.
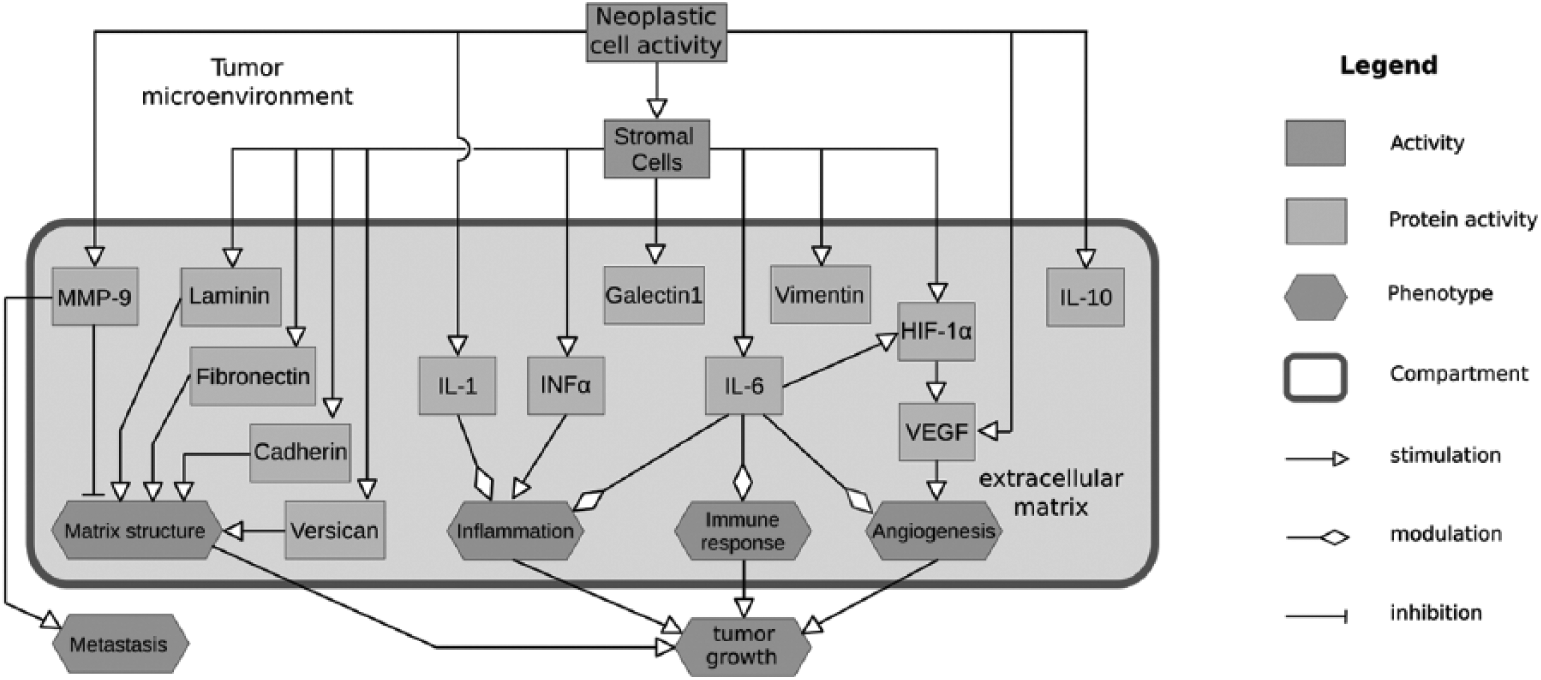
Interactions between EGFR, AR, ER, Ki67, INFα, Il-1, Il-6, VEGF, MMP9 and gender in OSCC tumour microenvironment presented by SBGN.
Conclusion
Understanding the interaction between neoplastic cells and their microenvironmental niche opens up new possibilities in OSCC diagnosis, prognosis and therapy. Although OSCC incidence and survival are gender related, the cross-reaction between estrogen and androgen receptors, estrogen and testosterone levels, and relevant stromal factors and proteins has so far not been investigated in OSCC. Similarly, following the knowledge gained from research into other cancer types, the induction of cytokines, MMPs, ECM and growth factors should be integrated with other factors relevant for early OSCC profiling and success of therapy. The level of stromal activation could become a significant factor for histopathological diagnostics as well as for decisions on the treatment and monitoring of OSCC patients. Stromal secretome profiling will contribute to a better future exploitation of the tumor microenvironment as a novel therapeutic target.
Footnotes
Abbreviations
CAF cancer-associated fibroblast
ECM extracellular matrix
HIF hypoxia-inducing factor
IL interleukin
MMP matrix metalloproteinase
OSCC oral squamous cell carcinoma
SBGN Systems Biology Graphical Notation
SMA smooth muscle actin
TAMC tumor-associated mast cell
TGF transforming growth factor
TNF tumor necrosis factor
VEGF vascular endothelial growth factor
Disclosures
Financial support: This project was supported by the Croatian Science Foundation under the title “The role of androgen and estrogen receptors in active stroma of oral cancer and their influence on patients’ survival” (I-1925-2015).
Conflict of interest: The authors have no conflicts of interest related to this article.