
Abstract
The social inhibition of return (sIOR) effect refers to the finding that response initiation times are longer if a movement is executed to a location where another person has responded to just before. Previous studies have examined the influence of the goal of the action on sIOR. In these studies, however, the movement endpoint and to-be-attained goal (e.g., touching/pressing a response key) were at the same spatial location. In the present two experiments, we disentangled movement endpoint and goal's identity and locations by means of introducing action effects that followed directly from a movement. Similar methods were previously shown powerful enough to clearly show the importance of action goals for other phenomena—a finding consistent with effect-based theories of action control, such as the ideomotor theory. The results of the present study revealed that sIOR was shaped by the movement endpoint location, not the goal's identity or location. That is, in both experiments, an sIOR effect was observed, but the magnitude of the sIOR effect was not modulated by repetitions/switches of goals or their locations. Thus, results indicate that goals play a negligible role in the emergence of the sIOR and, consequently, highlight the importance of action observation for the emergence of the sIOR effect.
Keywords
Human behaviour commonly occurs in social environments and requires some form of motor output. Research on human visual cognition and action control has recently been expanded to include studies of movements in such social contexts. In this research, classic paradigms of cognitive psychology are adapted such that they are completed by two participants with instructions and task requirements distributed across them (see Knoblich, Butterfill, & Sebanz, 2011; Wenke et al., 2011, for reviews). One particularly interesting example of this research concerns the inhibition of return (IOR) effect. In tasks typically used to investigate the IOR effect, participants respond to targets that randomly appear at one of several potential locations. The IOR effect refers to the finding that response initiation times (RTs) are longer to targets that appear at the location of another previous cue (Posner & Cohen, 1984) or target event (Maylor & Hockey, 1985) than if the target appears at a novel location. Research on the IOR effect in individuals acting alone has led to the suggestion that the longer RTs for repeated stimuli occur because inhibitory mechanisms are activated that hinder the return of attention and/or the reactivation of a response to the preceding stimulus location. It has been suggested that the mechanisms leading to IOR have developed through evolution to facilitate efficient visual search patterns because the inhibitory mechanisms discourage the reinvestigation of previously searched locations, thereby facilitating the investigation of new locations (e.g., Klein, 1988; see Klein, 2000, for a review).
Because humans have evolved and developed in social groups, it is very likely that people performed searches of the environment alongside other people. Consequently, it was reasoned that if the IOR effect reflects mechanisms that prevent reinvestigation of a location that has already been investigated recently, then a similar effect may be present when another individual has responded to a particular location immediately before—that is, an instance of social inhibition of return (sIOR; Welsh et al., 2005, 2007). In the initial experiments investigating this possibility, two participants sat in front of each other with their right index fingers resting on a home button located directly in front of each of them. Two further response keys were midway between both participants, to the left and right of a central point. The illumination of the response key signalled the target for a trial and prompted one participant to depress the respective key as fast as possible. During a block, a participant gave two subsequent responses followed by two responses of the partner, and so on (e.g., AABBAA …). It has been found that participants typically have longer RTs (a) when they respond to the target they themselves have responded to immediately before (IOR), and also (b) when their partner has responded to the same target immediately before (sIOR). Thus, the observation of another person's response appears to activate the mechanisms that lead to IOR much in the same way that the execution of a response leads to IOR (see Welsh, McDougall, & Weeks, 2009).
Visual transients versus response observation in sIOR
Because the IOR effect can be generated by the capture of attention from a variety of dynamic transient stimuli, such as abrupt onset (Posner & Cohen, 1984) and motion onset (Abrams & Chirst, 2003), as well as the execution of a previous response (Maylor & Hockey, 1985), subsequent experiments sought to determine the information necessary for (and thereby the mechanisms leading to) the sIOR effect. Work by Welsh et al. (2007) and Skarratt, Cole, and Kingstone (2010) was designed to determine whether observation of the response was sufficient for the generation of the sIOR effect, or if the sIOR effect was dependent on the nonsocial visual transients associated with target onset and motion of any stimulus (i.e., the hand) to and from the target. To this end, the amount and type of visual information available to the participants when observing their coactors’ action was manipulated. In the key conditions of these studies, participants only observed an initial portion of the movement towards the target because goggles with opaque lenses (Welsh et al., 2007) or physical barriers (Skarratt et al., 2010) were used to prevent the observer from seeing the abrupt onset of the target and the latter portion of the hand motion to and from the target, including target contact. It was found that sIOR emerged under these restricted vision conditions thereby providing some initial evidence that the mechanisms of sIOR are not dependent on visual transients occurring at the target location. Additional research revealed that sIOR emerged when coactors completed movements in similar directions even though the movements were executed to different targets (Welsh et al., 2009).
Because sIOR emerged under conditions in which visual transients were not observed at the target location but target-directed actions were observed, the observation of a response to a target location was suggested to be the key and, hence, sufficient information to generate sIOR (Welsh et al., 2005, 2007). It was further speculated that an action observation system, perhaps akin to the mirror neuron system that has been reported in monkeys (e.g., Rizzolatti & Craighero, 2004), would play an important role in recognizing and coding the observed actions, and hence plays an important role 1
It should be noted that this finding does not indicate that response observation and response corepresentation are the only mechanism that can lead to IOR in social context. For example, it has been argued that the sIOR effect could be due to attentional orienting rather than response observation. In the case of sIOR when peripheral target information is occluded, it could be argued that the initial (visible) part of a coactor's arm movement is a central cue that shifts attention to the target location (Atkinson et al., 2014; Doneva & Cole, 2014). The relative and/or independent contributions of this is currently unclear. We thank a reviewer for pointing this out and briefly readdress this point in the General Discussion.
Goals and the sIOR effect
Because the action observation system of monkeys (e.g., Kohler et al., 2002; Umilta et al., 2001) and humans (e.g., Buccino et al., 2001) seems to be sensitive to goals, it appears possible that cognitive effects that are assumed to depend on these systems are modulated by goals. Thus, if an action observation system that is sensitive to goals is responsible for the sIOR effect, then the sIOR effect may be sensitive to goals as well. This reasoning has led to a set of studies exploring the influence that goals have on the sIOR effect (e.g., Cole, Skarratt, & Billing, 2012; Welsh, Manzone, & McDougall, 2014).
It is generally accepted that humans perform most movements to attain certain goals. Goals, however, can be conceptualized on various levels of abstractness and proximity. For example, to achieve the goal of finalizing this manuscript, the achievement of numerous other, more concrete, goals such as producing particular letters on the screen by means of moving fingers to particular keys is required. Within the philosophy of mind, such distinctions are captured by referring to different levels of intentions. Searle (1980, 1983) distinguishes (rather abstract) prior intentions from intentions-in-action, which are linked to a specific movement. Similarly, Pacherie (2008) distinguishes distal, proximal, and motor intentions on the way from abstract goals to specific movements. For the present purposes, we distinguish action goals from action effects in the following way: Action goals refer to more abstract goals to be achieved within a given task (e.g., finishing a manuscript or following the instructions of an experiment). These goals are linked to, for example, prior intentions. Action effects, in contrast, refer to the perceivable effects contingently produced by a movement in the course of an action. 2
Action effects are essentially a subcategory of action goals, namely goals that directly follow from an action. Instead of using distinctions such as high- versus low-level action goals, we prefer goals versus effects as defined here, because the term action effect is well established in the literature on ideomotor theory (described in the next section), and action effects are what we actually manipulated in the experiments.
Some preliminary evidence for a role of action goals comes from a study by Hayes, Hansen, and Elliott (2010). In this study, participants responded and initiated their movements as fast as possible upon illumination of a stimulus (light). Critically, they performed a movements towards the illuminated location in some conditions or to the location opposite from the illuminated location in other conditions. One might argue that participants then had either the same action goal (both responded to the illuminated location) or different action goals (one participant responded to the illuminated location while the other responded to the opposite location). The results revealed a smaller sIOR effect when participants had different goals than when they had similar goals. The reduced sIOR effect in a condition when both participants had different goals suggests that action goals may indeed be effective in modulating the sIOR effect.
A further study more directly investigated the interplay of action goals and action effects/movements within the sIOR paradigm (Ondobaka, de Lange, Wiemers, Newman-Norlund, & Bekkering, 2012). Participants pursued either the same or a different action goal, and—manipulated orthogonally to this variation—this required a movement to the same or a different location as a coactor (similar to the standard sIOR task). Two findings were noteworthy. First, longer RTs 3
In fact, the dependent variable in this study was measured from stimulus onset until touch of the endpoint location (see their online Supplemental Material); thus the dependent measure was total response time—the sum of reaction time (stimulus onset until movement onset) and movement time (movement onset until touch of movement endpoint location). Hence, this dependent measure differs from what is used in other sIOR studies, which typically distinguish between reaction time and movement time.
Despite this initial evidence of a role for goals in sIOR, there are also data indicating that such action goals play no role in the emergence or modulation of sIOR. Using a variant of the sIOR paradigm, Cole et al. (2012) conducted a series of experiments where they explicitly defined goals as “the terminal component of a reaching action” (p. 738). As in the standard procedure, participants were required to reach to a cued location. The important manipulation (compared to previous attempts) was that participants had to complete a specific task (goal) at the target location or with an object at the target location. Across the experiments, participants were required to either touch or pretend to grasp a diamond (Experiment 1), pick up a real cup and pretend to either drink or throw liquid over their shoulder (Experiment 2), or pick up a pencil and either write with it or erase a written character (Experiment 3). Participants performed either the same or different goal actions at the same two target locations. In each experiment, an sIOR effect was found, and, importantly, the magnitude of the sIOR effect was similar regardless of whether both participants had the same or different goals. Because an sIOR effect was (a) present in all conditions and was (b) not modulated by goal compatibility, the authors concluded that the role of action goals in the emergence of the sIOR effect is negligible or nonexistent. This conclusion apparently stands in contrast to the broad acknowledgement of goals for human actions.
In sum, the evidence for a role of action goals for the sIOR effect is rather mixed at best. One factor that might be leading to the different sets of results is the relationship between the spatial location of the response and the location of the “goal”. To address this issue and take a new approach to the study of the role of goals in sIOR, the present research investigated the role of goals in sIOR by focusing on action effects. In sum, the unique approach of the present work was to employ a method in which the spatial location of the response was different from the spatial location of the goal. Thus, while the broad topic of this research can be subsumed under action goals, it is important to note that the particular question and approach was different from those in the aforementioned studies. To enable an understanding of the rationale behind this approach, an introduction to ideomotor theory is provided in the next section.
Ideomotor theory and action selection
Many studies in recent cognitive psychology have dealt with the question of how particular bodily movements and actions are selected, addressed, and initiated. Ideomotor theory of action control ascribes a crucial role in action selection to the events that immediately follow from actions—their action effects (Harleß, 1861; Hommel, Müsseler, Aschersleben, & Prinz, 2001; Shin, Proctor, & Capaldi, 2010; for a brief history see Stock & Stock, 2004). Briefly, such accounts are based on the tenet that an action is (only) accessible by anticipating its sensory consequences—that is, by anticipating the perceptual effects of the action (Elsner & Hommel, 2001; Kunde, 2001; Wolfensteller & Ruge, 2011; but see Herwig, Prinz, & Waszak, 2007, for possible constraints). In other words, action control operates on the perceptual representations of events, which appear to be independent of the source (i.e., the owner) of generation (Dolk, Hommel, Prinz, & Liepelt, 2013, 2014; see Dolk, Hommel, Colzato et al., 2014, for a review). Critical to the present study, the bodily movements and their effects can be spatially separable. For example, several studies have revealed that meaningful action–effect associations can be developed between key presses and tones (Elsner & Hommel, 2001) and between key presses and lights on opposite sides of space (Hommel, 1993; see also Kiernan, Ray, & Welsh, 2012).
Overall, an impressive amount of evidence has accumulated suggesting the merit of these ideas with responses including key presses, vocal utterances, continuous movements, and even facial expressions (Badets, Koch, & Toussaint, 2013; Chen & Proctor, 2013; Janczyk, Yamaguchi, Proctor, & Pfister, 2015; Koch & Kunde, 2002; Kunde, 2001, 2003; Kunde, Lozo, & Neumann, 2011; Kunde, Müsseler, & Heuer, 2007; Kunde, Pfister, & Janczyk, 2012; Paelecke & Kunde, 2007; Pfister, Janczyk, Wirth, Dignath, & Kunde, 2014). Of importance for the present study, several studies have shown that action effects also account for more complex phenomena such as the determination of the Simon effect (Hommel, 1993) and the determination of the size and direction of specific dual-task interference (Janczyk, Pfister, Hommel, & Kunde, 2014). In all these aforementioned studies, the action effects were produced by, and are thus under the control of, the actor. However, a similar case for the role of action effects has also been made for joint task performance in which action effects can be produced by other (social or nonsocial) entities and are, thus, not exclusively under the actor's control (e.g., Dolk et al., 2013; Dolk, Hommel, Prinz, et al., 2014; Kiernan et al., 2012; Pfister, Dolk, Prinz, & Kunde, 2013).
More generally speaking, intentionally generated action effects can be considered a subclass of action goals that can easily be manipulated and controlled in the laboratory. In the typical sIOR setting (Welsh et al., 2005, 2007), it appears that participants pursue the production of action effects related to depressing the left or right response key. To achieve this goal (i.e., the perceivable and intended effect of a given action), a movement to the very same location is required. As a consequence, both movement endpoint and action effect locations are necessarily confounded, regardless of the role of action goals (e.g., write or erase a letter as in Cole et al., 2012, or select a higher or lower card as in Ondobaka et al., 2012). The colocation of movement endpoint and action effect is by no means a flaw in the design because their spatial aligning was an experimental necessity. On the other hand, this arrangement does not allow one to determine their independent influences on sIOR. Thus, until now it is unclear whether (a) the location of the to-be-pursued action effect or (b) the location of the endpoint of the required movement is responsible for the sIOR effect. Welsh et al. (2007) reported an sIOR effect in conditions where participants only saw a brief and early period of the partner's response and concluded that “knowledge of the other person's behavior” (p. 955) is critical. However, this is not necessarily the case because movement endpoints and action effects were spatially confounded as well (but see Welsh et al., 2009). Thus, it could also be that inferring the partner's action effects was responsible for (the emergence of) the sIOR effect.
The present study
In the present study, we continued research on the role of goals for the sIOR effect, but we focused on the influence of action effects as a means of dissociating the spatial location of the response and of the goal. To this end, we introduced “effect lights”, and the participants’ task was to switch on these effect lights. Importantly, the location of the effect lights was spatially separated from the movement endpoints (e.g., Hommel, 1993; Kiernan et al., 2012). Experiment 1 used a set-up similar to the standard sIOR task (e.g., Welsh et al., 2005, 2007) with the addition of central (red or green) effect lights. Experiment 2 used a method that further disentangles the locations of action effects and of movement endpoints. While for one participant, the effects occurred at the location of the movement endpoint, for the other participant the effects occurred at the location opposite to the movement endpoint. A similar approach was used by Hommel (1993) to investigate whether the Simon effect reflects (spatial) compatibility between the stimulus and the movement endpoint or the stimulus and the action effect. When movement endpoints and action effects spatially coincided, a standard Simon effect was observed. However, when the action effects appeared at the side opposite to the movement endpoint, the Simon effect was reversed (at least when instructions emphasized the action effects). These results suggest that the (spatial) compatibility between stimulus and action effect is important and that the location of the goal (the goal itself) may dominate when participants are instructed to focus on generating the goal. Notably, similar findings have been made with the social Simon effect (Kiernan et al., 2012) and the backward crosstalk effect in dual-task situations (Janczyk, Pfister, Hommel, et al., 2014). Accordingly, the set-up was slightly modified, and now the central light's colour indicated which of the two peripheral lights was to be switched on. In both experiments, participants neither saw nor heard the touch of the response key of the other participant. Importantly, instructions were phrased in a way that participants understood their task was to “switch on a light”. This way of directly emphasizing the action effects has been proven effective in boosting the impact of action effects and separating the spatial coordinates of the movement endpoint and the action effect in previous studies (Hommel, 1993; Janczyk et al., 2015; Kiernan et al., 2012; Shin & Proctor, 2012).
In both experiments, a goal-congruent control condition and a goal-incongruent experimental condition were implemented. Within the goal-congruent conditions, a repetition of the movement endpoint yielded a repetition of the effect identity (colour; Experiment 1) or the effect location (Experiment 2). Thus, movement endpoint and effect were purposefully confounded so that an sIOR effect in the typical direction can be expected. Within the goal-incongruent conditions, the repetition of movement endpoints and action effect identity or location were disentangled. If goals and their locations are indeed not critical, and the observation of the response has a larger role in sIOR, then the magnitude and direction of the sIOR should be similar in goal-congruent and goal-incongruent conditions: longer RTs for repeated than for different movement endpoints independent of the location or identity of the action effects. In contrast, if action effects (and not movement endpoints) are the driving force, the sIOR should be reversed in the goal-incongruent condition—longer RTs for repeated than for different action effects independent of movement endpoints.
Experiment 1
As a first approach, we ran an experiment with a set-up very similar to the standard sIOR task, that is, the target location was indicated by peripheral lamps. The only modification to previous studies (e.g., Welsh et al., 2007) was a central light that could be illuminated in either red or green colour. In the goal-congruent condition, the response keys of the two participants that were located at the same side of the set-up triggered the central light being illuminated in the same colour such that a repetition of the movement endpoint implied a repetition of the action effect's identity. We expect a standard sIOR in this condition. In the goal-incongruent condition, the same colour of the central light was attained by the two participants by touching the opposite response keys. In this case, repetition of the movement endpoint implies a switch of the action effect's identity, and a switch of the movement endpoint implies a repetition of the action effect's identity. If action effects do not influence the sIOR, size and direction of the sIOR should be equivalent in both the goal-congruent and the goal-incongruent condition. A reversal of the sIOR in the goal-incongruent condition, however, would indicate that the action effect (and thus more broadly: the goal) is more relevant to the emergence of the sIOR effect than the endpoint of the movement. Consequentially, what counts then is not the repetition of the movement endpoint, but rather the repetition of the action effect. Any pattern of results in between these two extremes, as indicated at least by a significant interaction, would suggest a role of the action effects as well as other sources of information (such as the movement) affecting the mechanisms yielding the sIOR effect.
Method
Participants
Thirty-two undergraduate students (mean age = 29.1 years, 23 female) from the University of Würzburg participated for course credit or monetary compensation. Participants were recruited in pairs and reported normal or corrected-to-normal vision. Informed written consent was obtained prior to experimentation, and participants were naïve regarding the hypotheses of this experiment.
Stimuli and apparatus
The experimental set-up is depicted in Figure 1. Home buttons were custom-built keys that could be depressed and released, and response keys were touch-sensitive plates. To prevent participants from hearing when the other participant made his/her response, all participants wore sound-attenuating headphones. Two peripheral and one central light emitting diodes (LEDs) were placed on the table. The centrally located light served as the (visual) action effect in this experiment and was illuminated either green or red; the two peripheral lights served as the imperative stimuli and were illuminated yellow until a home button was released (movement initiation). All lights were visible to both participants (as is the case for the peripheral lights in the standard sIOR paradigm). A visual barrier placed in between the participants prevented them from seeing the other participant touching the response key, while a gap (7.5 cm) in the barrier allowed participants to see the initial portion of the movement. The barrier thus prevented the participants from observing their partner contacting the button while still providing sufficient information to infer the endpoint of their movement. RT was defined as the time interval from the onset of the stimulus light until the participant's finger left the home button. Movement time (MT) was defined as the time interval from release of the home button until a response key was touched.
Illustration of the experimental set-up. In most aspects, the set-up was similar to the one used by Welsh et al. (2005) and Skarratt et al. (2010), except that an additional stimulus lamp (“central light”; which could be illuminated either green or red) was located centrally. The two peripheral lights were illuminated in yellow colour. To view this figure in colour, please visit the online version of this Journal.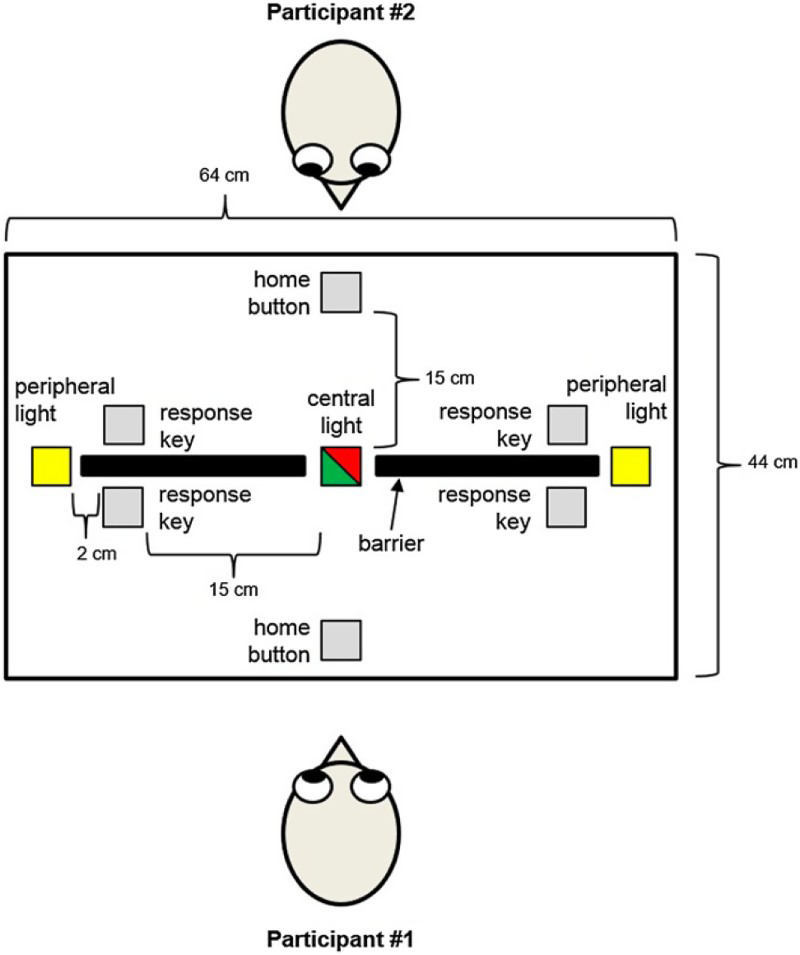
Procedure
Participants were tested in pairs in a single session of about 45 min. The experiment was completed in a dimly lit experimental room. Both participants were assigned their seat and received written instructions. The task was to illuminate the central light in red or green colour by touching the response key located close to the illuminated stimulus light. Participants were told to respond in an alternating order. The instructions emphasized speed in responding while maintaining errors at a low rate, but made no mention about which response key triggers which colour of the central light. The mapping of response key to light colour was briefly explored by both participants subsequently. In the goal-congruent condition, the response keys on the same side of the board triggered the central light to be illuminated with the same colour. In other words, a repetition of the movement endpoint implied a repetition of the action effect colour. In the goal-incongruent condition, the response keys on the opposite sides triggered the central light to be illuminated with the same colour. Thus, a repetition of the movement endpoint implied a switch of the action effect colour and vice versa. Each trial started when both home buttons were depressed. Then, one of the stimulus lights was illuminated yellow until a home button was left. Participants were to respond by turns, always starting with the same participant. Touching the required response key immediately illuminated the central light in the designated colour for 500 ms. Participants were to immediately return to their home button and depress it. Following another 500 ms after offset of the effect light, the next trial began. If participants responded in the wrong order or if one participant gave a wrong response, the current block was terminated immediately.
Each pair of participants started with six unanalysed practice blocks of nine trials to allow them to become familiar with the task and learn the response–effect mapping. Forty experimental blocks with 17 trials each were administered subsequently with a break and change of condition after 20 blocks. The mapping of stimulus location (and thus movement endpoint) to central light colour and order of conditions were counterbalanced across participants.
Design and analyses
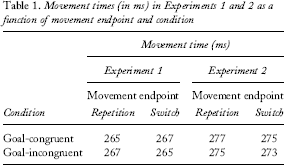
Although the main dependent measure was RT, analyses were also conducted on MTs for completeness and to ensure that any condition differences in RT were not the results of an RT/MT trade-off. Only correct trials were considered for RT and MT analyses. Trials were considered invalid if MTs were greater than 800 ms (0.15% of the data) and when RTs were less than 100 ms or greater than 800 ms (additional 1.40% of the data). Data were classified according to whether or not the same response key as that in the previous trial of the partner was touched (i.e., movement endpoint repetition vs. switch). Note that the first trial of a block was excluded because it is neither a repetition nor a switch.
Results
Mean correct RTs are illustrated in Figure 2 (left panel), and RTs and MTs were submitted to separate 2 × 2 analyses of variance (ANOVAs) with movement endpoint repetition (repetition vs. switch) and condition (goal-congruent vs. goal-incongruent) as repeated measures. Mean RTs were comparable in both conditions, F(1, 31) < 0.01, p = .957, Illustration of mean correct reaction times (RTs) as a function of movement endpoint repetition and condition (left panel: Experiment 1, right panel: Experiment 2). Error bars are 95% within-subject confidence intervals calculated separately for each condition (Pfister & Janczyk, 2013).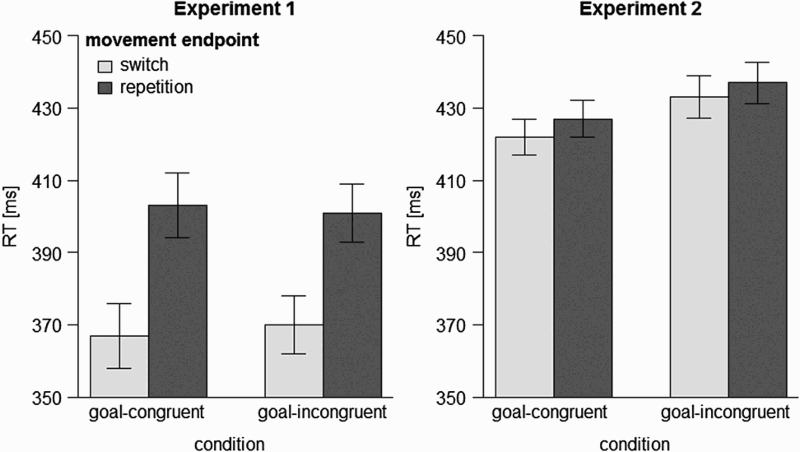
Movement times (in ms) in Experiments 1 and 2 as a function of movement endpoint and condition
Comparable numbers of blocks were terminated due to errors in the goal-incongruent condition (5.9%) compared to the goal-congruent condition (10.0%), t(15) = 1.81, p = .091. In all except one case, blocks were terminated due to an erroneous response order.
Discussion
As expected, the goal-congruent condition of Experiment 1 yielded a stable sIOR effect. Thus, the data provide a verification of our particular experimental set-up. The theoretically more interesting result, however, is that the sIOR effects in the goal-congruent and the goal-incongruent condition were of similar magnitude and direction. This finding implies that the movements per se—not the goal states as manipulated by action effects—determine the sIOR. Given the amount of evidence suggesting that goals impact human behaviour in individual and social contexts (e.g., Hommel, 1993; Hommel et al., 2001; Janczyk, Pfister, Hommel, et al., 2014; Kiernan et al., 2012; Kunde, 2001), such a result was not entirely expected. Although the present findings are consistent with the conclusion of other recent studies on the role of action goals for the sIOR effect (Cole et al., 2012; Welsh et al., 2014), there are several possible reasons for the present findings. First, it might be that the experimental manipulation of goal-(in)congruency and the action effects were not sufficiently effective and strong enough to drive action effect coupling in the social setting (cf. Kiernan et al., 2012). In fact, there was no main effect of condition, which would have indicated an overall effect of the experimental manipulation. Second, given that the peripheral lights—indicating the response—were visible to both participants, it may be that the illumination of the LEDs drove a capture of attention that led to an IOR effect on its own regardless of the partner's movement and action effect.
It should be noted that, even though attentional capture by the peripheral light might have been the main driver of the sIOR, the action effects (i.e., the additional abrupt onset of the central light) did not modulate the sIOR effect. Thus, the results of Experiment 1 do not support a proposed role for goal-based modulations of the sIOR effect. Nonetheless, because the imperative stimulus (peripheral light) and the movement endpoint location were confounded, their role for the sIOR effect could not be completely separated with the design of Experiment 1. For this reason, the imperative stimulus and the movement endpoint location were separated in a second experiment.
Experiment 2
In Experiment 2, a manipulation that was shown to be very effective in disentangling movement- and goal-contributions in previous studies (Hommel, 1993; Janczyk, Pfister, Hommel et al., 2014; Kiernan et al., 2012) was adapted for the present research question. Instead of presenting the imperative stimulus in the periphery and the action effect in the centre, the roles of the lights were reversed such that the effect light that was to be illuminated (and hence the correct movement endpoint) was determined by the colour of the central light, whereas touching a response key triggered a peripheral light to be illuminated as the action effect. Instructions highlighted the action effects, and participants were instructed to illuminate the left or right peripheral light depending on the colour of the central light. In the goal-congruent condition, touch of a response key caused the nearby peripheral light to be illuminated. This condition was important as a baseline to test whether an sIOR effect can be found following a central, symbolic stimulus. Note that, although the observing participant did not see her/his partner touching the button due to the barriers, the observer did witness the abrupt onset of the nearby peripheral light. Thus, similar to Experiment 1, an sIOR effect in this condition might be due to the response and/or the attention-capturing abrupt onset associated with the illumination of the peripheral light. In the goal-incongruent condition, the association of movement endpoint/response key and action effect light was reversed for one participant, meaning that touching the response button always triggered the illumination of the opposite effect light. Thus, in this condition, the stimulus, the endpoint of the movement, and the location of the action effects were separated. The logic and predictions for this condition were the same as those in Experiment 1.
Method
Participants
Thirty-two new undergraduate students (mean age = 21.1 years, 28 female) from the University of Würzburg participated for course credit. Other details were the same as those for Experiment 1.
Stimuli and apparatus
The experimental set-up was the same as that in Experiment 1 (see Figure 1) except the assigned role of central and peripheral lights. In contrast to Experiment 1, the centrally located light was now used as the stimulus light, and the two peripheral lights were the effect lights.
Procedure
Participants were assigned their seat and received written instructions about which effect light (i.e., the yellow lights to the left/right of the barrier) they were to switch on in response to the colour of the central (green/red) stimulus light—the stimulus–effect mapping. Instructions explained that participants were to respond in an alternating order and emphasized speed in responding while maintaining errors at a low rate, but made no mention about which response key triggers which effect light. Following those instructions, participants were given the opportunity to explore which response key will trigger which effect light. In the goal-congruent condition, both participants experienced that the response key adjacent to the respective effect light was effective in illuminating that LED. In other words, both participants were experienced with a compatible response–effect (RE) mapping. 4
In the goal-incongruent condition, the mapping was changed after the first half of the blocks, and thus this procedure was repeated. To equate both conditions, this “experience”-procedure was also repeated after the first half of the blocks in the goal-congruent condition.
Each trial started, if both home buttons were depressed, with the stimulus light being illuminated either red or green until a home button was released. Participants were to respond in turns, always starting with the same participant. Touching the required response key immediately made the goal light be illuminated for 500 ms, and participants were to immediately return to their home button and depress it. Following another 500 ms after offset of the effect light, the next trial began. A block was terminated if participants responded in the wrong order, or gave a wrong response to the colour of the stimulus light.
Each pair started with six unanalysed practice blocks of nine trials. Forty experimental blocks with 17 trials each were administered subsequently with a break after 20 blocks. The mapping of stimulus colour to effect light and the order of conditions were counterbalanced across participants.
Design and analyses
Only correct responses were considered for RT/MT analyses. Trials were considered invalid if MTs were greater than 800 ms (0.39% of the data) and when RTs were less than 100 ms or greater than 800 ms (additional 1.93%). Data were classified according to whether or not the same response key as that in the previous trial of the partner was touched (i.e., movement endpoint repetition vs. switch). Note that the first trial of a block was excluded because it is neither a repetition nor a switch.
Results
Mean correct RTs are illustrated in Figure 2 (right panel), and RTs and MTs were submitted to separate 2 × 2 ANOVAs with movement endpoint repetition (repetition vs. switch) and condition (goal-congruent vs. goal-incongruent) as repeated measures. Mean RTs were higher in the goal-incongruent than in the goal-congruent condition, F(1, 31) = 6.15, p = .019,
To rule out that the nonsignificant interaction was somehow compromised by the overall differences of the RT levels in both conditions, we ran this analysis again but included only Quintiles 1–4 from the RT distribution of the goal-incongruent condition. This procedure effectively reversed the main effect of condition with now faster RTs in the goal-incongruent than in the goal-congruent condition, F(1, 31) = 30.30, p < .001,
Overall, more blocks were terminated in the goal-incongruent condition (19.7%) than in the goal-congruent condition (10.9%), t(15) = 3.59, p = .003. In 75.5% of these cases, the error was a wrong response; in 24.5% it was an erroneous response order. This distribution did not differ between the two conditions, χ²(1) = 0.49, p = .484.
Between-experiment analysis
Visual inspection of Figure 2 reveals the sIOR effect to be much larger in magnitude in Experiment 1 than in Experiment 2. Collapsed across conditions in each experiment, the respective sIOR effects were 33 ms (Experiment 1) and 5 ms (Experiment 2). To corroborate this observation, we submitted the data to a mixed ANOVA with movement endpoint repetition as a repeated measure and experiment as a between-participants variable. RTs were higher for Experiment 2 than for Experiment 1, F(1, 62) = 9.13, p = .004,
Discussion
Experiment 2 yielded several important results. First, even though the magnitude of the sIOR effect was smaller than that following peripheral stimuli (Experiment 1), Experiment 2 is the first study to reveal an sIOR as a consequence of centrally presented, symbolic stimuli (see Figure 2 and the between-experiment analysis). The finding of a larger sIOR effect in Experiment 1 than in Experiment 2 might indicate that attentional capture by the peripheral stimuli contributed to the emergence and magnitude of the sIOR effect in Experiment 1, though it is unclear if it was the only process that leads to sIOR. Note, however, that this suggested role of attentional capture in Experiment 1 does not necessarily invalidate the theoretically relevant observation that goals did not influence the sIOR effect. Second, the main effect of condition provides evidence that the manipulation associated with RE compatibility indeed influenced participants’ behaviour. Third, despite this influence on overall behaviour and the fact that a very similar manipulation was effective in reversing the (social) Simon effect (Hommel, 1993; Kiernan et al., 2012) and the backward crosstalk effect in dual-task situations (Janczyk, Pfister, Hommel et al., 2014), there was again no evidence for any modulation of the sIOR effect by this manipulation. What is particularly interesting about the results of Experiment 2 is that the abrupt onset of the peripheral effect in the goal-incongruent condition occurred in the side of space opposite from the movement endpoint (for one participant). Such abrupt onsets typically capture attention and, as such, could have disrupted sIOR effects generated by the movement. The absence of a significant interaction between condition and movement endpoint repetition indicates that this abrupt onset did not modulate the sIOR effect associated with movement observation. Thus, it appears that the sIOR was primarily, if not entirely, determined by repetitions of the actual manual movements rather than of the goal states.
General Discussion
Previous research has demonstrated that RTs are longer when individuals respond to targets at a location that has just been responded to by a partner—a phenomenon known as the sIOR effect (e.g., Cole et al., 2012; Skarratt et al., 2010; Welsh et al., 2005, 2007). It is widely acknowledged that goals play an important role in shaping action planning. According to ideomotor theory, action effects (i.e., the immediate goal states achieved by an action) have a generative role: Actions are accessed and selected via anticipating their action effects, which can be considered a particular subclass of goal states (Harleß, 1861; Hommel et al., 2001; Kunde, 2001). With the importance of goals in mind, the present research investigated the role of goals for the sIOR effect by focusing on (and manipulating) action effects and their location relative to the location of the movement endpoint. Given that movement endpoints and action effect locations were spatially confounded in previous research, such attempts are particularly relevant for improving our understanding of the underlying mechanism yielding the sIOR effect.
Movement endpoint and action effect location were (spatially) disentangled in the present study, and the to-be-pursued action effects were clearly identifiable visual events in the environment. Even though the magnitude of the sIOR effect was larger when target information was provided by peripheral stimuli (Experiment 1) than with central, symbolic stimuli (Experiment 2), reliable sIOR effects were observed in both experiments for RTs. 6
For MTs there was no systematic variation depending on the independent variables. This might be taken to suggest that sIOR affects movement planning and initiation, but not the speed of movement execution.
In sum, our present results suggest that action effects appear to have no impact on the sIOR effect. 7
This does of course not mean that participants in an sIOR task are not performing actions but merely simple unintentional behaviour. Without participants’ carrying out an intention-in-action (Searle, 1980, 1983), no bodily movement will occur at all, and thus the self-referential component in the conditions of satisfaction of an action is fulfilled (see also Janczyk, Pfister, Wallmeier, & Kunde, 2014, for details). Our results, however, implicate that a repetition or switch of the effect's identity or location is not the critical moment for the sIOR and therefore helped resolving a confound in previous studies.
Although previous and present data suggest that goals do not shape sIOR, it is possible that the operationalization of same versus different action goals was rather too weak to exert a measurable influence on dependent measures in the Cole et al. (2012) study. There might actually be a difference between the operational definition of what Cole et al. defined as a goal (for example, in Experiment 3 where participants were asked to write or erase) and the goal as defined by the participants (i.e., reaching out to and picking up an object). In this case, it is possible that the participants attained their goal and succeeded with their tasks without taking the experimenter's goal definition into account. The data presented by Cole and colleagues are somewhat ambiguous as to whether this happened or not: If participants were representing the partner's (different) goal, one could expect longer overall RTs when both participants pursue different goals. Yet, this finding was only reported in one experiment (Experiment 3); in Experiment 1 participants had even shorter RTs in different goal conditions (though this effect did not surpass conventional levels of statistical significance, p = .074). Remember also that Ondobaka et al. (2012) did observe an influence of action goals, and the exact reason for this discrepancy with the Cole et al. (2012) study remains unclear at present. Apart from the use of different dependent measures in both studies (see Footnote 2), several other differences between both studies exist. First, in the Ondobaka et al. study, participants did not respond in alternation for several times in a row within the same block, but each trial consisted of the response of the coactor followed by the participant's response. Second, and probably more important, the current action goal of the partner was task relevant for the participants to derive their own action goal in the Ondobaka et al. study. Moreover, whereas in this study the participants were simply asked to touch a response button, in the Cole et al. study they had to perform a particular action at the movement endpoint location. All of these differences might be responsible for the different outcomes, but at present we can only speculate on why the differences emerged.
Although the present data are consistent with the conclusion that goals do not influence sIOR, the broader conclusion drawn here differs slightly from that of Cole et al. (2012). Specifically, Cole et al. suggested that the absence of an influence of goals provides evidence against action corepresentation being the core element underlying the mechanisms leading to the sIOR effect. It was reasoned that, because action observation systems are sensitive to goals (see, e.g., Rizzolatti & Craighero, 2004; or van der Wel, Sebanz, & Knoblich, 2013), the lack of sensitivity to goals in sIOR reveals that sIOR is not dependent on an action observation system. Instead, Cole and colleagues suggested that attentional shifts within the observer, not action corepresentation per se, are the main driver of the sIOR effect. In a follow-up study, Atkinson, Simpson, Skarratt, and Cole (2014) provided further evidence in favour of an attention system contributing to the sIOR effect. The present results, however, seem to support an important role for the observation of response-initiation in generating the sIOR effect. Evidence for this conclusion is drawn, in particular, from Experiment 2 in which the sIOR effect emerged after central stimuli and, importantly, was not modulated by attention capturing peripheral action effects in the environment. It thus appears that indeed the response per se or the initiation thereof is an important feature yielding an sIOR effect in the present study. In particular, one can conclude that observing the partner's movement—or at least inferring it from seeing its initiation—seems to induce an equivalent representation in the observer. This bidirectional principle of perception and action in turn can be considered responsible for the activation of inhibitory mechanisms that affect subsequent behaviour (Welsh et al., 2005).
At the same time, however, the larger sIOR effect in Experiment 1 than in Experiment 2 further suggests that action observation and attentional shifts are not mutually exclusive processes underlying the emergence of an sIOR effect. Yet, both contributed to the sIOR (especially in Experiment 1) although they might be relatively independent (see for additional discussion, Manzone, Cole, Skarratt, & Welsh, submitted). Still, the peripheral effect lights illuminated in Experiment 2 together with the fact that sIOR was not modulated by these attention capturing events makes a strong case for the crucial role of movement observation (or at least the initiation thereof) for the sIOR effect. In a complementary way, Welsh et al. (2014) showed that the mere generation of response-contingent auditory effects did not lead to an sIOR effect when the observer did not see any part of the response, again suggesting the importance of visual action observation for the sIOR effect.
In sum, action effects have been shown to be able to shape human performance in many varieties of tasks (e.g., Kunde, 2001) and also in social settings such as the social Simon effect (Kiernan et al., 2012). In contrast, our results—supported by those of Cole et al. (2012)—converge on the notion that sIOR might be a special case in that it is resilient to a role of action effects and or action goals in general (for another finding where movements rather than goals seem to be important, see Thomaschke & Dreisbach, 2013). Although not necessarily discounting a role of attention in sIOR, the present results do highlight an important role of movement observation in generating the sIOR effect.