
Abstract
The effects of distance above ground on fungal colonization during air-seasoning were evaluated over time on blackgum (Nyssa spp.) and red oak (Quercus spp. Section Lobatae) ties 150, 200, and 300 mm from the ground by culturing increment cores removed from three locations on each of 200 ties over the seasoning period. Fungi were present in many ties at the start of seasoning. Fungal isolations steadily increased with time, but there were no significant differences in frequency for ties seasoned at different sill heights for either timber species. The results indicate that lowering the sill height had no noticeable effect on the rates of fungal colonization of the timber species tested.
Introduction
Railroads in the United States continue to use wood for almost all railroad tie (sleeper) replacements because of its exceptional strength, flexibility, and renewable nature. Concrete ties are used sparingly in North American because they do not perform well with heavier trains and existing railbed foundations (Webster 1992; Webb and Webb 2016). More than 50 timber species are currently listed as acceptable for use as ties by the Railway Tie Association (RTA 2014; Webb and Webb 2016). The process of tie production begins with seasoning to remove moisture prior to chemical treatment. The vast majority of ties are air-seasoned prior to treatment, which is simple and inexpensive, but risks fungal degradation during the seasoning period (Thompson and Koch 1981; Conners 2008; Webb and Webb 2016).
The risk of fungal colonization during seasoning has long been recognized; some of the first studies on timber product decay were performed on ties using fungal fruiting bodies as decay indicators in seasoning yards (Mathewson 1930; Humphrey and Richard 1939; Mathewson et al. 1949; Taylor et al. 2013). The results of these early studies led to a number of recommendations for proper tie seasoning such as reduced seasoning time, tie stacking to permit maximum air-flow to accelerate drying, vegetation removal from the site, gravel use to minimize water pooling, and eventually, placement of tie stacks on preservative-treated timbers (sills) to limit ground contact. These procedures are listed in Standard M1 of the American Wood Protection Association Standards (AWPA 2017b) and in the recommended practices noted in Chapter 30 of the American Railway Engineering and Maintenance-of-Way Association Manual for Railway Engineering (AREMA 2019). Of particular interest in the AWPA Standards and AREMA recommendations is the requirement that the bottom layer of ties is supported at least 300 mm above ground on treated or otherwise decay-resistant sills. This requirement was designed to ensure untreated, seasoning ties were not exposed to direct soil contact or excessive splashing, but it created a secondary issue for seasoning yards. While seasoning yards are heavily mechanized to minimize the risk of injury, this sill height requirement is generally achieved by stacking two, 150 by 200 mm or 175 by 225 mm, treated crossties. The first crosstie is relatively easy to position, but manual placement is typical for the second, creating the risk of worker injury. A reduction in the sill height requirement could permit the use of a single crosstie; however, this would also move tie stacks closer to the ground, and could result in moisture conditions that increase the risk of fungal colonization during seasoning (Duncan and Lombard 1965; Conners 2008). There are surprisingly few data examining the relationship between sill height and fungal colonization in this application; although fungal colonization during air-seasoning is well correlated to strength loss (Kirker and Winandy 2014; Highley 1995; Taylor et al. 2013, 2015).
The purpose of this work was to examine the relationships between distance from the soil, environmental conditions, and fungal colonization of ties in a seasoning yard. We hypothesized that reducing the sill height would result in increased relative humidity around ties closest to the ground, and that these conditions would facilitate increased fungal colonization.
Materials and methods
The assessed ties were from a seasoning yard located in Guthrie, Kentucky. The site is located in AWPA Deterioration Zone 4 (but close to Zone 3) where Zone 1 is considered a low risk of decay and Zone 5 poses a high decay risk. The site is representative of many tie-seasoning sites in the region (AWPA 2017a; Kirker et al. 2017). This site would have a Scheffer climate index for above ground decay of approximately 60 where 0 is no risk of decay and 100 is considered extreme (Scheffer 1971). Ties were exposed between June 2017 and April 2018.
Ninety-eight freshly cut blackgum (Nyssa sylvatica) and red oak (Quercus rubra group) ties (225 mm by 225 mm by 2.55 m long) without obvious fungal colonization were selected for this study. Blackgum is considered susceptible to fungal colonization/degradation while red oak is slightly less susceptible (Scheffer and Morrell 1998). The ties were laid out, tagged for future identification, and three increment cores were removed on one wide face at the center and 450 mm inward from each end. This zone corresponds to the rail bearing area of an in-service tie where fungal damage would be most critical. Increment cores were placed in plastic bags, labelled with tie number and location, then stapled shut to minimize entry by other fungi. Increment core holes were plugged with tight-fitting, creosoted, wood dowels to minimize entry by other fungi through these holes.
Culturing
Fungal colonization provides a measure of susceptibility to fungal colonization during air-seasoning. Increased relative humidity and temperature as a result of proximity to the soil would be expected to improve conditions for fungal spore germination and subsequent mycelial growth in the wood (Zabel and Morrell 2020). This, in turn, would increase the probability that any given wood sample would contain a fungus. Increment cores from all ties were surface-flamed to minimize the growth of contaminating fungi encountered after core extraction. Each core was placed on benomyl-amended 1% malt-extract agar. Benomyl retards the growth of fast-growing, non-basidiomycetes (Eaton and Hale 1993). This is useful as nearly all major wood decay fungi are basidiomycetes (Duncan and Lombard 1965; Schwarze et al. 2013). For the purposes of discussion, basidiomycetes will be referred to as decay fungi although we recognize that they do not all-cause wood decay and that some ascomycetes also decay wood. Plated cores were examined for evidence of fungal growth over a one-month period, and decay fungi growing directly from the wood were isolated for later identification. Fungal isolates were counted as unique taxa per core sample where multiple taxa could be isolated from a single core. Potential decay fungi were segregated by growth morphologies as well as the presence of hyphal clamp connections and septa. Pure sub-cultured morphogroups were subjected to conservative DNA identification where multiple isolates of each morphogroup were sequenced (Cappellazzi et al. 2018). Fungal DNA was extracted from pure-culture vegetative mycelium using the cetyl trimethylammonium bromide (CTAB) protocol (Gardes and Bruns 1993) with minor modification.
DNA analysis
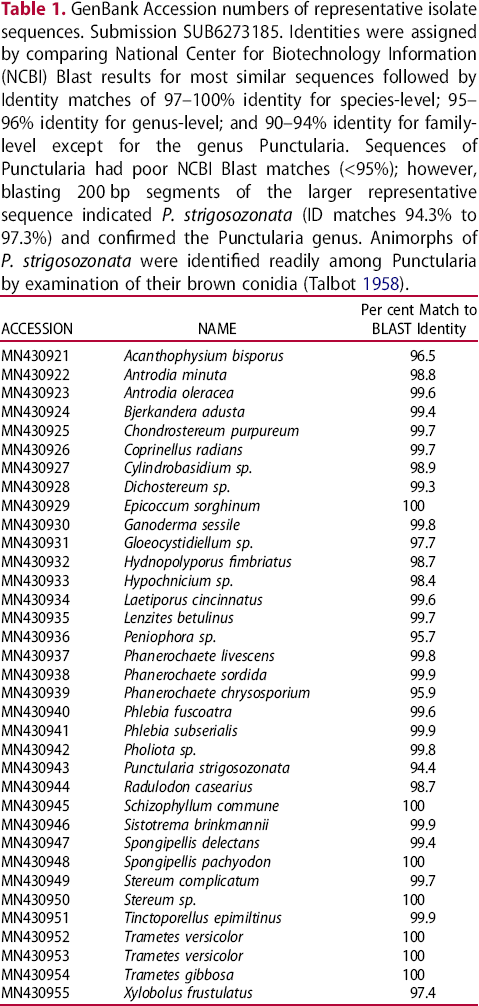
GenBank Accession numbers of representative isolate sequences. Submission SUB6273185. Identities were assigned by comparing National Center for Biotechnology Information (NCBI) Blast results for most similar sequences followed by Identity matches of 97–100% identity for species-level; 95–96% identity for genus-level; and 90–94% identity for family-level except for the genus Punctularia. Sequences of Punctularia had poor NCBI Blast matches (<95%); however, blasting 200 bp segments of the larger representative sequence indicated P. strigosozonata (ID matches 94.3% to 97.3%) and confirmed the Punctularia genus. Animorphs of P. strigosozonata were identified readily among Punctularia by examination of their brown conidia (Talbot 1958).
Stacking procedures
Ten ties of each species were shipped to Corvallis, Oregon to be characterized for initial material properties as part of a separate strength study (Rogers 2019). The remaining 88 ties of each species were placed into normal German-style seasoning stacks with treated sills that left ties a minimum of 150, 200, or 300 mm off the ground. A normal German-style stack has layers that each contained nine ties. The test ties were placed in courses of five test ties between two non-test ties within a layer to avoid edge effects. Only the first three layers above the treated sill received test ties. Non-test ties were then placed on top of the first three layers to create a full stack containing approximately 204 ties. Two stacks of each species were set at each sill height, creating 12 total stacks. Stacks were located near a riparian zone adjacent to a small creek. The site was well-drained, free of vegetation around the stacks, and did not contain any untreated wood debris that might serve as an inoculum source as per the current AREMA specifications (AREMA 2019). This test site was the most likely section of the yard to have high humidity to foster fungal growth, creating the worst possible site conditions within AREMA and AWPA recommendations. Relative humidity (RH)/temperature sensors (HOBO U23 Pro v2 Temperature/Relative Humidity Data Loggers, Onset Computer Corporation, Bourne, MA) were attached to the underside of ties in the first and third layer of a single stack at each of the three sill heights to monitor the effect of sill height on temperature and humidity. Sensor data were collected every 30 min between June 2017 and May 2018.
The gum stacks were disassembled and sampled after three months of air-seasoning using the same increment core procedures described above, while oak stacks were sampled after six months in recognition that this species requires a longer seasoning period (Conners 2008; AREMA 2019). Ties were returned to the same position in a respective stack after sampling. Gum ties were removed from the test after six months when they had reached a moisture content suitable for treatment, and transported to Corvallis, Oregon where a third set of increment cores were removed for fungal sampling. Oak ties were seasoned for an additional five months and similarly shipped to Oregon for sampling.
Statistical analysis
A total of 822 increment cores were removed from ties of each species over the exposure period (294 from 98 ties at time zero and 264 cores at both the interim and final sampling points). The isolation data were summarized by species, stack location (i.e. distance above ground), and seasoning time. The data were analysed for fungal community differences and isolate abundance in treatments (sill height) using ANOVA (α=0.05), multi-response permutation procedure (MRPP), indicator species analysis, and outlier analysis using PC-ORD software version 7 (Wild Blueberry Media, Corvallis, OR). Pairwise comparisons were not corrected for multiple comparisons and we expected some type-I errors given the number of comparisons made (Neyman et al. 1933; Mielke et al. 1976; Parchami et al. 2008).
Results and discussion
Environmental monitoring
There was a temperature/humidity sensor failure for the first three months of exposure, but data from the next eight months of sampling were usable.
Daily average temperature and relative humidity were fairly stable over the second three-month monitoring period (September to December 2018) (Figure 1). The average difference between sensors was less than 0.3°C and 1.5% relative humidity (RH) for the bottom three layers. As expected, temperatures decreased in the stacks through the Fall and Winter, but there were few differences between layers. RH values were also similar at various heights within the stack, although RH declined with increased air-seasoning time. This would reflect gradual tie drying, thereby decreasing the amount of moisture evaporating from wood surfaces. Neither RH or temperature varied noticeably with sill height, suggesting fungal colonization should not be influenced by the comparatively small environmental differences related to stack distance from the ground.
Temperature and relative humidity within stacks of air-seasoning blackgum and red oak on 150, 200, 300 mm-high sills. Error bars represent one standard deviation.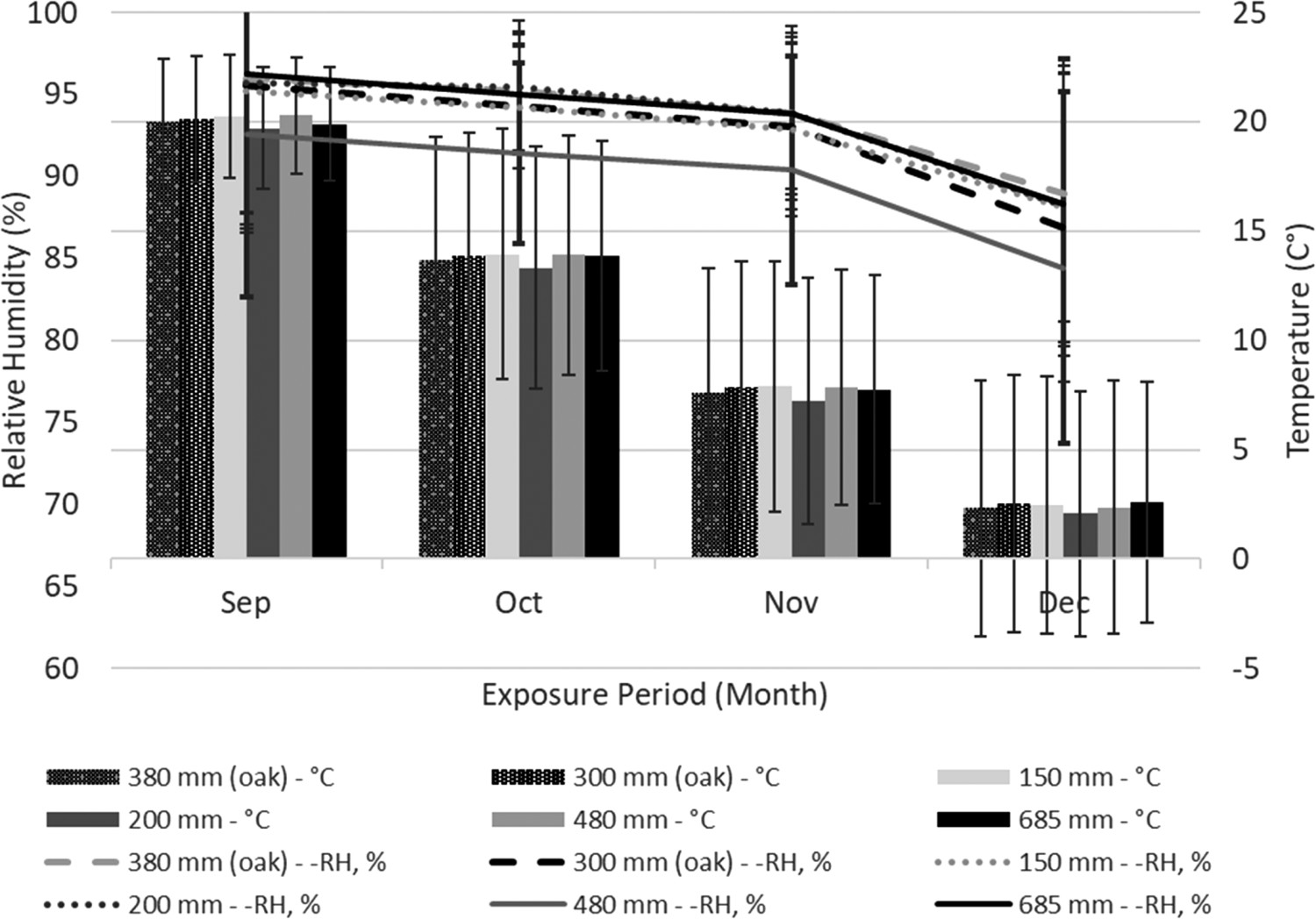
There was a slight RH increase when sensors were transferred to the red oak ties (Figure 2). It is unclear why this happened although temperatures were also lower during this period and RH increases as temperature decreases with a given amount of moisture in the air (FPL 2010). As expected, stack temperatures increased during the spring. Stack RH's were over 90% during the cooler months, then declined to around 70% as temperatures increased. Elevated RH's would be a concern for fungal colonization; however, they were associated with cooler temperatures. The results indicate stacks present a stable moisture and temperature environment that should be conducive to fungal colonization. There were slight, consistent differences in temperature and RH among sensors at differing sill heights or heights within the same stack, but they were not consistent with a height gradient; e.g. records at 325 and 840 mm were very similar.
Temperature and relative humidity within stacks of air-seasoning red oak ties on 150, 200, or 300 mm-high sills. Error bars represent one standard deviation.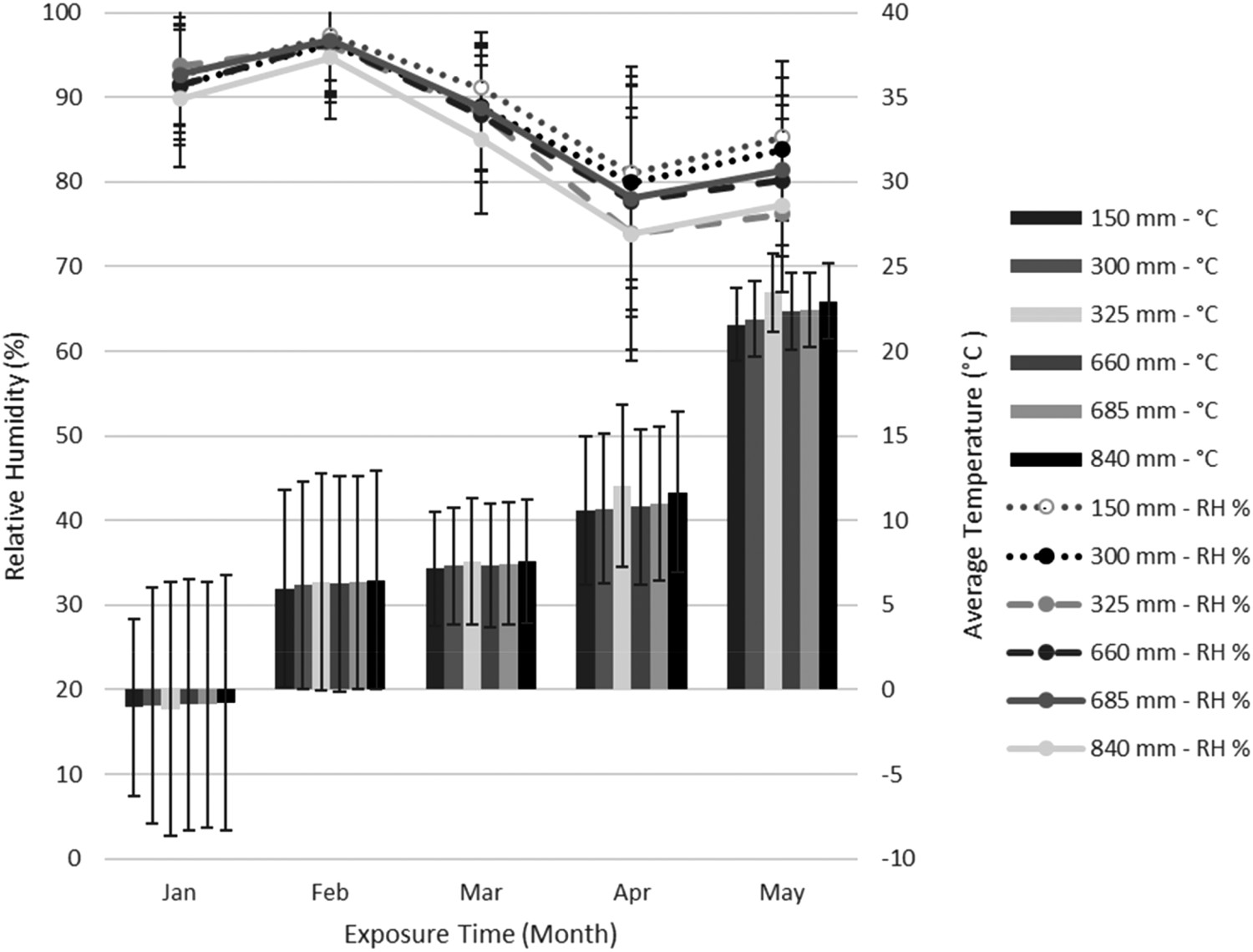
Fungal isolations
Blackgum: 31 fungal isolates were recovered from the gum ties at the start of air-seasoning. These represented six taxa, and over half of these fungi were isolated from ties placed in stacks 200 mm off the ground (Figure 3). Since ties were randomly allocated to stacks immediately after sampling, and the fungal culturing results were not available for 30 days, there was no way to initially allocate colonized ties evenly among sill heights.
Fungal frequency in blackgum ties air-seasoned for 6 months on German-style stacks that were 150, 200, or 300 mm off the ground at a treating plant in Guthrie, Kentucky (0, 3, and 6 months of air-seasoning).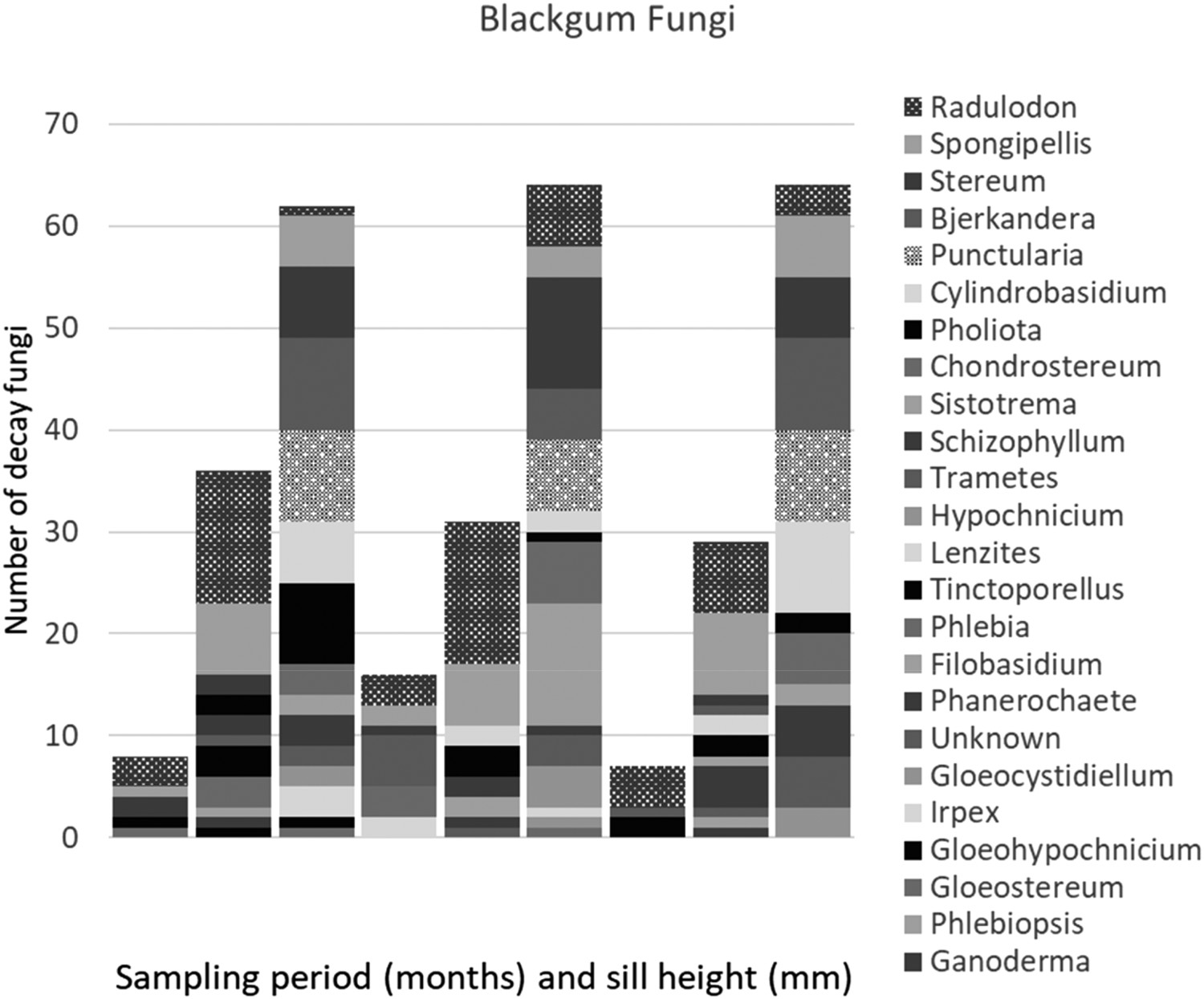
The presence of decay fungi in ties at the start of seasoning reflects a combination of incipient heart-rot fungi remaining in freshly cut ties from standing trees, as well as other fungi that may have invaded the wood in the short time between felling and sampling (Parfitt et al. 2010; Schwarze et al. 2013). These results indicate many fungi are present at the start of air-seasoning, placing added importance on the need to process ties rapidly after arrival so they can begin to dry to levels that inhibit the growth of fungi already present and limit further fungal entry.
Fungal isolations from blackgum increased markedly after three months of air-seasoning. Thirty-six fungi representing 21 taxa were isolated from stacks 150 mm off the ground, while 31 and 29 fungi were isolated from stacks placed 200 or 300 mm off the ground, respectively (Figure 3). Many of the isolates, such as Trametes versicolor, Irpex lacteus, Phanerochaete sp., Schizophyllum sp., Stereum sp., and Bjerkandera adusta are white rots commonly associated with hardwood decay. Some of these fungi have also been associated with air-seasoning railroad ties (Humphrey and Richard 1939; Duncan and Lombard 1965), decay in standing trees, (Hepting 1971; Zabel and Morrell 2020), or other timber species during air-seasoning (Sexton et al. 1992).
Air-seasoning for an additional three months further increased isolation frequency at all three stack heights. One-hundred and ninety fungi, representing 16 taxa, were isolated from gum ties after six months of seasoning. Interestingly, isolation frequencies were similar in all stacks regardless of stack distance from the ground. Ties at all three stack heights were well colonized by decay fungi.
While there were variations in taxa isolated from blackgum ties at different heights, the differences were slight. Results indicate air-seasoning gum is colonized by a wide array of decay fungi. Isolations represented increased frequencies of fungi initially present as well as several newly colonizing species. Punctularia sp, a white-rot fungus, was only isolated after six months. Some fungi initially present, such as Chondrostereum purpureum and Irpex lacteus were isolated after three months of seasoning, but not after six months. The absence of these fungi could reflect changing environmental conditions, such as MC, or competition from other fungi that limited their growth. Most taxa were present after both three and six months of seasoning. There was a noticeable increase in the frequency of Sistotrema brinkmannii, which went from single isolation after three months of seasoning to 16 isolations at six months (Figure 3). The most commonly detected fungi at most sill heights and sample times were Spongipellis delectans (a brown-rot fungus) and Radulodon americanus (a white-rot fungus), with notable increases in the white-rot fungi Sistotrema, Bjerkandera, and Stereum during the final three months (Ryvaarden 1972; Nakasone 2001). This succession may be a result of temporal changes, fungal competition, or changing wood chemistry over time (Boddy and Rayner 1983; Rayner and Boddy 1988; Boddy et al. 2007).
Blackgum was rapidly colonized by a variety of decay fungi during seasoning. Pairwise comparisons from MRPP analysis between height groups in species space showed that there were no differences between any bottom tiers, or between the bottom tiers and uppermost tiers. Pairwise comparisons of total isolate counts by MRPP did not show any significant differences between any air-dried height groups, while pairwise analysis of the final sampling period (6 months) with baseline comparisons removed did indicate evidence for increased fungal isolation with increased sill height (MRPP A: 0.10, p-value: 0.04 between 200 and 300 mm above ground), and a difference between 300 and 825 mm above ground (MRPP A: 0.13, p-value: 0.02) where 825 mm corresponded to the third row of ties on 300 mm sills. The results indicated no increased fungal activity from reduced sill height, and any fungal protection imparted by increased height was only observed in ties seasoned at least 825 mm off the ground. Sill height reduction had no significant effect on the frequency of fungal isolations in gum.
Red Oak: only four decay fungi were initially isolated from freshly cut oak ties (Figure 4). Isolations remained low at the six-month sampling point with only nine total isolates. Four fungi were isolated from the red oak ties seasoned on 150 mm sills, while two and three decay fungi were isolated from ties seasoned on 200 or 300 mm sills, respectively, at six months (Figure 3).
Fungal frequency in red oak ties air-seasoned for 12 months on German-style stacks that were 150, 200, or 300 mm off the ground. Ties were exposed in Guthrie, Kentucky and sampled at 0, 6, or 11 months after delivery. No decay fungi were isolated at the start of the test (Time 0) from ties that ended up in stacks on 150 mm sills.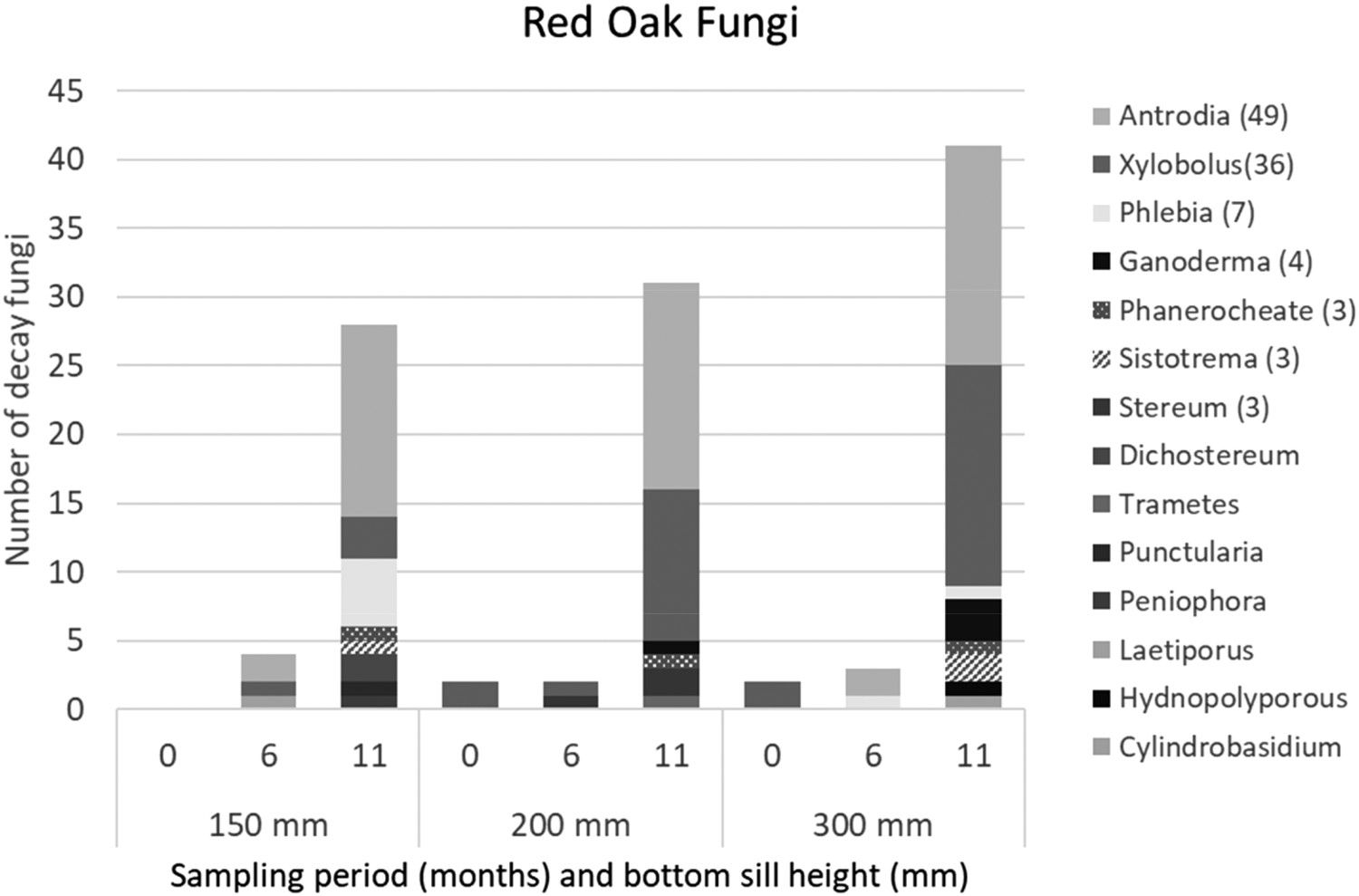
Fungal isolation frequencies increased sharply with an additional five months of exposure, although levels of fungi remained lower than in blackgum ties seasoned for only six months. Fungi in fourteen taxa were isolated 28 times from ties seasoned on 150 mm sills, with 31 and 41 isolates obtained from ties seasoned 200 and 300 mm off the ground, respectively. The much higher level of fungal isolations from ties seasoned farther above the ground was surprising and suggests the minor differences in sill heights had little effect on the ability of fungi to colonize ties in stacks closer to the ground.
Antrodia and Xylobolus spp. were the most abundant decay fungi isolated from red oak ties seasoned at all three heights, especially at the end of the air-seasoning period. Antrodia contains brown-rot species, while Xylobolus is a white-rot genus. Most other isolates were present at low levels during seasoning. While the presence of any decay fungus is important, these results suggest reducing sill height to bring stacks closer to the ground did not affect the frequency or abundance of fungal species invading stacks. An analysis of variance showed that counts of fungal isolations were significantly higher in red oak ties seasoned on 300 mm sills compared to other sill heights (p-value 0.019; d.f. 87); however, differences in the number of ties with decay fungi at each height were not significant (α=0.05), nor were the number of isolates per tie. Elevated isolations at the 300 mm sill height were likely due to an abundance of Xylobolus spp. from ties in one stack. MRPP of fungal communities grouped by height and by stack layer did not reveal fungal community differences between heights, and that community differences were slightly less heterogenous than expected by chance. Overall, results indicate that sill height had no significant effect on fungal colonization.
It is important to note that these results reflect tie seasoning in a well-maintained yard that lacked vegetation or decayed wood in the immediate area, was well gravelled, and did not have standing water. Ties were placed so that the primary wind direction facilitated drying, and the facilities closely followed the AREMA and AWPA guidelines for proper tie seasoning.
Conclusions
Railroad ties were rapidly colonized during air-seasoning, illustrating the risks of exposing wet, unprotected timber while drying. Ties were colonized by a wide array of decay fungi, illustrating the risks of exposing moist wood for long periods of time. However, moving ties closer to the soil during air-seasoning had no significant effect on the environmental conditions around the ties, the frequency with which fungi colonized the ties, or fungal community development in a properly managed seasoning yard.
Footnotes
Disclosure statement
No potential conflict of interest was reported by the author(s).