
Abstract
As biological control agents, bacteriophages can both inhibit the pathogenic bacteria and remove the bacterial biofilms from the seafood. Vibrio cholerae is the pathogen of cholera and the severe infection could lead watery diarrhea and even death. The double-layer agar plate method was used to isolate and screen the V. cholerae bacteriophages from the samples of aquaculture water and sewage. Purified bacteriophages were examined through genome sequencing, as well as morphological and biological characterizations. Among the isolated bacteriophages, bacteriophage VC3 was found to be a long-tailed bacteriophage. Whole-genome sequencing showed that the full length of VC3 genome was 27,645 bp. It was a circular dsDNA, with 40.37% G + C content. The optimal multiplicity of infection was 1, the incubation period was 20 min, the burst period was 40 min, and the lysis volume was 73 PFU/cell. Bacteriophage VC3 exhibited good activity under low temperatures and neutral pH conditions. Bacteriophage VC3 could effectively inhibit and eliminate the biofilm of V. cholerae. In addition, bacteriophage VC3 significantly inhibited V. cholerae in fish fillets and shrimp meat. At the same time, it also showed lytic activity against 9 pathogenic bacteria, indicating that it has the potential to inhibit a variety of pathogenic bacteria.
Introduction
Vibrio cholerae is the causative agent of cholera, a severe infectious disease of intestine (Kechker et al., 2017). It is a Gram-negative bacillus that is widely distributed in the water environment. The shape of V. cholerae is curved or comma-shaped and it is mobile and possesses a single flagellum and fimbriae; some V. cholerae are capsuled (Cai and Sun, 2010; Cai and Wang, 2009). V. cholerae secretes cholera toxin (CT) and severely infected individuals could be watery diarrhea. Cholera outbreaks and epidemics could lead fatal (Zhou, 2023). V. cholerae can easily grow in foods that are not processed appropriately and contamination of raw aquatic products was widely existed first (Yang et al., 2022a; Zhao et al., 2024; Han et al., 2024).
Bacterial biofilm (BF), also known as biofilm, is a spatially structured community formed by extracellular polymeric substance produced by microbial cell to surround itself. It is a common phenomenon in nature (Qu, 2018). The formation of biofilm is regulated by quorum sensing cell density-dependent gene expression, which mainly affects the formation of BF by regulating the synthesis of extracellular polysaccharides, adhesins, surfactants, and other substances (Xiao et al., 2022; Zeng et al., 2023). The cells in a biofilm aggregate are very similar and interconnected by a self-generated extracellular matrix. The biofilm represents an ecological environment related to quorum sensing, which can control the bacterial virulence and drug resistance by controlling the formation of biofilm (Lu et al., 2019; Xiao et al., 2022). Biofilms are extremely resistant to antibiotics, as well as the cleaning and sterilization processes (Yang et al., 2022c). Different methods are used to destroy the pathogenic BFs, such as antibacterial photodynamic therapy (Kesel et al., 2014), inducing biofilms detachment (Cerca et al., 2013), etc. However, these methods are not able to exert the desired effect.
Bacteriophage, a bacterial virus, is a type of virus that parasitizes bacteria, actinomycetes, cyanobacteria, spirochetes, mycoplasma, etc. (Kazi and Annapure, 2016). Bacteriophage replication depends on the host bacteria and it encodes a variety of enzymes to copy itself in the bacteria that are infected by bacteriophages. The depolymerase and lysoin encoded by the phage can degrade polysaccharides and peptidoglycans (PGs), assisting phage propagation, and cracking the BF (Topka-Bielecka et al., 2021; Zhai et al., 2021; Hao et al., 2023). Also, phage can synthesize endolysin in the host cell, directly targeting the bonds in the PG of the bacterial cell wall to specifically lyse the bacteria (Fischetti, 2018; Hao et al., 2023). The depolymerization enzyme can also recognize, bind to, and degrade polysaccharide compounds on the bacterial cell wall, destroying the structure of the bacterial cell wall, exposing the receptor, and making the phage easy to bind to its host bacterium(Latka et al., 2017). Bacteriophages preparations can be splashed or made into feed and thus phages are easy to administer into hosts and have wide application potential (Kazi and Annapure, 2016). Due to these advantages, bacteriophage therapy has gradually attracted the attention of scientists.
Cholera is a highly fatal disease with rapid onset and severe diarrhea. Seafood may be contaminated with V. cholerae during processing and transport, leading to the rampant spread of seafood-borne diseases. Bacteriophages therapy can effectively control V. cholerae contamination in seafood (Kim et al., 2024; Lasagabaster et al., 2020). At the same time, the FDA has designated phages as generally recognized as safe. To avoid cross-contamination, the FDA has allowed the use of a number of commercial phages on surfaces that come into contact with food and in food processing facilities (Kim et al., 2024; Sulakvelidze, 2013).
In the study, a V. cholerae bacteriophage was isolated from aquaculture farms by taking Litopenaeus vannamei culture water and culturing it with V. cholerae as the host bacterium using the bilayer plate method. The morphology, genome sequence, biological characteristics, and host bacterial spectrometry of isolated strain were studied. Furthermore, isolated strain was screened to have the ability to inhibit and remove V. cholerae biofilm. This study provides new method and a novel bacteriophage strain to control food pathogens.
Materials and Methods
Materials
Samples and strains
Ten replicates of seafood aquaculture water and seafood aquaculture environmental effluent were collected from the aquaculture market in Haizhou District, Lianyungang City, Jiangsu Province, China. Collected samples were quickly brought back to the laboratory and stored at 4°C. The phages were isolated from the aquaculture water of L. vannamei (shrimp) collected from the aquaculture market. V. cholerae (No. CICC 23794), Escherichia coli (No. CICC 10899), Vibrio traumaticus (No. CICC-10383), Vibrio alginolyticus (No. CICC-10475), Edwardsiella tarda (No. CICC 10630), Aeromonas hydrophilicus (No. CICC 10868), Aeromonas salmonicida, Aeromonas hydrophila (No. CICC 23934), and Pseudomonas fluorescens (No. CICC 21620) were purchased from China Industrial Culture Collection Center (CICC), Bacillus subtilis WB600 was purchased from Mall of China Beinanachuanglian Bio-technology Co, Ltd. V. parahaemolyticus (Guangdong Provincial Microbial Strain Conservation Center [GDMCC] NO.1.2391), V. harveyi (GDMCC NO.1.2066), and Aeromonas sp. (GDMCC NO.1.890) were purchased from GDMCC. Lactobacillus rhamnosus was isolated in the laboratory.
Media, reagents, and methods, see Supplementary Data.
Results
Isolation and purification of bacteriophage and genomic analysis
After multiple rounds of screening and eight purifications of strains with transparent plaques, the bacteriophage of V. cholerae was finally obtained and named VC3. The morphology of the bacteriophage spots is shown in Figure 1. The bacteriophage spots showed high transparency and had a regular shape. Under the transmission electron microscope, VC3 consisted of a head and a tail, and belonged to the bacteriophage of the family of long-tailed viruses.
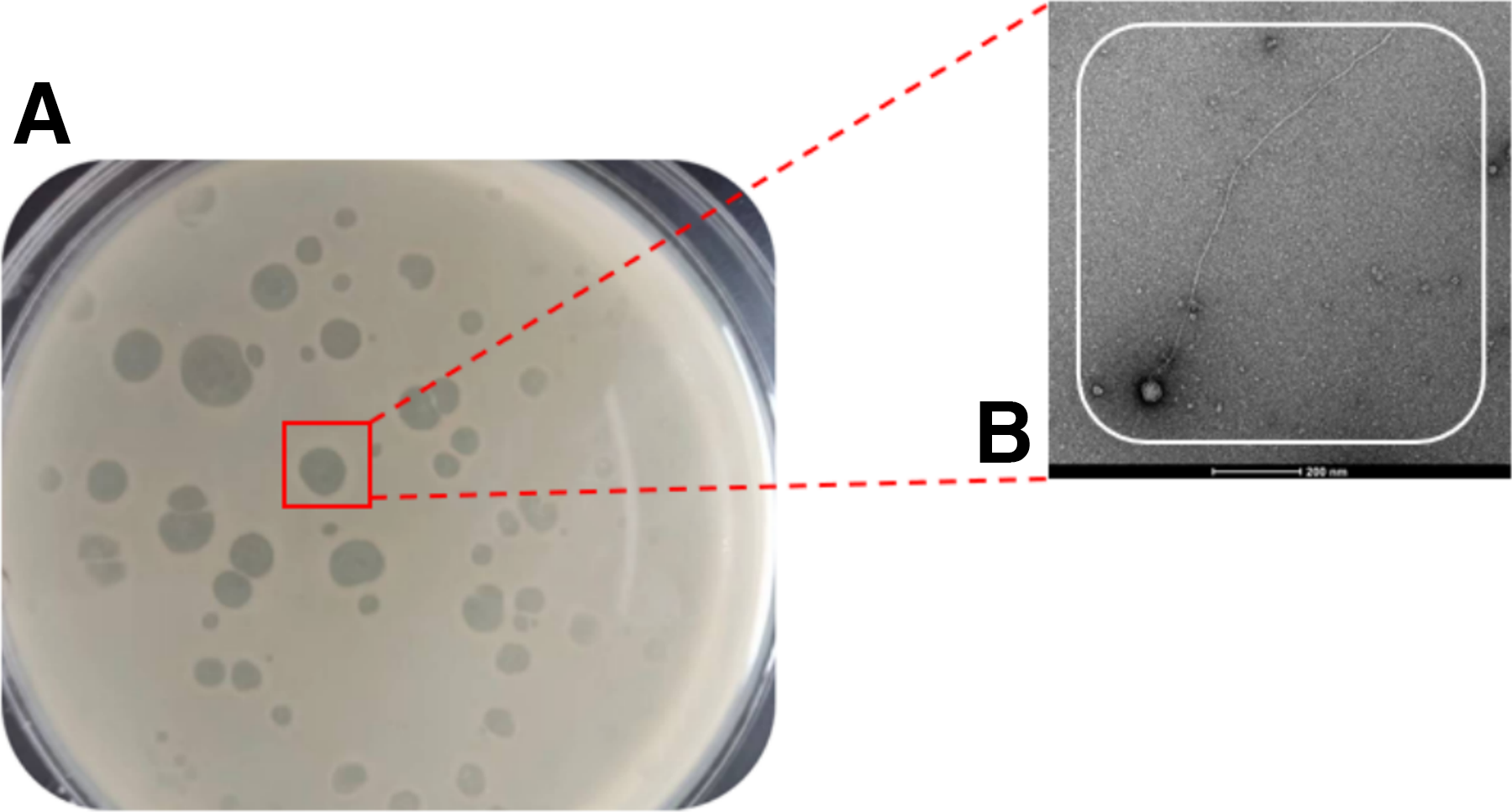
Identification of phages:
The whole genome sequence of bacteriophage VC3 was deposited in the BankIt database under accession number PP059946.The bacteriophage VC3 genome was 27645 bp in length. It was a circular dsDNA with 40.37% G + C content. The ORF prediction of the bacteriophage genome was performed by the Open Reading Frame Finder [ORFfinder Home - NCBI (nih.gov)] in NCBI with 60 ORFs. Seventy potential promoters were obtained using the BPROM program (Softberry). Meanwhile, there were 36 protein-coding genes, 30 bacteriophage hit protein genes, 6 hypothetical proteins (as in Supplementary Table S1) (PHASTER), and no tRNA genes (Pham et al., 2016), in the bacteriophage VC3 genome, which implied that the replication of VC3 was completely dependent on the host’s translation machinery (Fig. 2) (https://proksee.ca/projects/19192fd5-4f86-4a7c-93ee-9314972fc72a). Among the 30 phage hit protein genes, there were 11 protease genes, among which ORF4 was found to belong to NlpC/P60 family protein by genome sequence comparison.
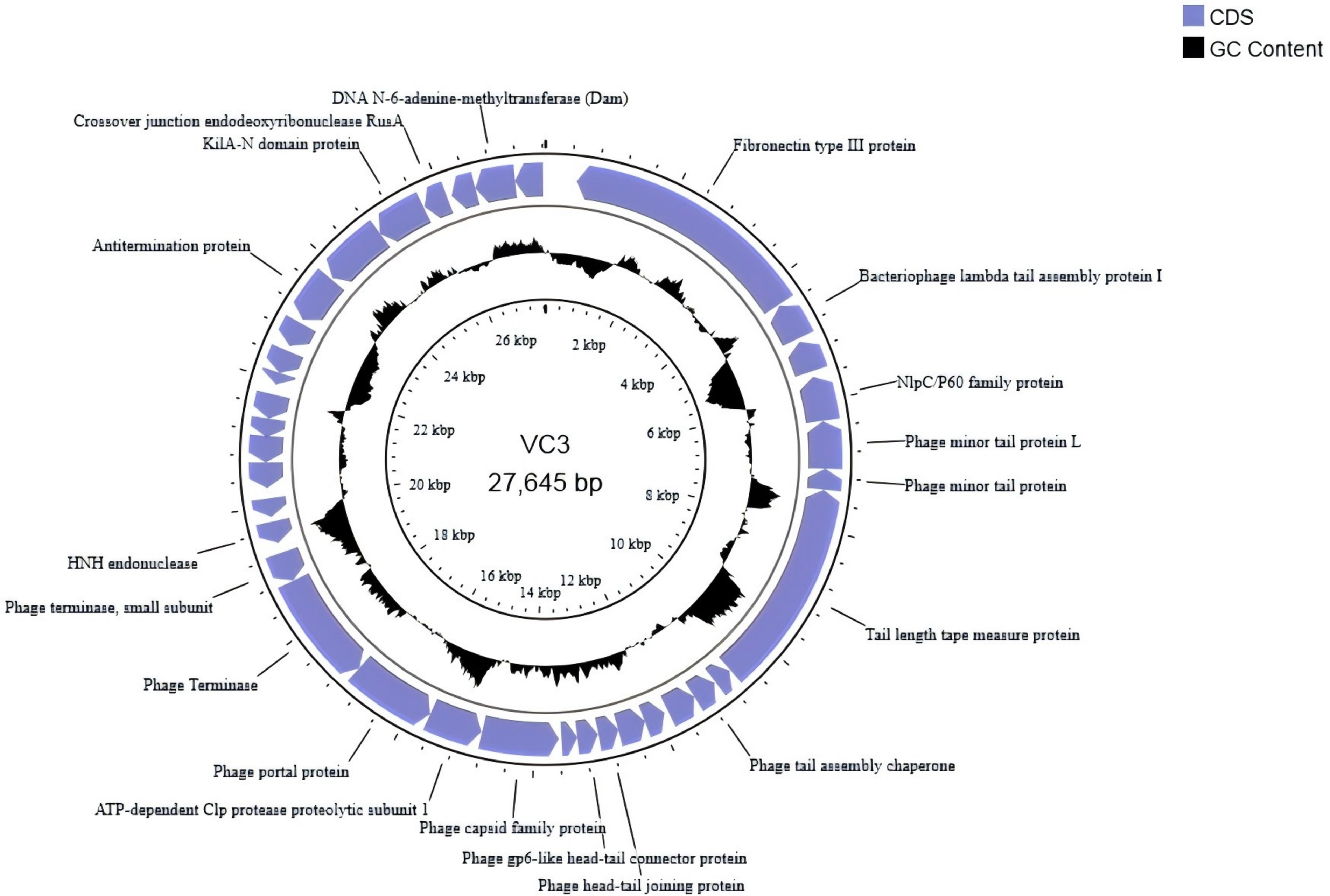
Mapping of bacteriophage VC3 gene (predicted coding regions are shown with arrows to indicate the direction of transcription and black circles to indicate G+C content).
COG classification of phage-encoded proteins showed [COGs - Clusters of Orthologous Groups (nih.gov)] (Supplementary Fig. S1) that there were 13 encoded proteins with major functions: general function prediction proteins (3); replication, gene recombination, and repair-related proteins (2); functionally unknown proteins (6); proteins related to intracellular, secretory, and vesicular transport (1); and proteins related to defense mechanism (1).
Identification of bacteriophage
Phylogenetic trees illustrate the origin and evolution of species. As shown in Supplementary Figure S2, bacteriophage VC3 and Vibrio bacteriophage phiTY18 (accession number MW451250) were located on a single branch, which suggested that phage VC3 was a phage of genus Vibrio. The homology between VC3 and phiTY18 was 45.35% (Yang et al., 2020), and phage VC3 was identified as a new strain of Vibrio bacteriophage.
Multiplicity of infection and one-step growth curve of bacteriophage
The optimal multiplicity of infection (MOI) of bacteriophage VC3 was found to be 1 (Supplementary Fig. S3A). The one-step growth curve is an important indicator of the lysis ability of bacteriophage. Supplementary Figure S3B shows that the bacteriophage had a latency period of 20 min and a burst period of 40 min. Bacteriophage lysis was calculated by dividing the titer of the bacteriophage observed at the end of lysis by the concentration of the host bacterium at the initial stage of infection (Pang et al., 2007). The lysate of phage VC3 was 73 PFU/cell.
Stability of bacteriophage
Under ultraviolet irradiation, the titer of phage VC3 gradually decreased, and the titer dropped by almost two titers after 60 min of continuous ultraviolet irradiation (Fig. 3A). Thermal stability measurement showed that the titer of phage VC3 was gradually reduced due to the influence of incubation time. When the incubation at 50°C or 70°C was 40 min, the activity of the phage decreases significantly. The phage could still maintain good activity after incubation at −80°C for 120 min, however, it completely lost activity at 50°C and 70°C for the same incubation time. (Fig. 3C). The results of acid and alkalinity resistance revealed that phage VC3 showed good activity in the pH range of 4–10 (Fig. 3B).
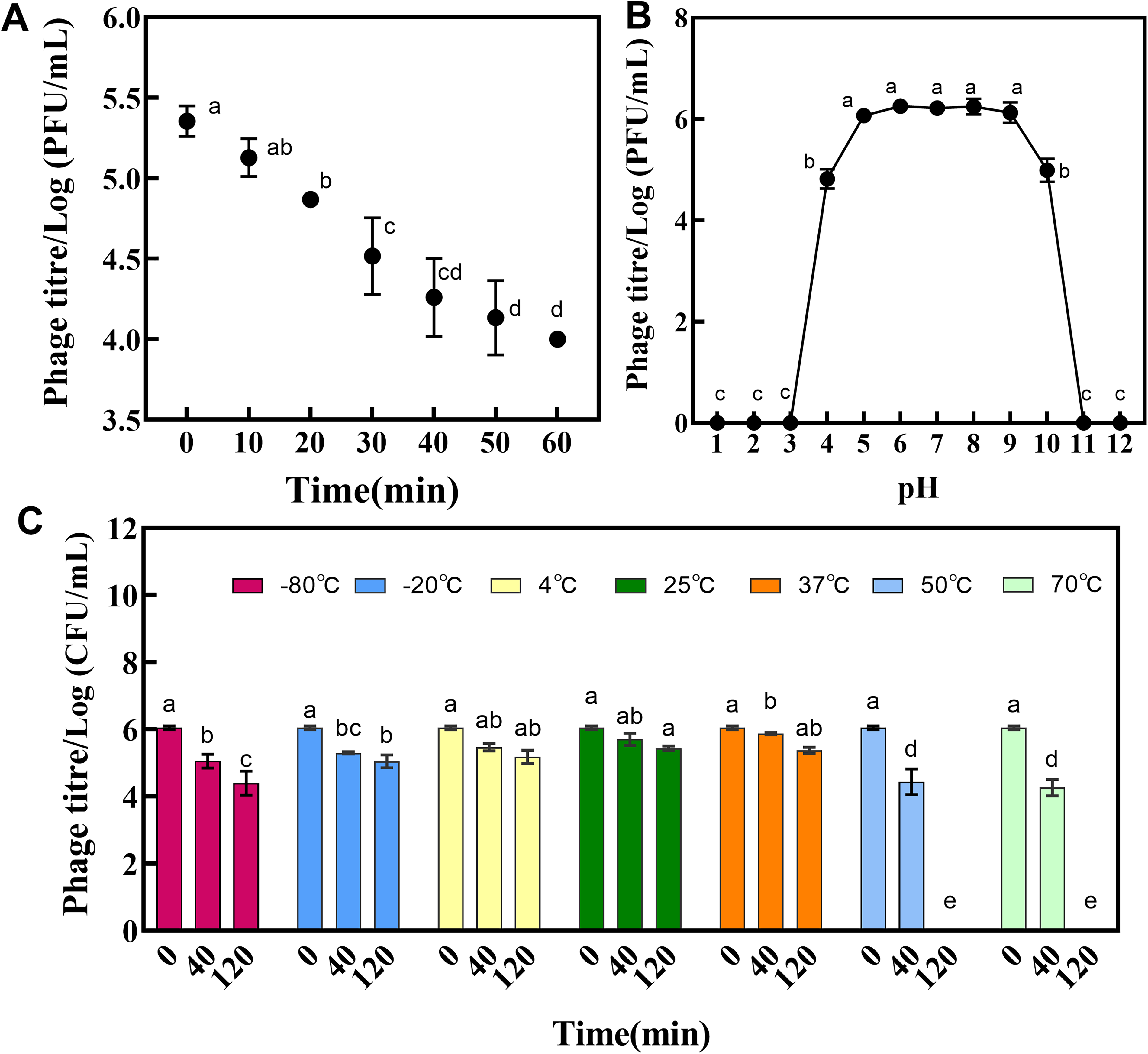
General characteristics of bacteriophage VC3:
Host bacterial spectrum of the bacteriophage
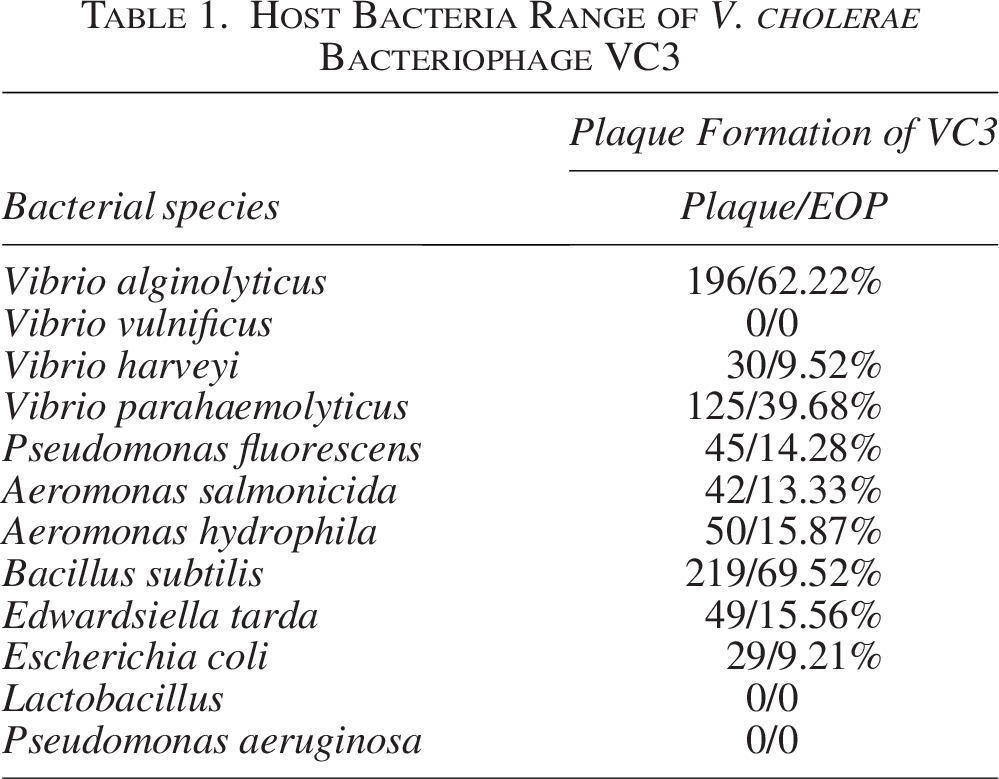
The lytic effect of phage VC3 on 12 types of bacteria is shown in Table 1. Bacteriophage VC3 had strong lysogenic effect on V. alginolyticus and B. subtilis, while its lytic effect on other bacteria was relatively weak. There was no lysis of Pseudomonas aeruginosa, Vibrio vulnificus, and Lactobacillus.
Host Bacteria Range of V. cholerae Bacteriophage VC3
Inhibitory effect of bacteriophage VC3 on V. cholerae
During the cocultivation of bacteriophage VC3 and V. cholerae, bacteriophage VC3 showed a good inhibitory effect on V. cholerae. The best inhibitory effect on V. cholerae was observed when bacteriophage VC3 was allowed to act on bacteria for 6 h. The best biofilm removal was observed when bacteriophage VC3 acted for 3 h (Fig. 4A, B).
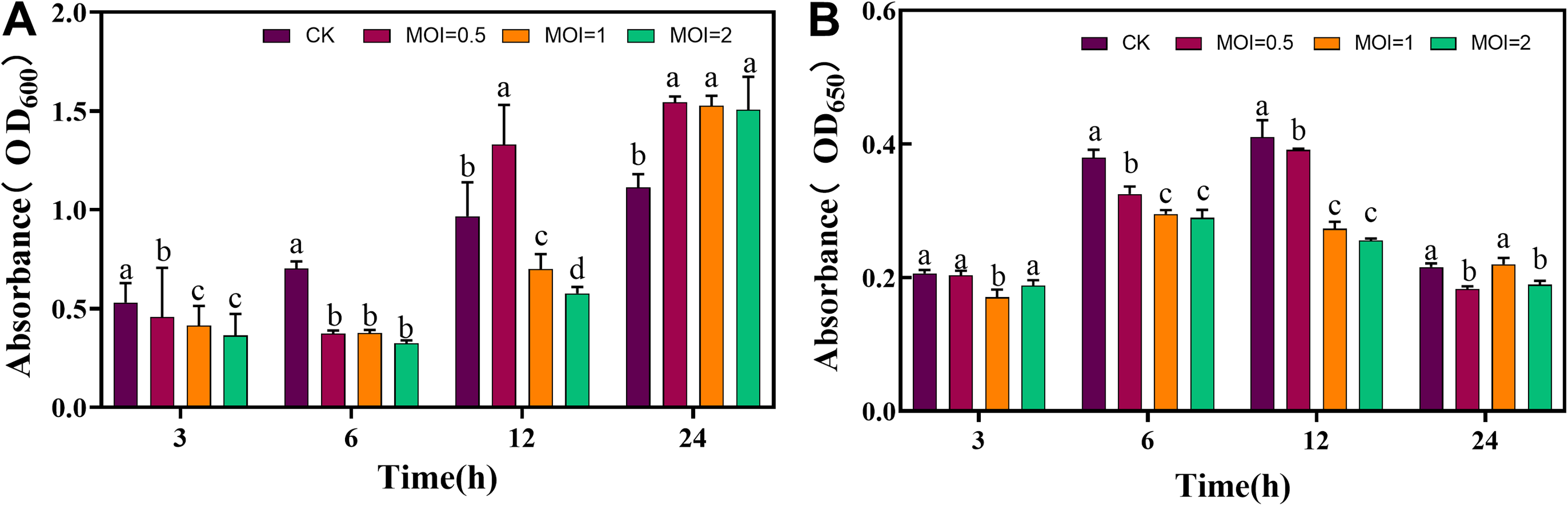
Supplementary Figure S4 shows the SEM images of co-cultivated bacteriophage VC3 and V. cholerae, which revealed the cracking of V. cholerae’s bio-periplasmic membrane. After 3 h of bacteriophage VC3 action, the bio-periplasmic membrane of V. cholerae cracked, and the bacterial growth decreased. At this point, bacteria were in a single planar shape. After 6 h of bacteriophage VC3 action, only a small amount of extracellular matrix of the biofilm existed in the image, with the residual bacterial fragments and empty housings. and the number of bacteria decreased dramatically. After 12 h of phage action, only scattered bacterial fragments were visible in the image. This comprehensive morphology analysis suggested that bacteriophage VC3 could break the periplasmic membrane of V. cholerae, and the optimal action time was 12 h.
Scavenging of established V. cholerae biofilms by bacteriophage VC3
Bacteriophage VC3 had an inhibitory effect on V. cholerae (OD600nm = 1.0) and a scavenging effect on matured biofilm (Fig. 5A, B). The results showed that the matured biofilms of V. cholerae started to decrease after 3 h of bacteriophage VC3 action, and continued to decrease till 24 h. Highest removal of V. cholerae biofilms by bacteriophage VC3 was observed at 3 h, with MOI = 2 (Fig. 5A, B).
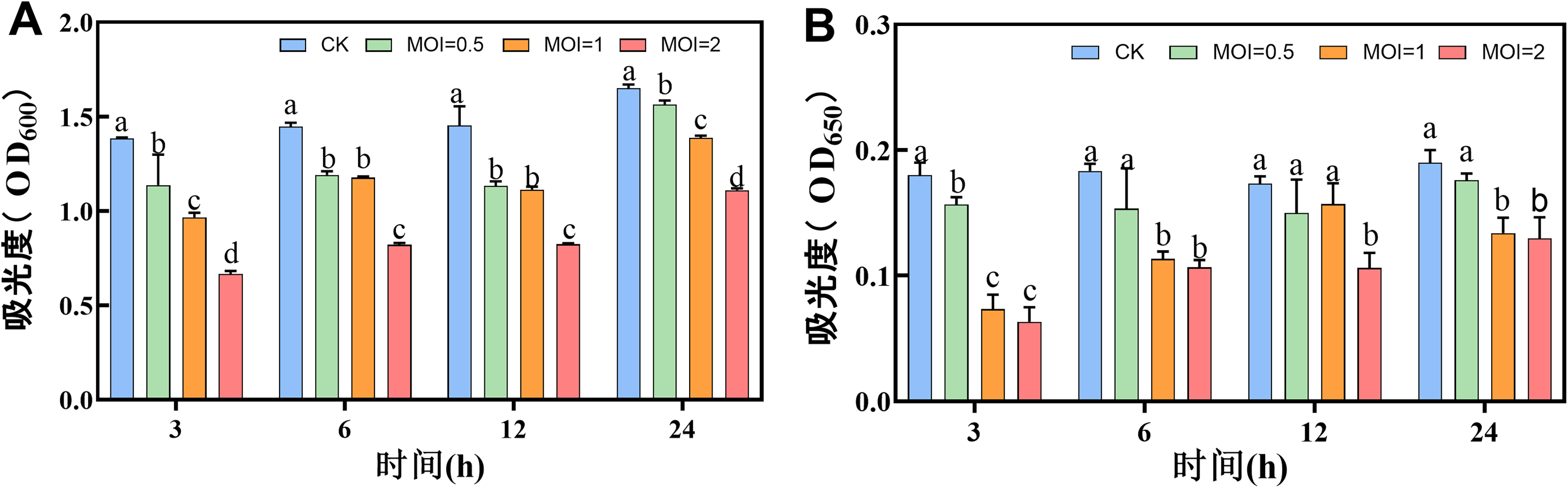
Phage VC3 action on V. cholerae
The results of scanning electron microscopy are shown in Supplementary Figure S5. Bacteriophage VC3 could destroy the biological periplasm to different degrees at different time points. A large number of overlapping and stacked bacteria were seen in the scanning electron microscope images of the control group, and the extracellular matrix could be observed between the bacterial cells. After 12 h of phage action, the extracellular matrix was hardly observed and there were only scattered bacteria. The images show that phage VC3 had a clearing effect on V. cholerae biofilm and the optimal time of action was 12 h.
Effect of bacteriophage VC3 on V. cholerae in selected seafood
The inhibitory effect of bacteriophage VC3 on V. cholerae in fish and shrimp is shown in Figure 6. The best inhibitory effect of bacteriophage VC3 on V. cholerae in fish fillets and shrimp meat was observed after 3 h of action at 25°C. At 4°C, bacteriophage VC3 had the best inhibitory effect on V. cholerae in fish fillets and shrimp meat after 4 h of interaction.
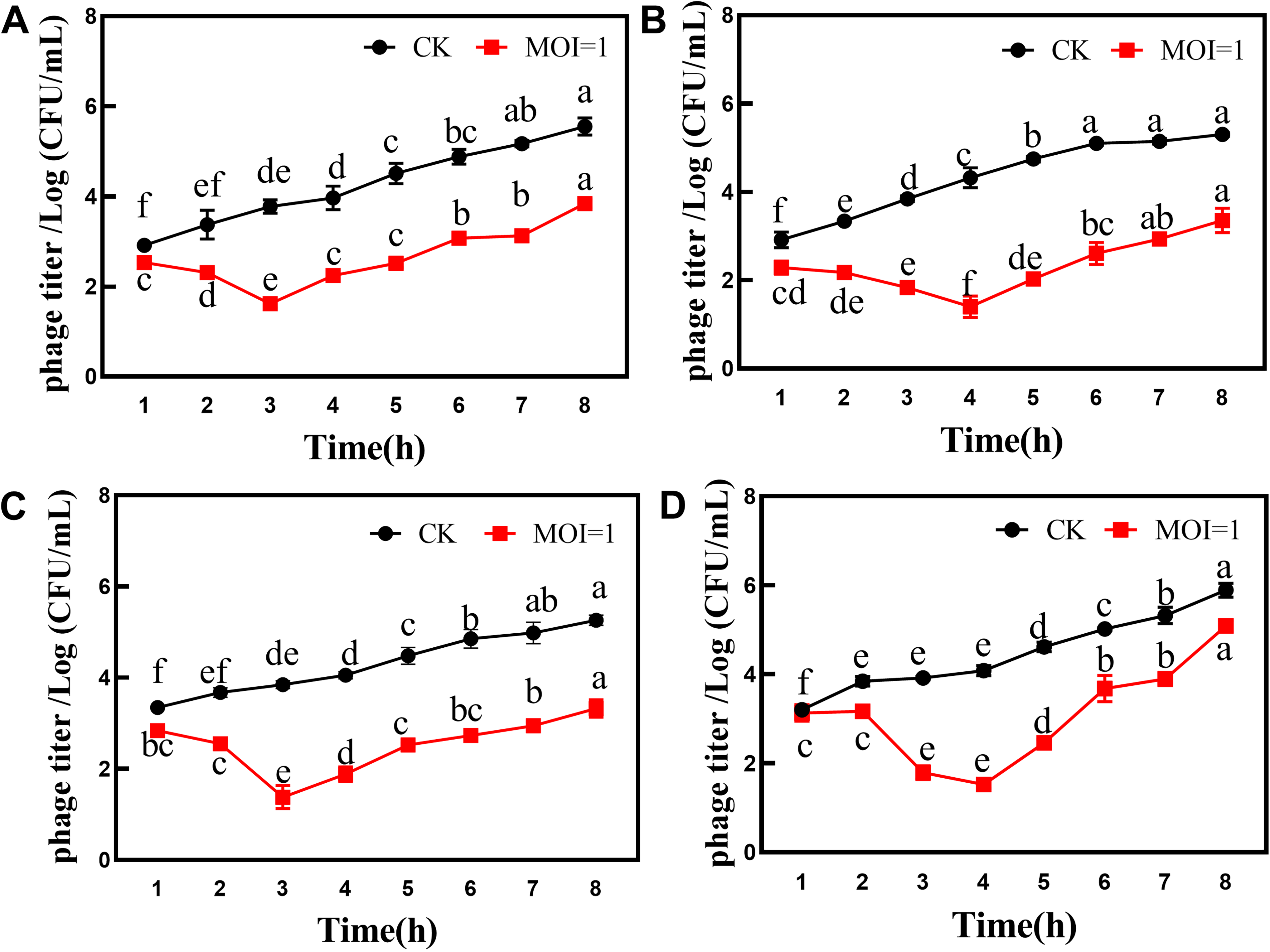
Inhibition of V. cholerae in fish and shrimp by phage VC3:
Discussion
In this study, a strain of V. cholerae bacteriophage VC3 was isolated from the culture water of L. vannamei and it was a virulent bacteriophage based on the large and transparent lysis spots. Its whole genome sequencing was compared with public databases, and the maximum sequence identity was 45.72%, so bacteriophage VC3 was a new bacteriophage strain (Adriaenssens and Brister, 2017). Meanwhile, the bacteriophage genome did not contain tRNA genes and studies have shown that tRNAs in the bacteriophage genome make up for the lack of tRNAs in the host bacterium (Bailly-Bechet et al., 2007; van den Berg et al., 2023; Dart and Ahlgren, 2023; Kim et al., 2023; Gao et al., 2023a; Namonyo et al., 2023), and that bacteriophages can improve the efficiency of translating proteins such as lytic enzymes.
MOI is one of the indicators of the effectiveness of bacteriophage lysis of pathogenic bacteria. MOI of bacteriophage VC3 was 1, which was lower than that of the bacteriophage isolated by Yan et al. (2023) and Yang et al. (2022b). The concentration of bacteriophage VC3 could be increased to investigate whether the lysis ability of bacteriophage VC3 on pathogenic bacteria could be increased.
Phage latency and lysis amount are important indicators for evaluating phage lysis ability. The one-step growth curve of phage showed that the latency of bacteriophage VC3 was 20 min and the lysis capacity was 73 PFU/cell, which was higher than that of the phage isolated by Zhang et al. (2023).
Biological characterization tests showed that bacteriophage VC3 had good adaptability at low temperatures, but is completely inactive at temperatures over 50°C for 2 h and it had a narrow pH range. Meanwhile, bacteriophage VC3 cannot adapt to strong acidic and alkaline environments. which is a limitation in practical applications (Pinheiro et al., 2019).
Bacteriophage VC3 had a broader host bacterial spectrum and it was able to play a better inhibitory role against a wide range of pathogenic bacteria in practical application. However, the limitation was lack of specificity compared with phage isolated by Yan et al. (2023) and Bhaya et al. (2011). The specificity of bacteriophage VC3 may be improved by gene editing techniques (Pastuszka et al., 2023; Shapiro et al., 2018). Also, it was special that bacteriophage VC3 had strong lysogenic effect on B. subtilis, and no effect on V. vulnificus. B. subtilis is Gram-positive bacterium and it is less common with Vibrio (Gao et al., 2023b). Meanwhile, the V. cholerae and V. vulnificus were same Vibrio. The identified targets require further research.
The results of the study showed that VC3 could inhibit and crack the biofilm, but inhibition of biofilm was more effective than cracking of biofilm by VC3. The reason may be the restriction caused by the matured periplasmic membrane, which prevented the phage from entering the cell and lyse the bacteria (Namonyo et al., 2023). On the other hand, it may also be due to the resistance of bacteria to bacteriophage, which protected the bacteria, even after the bacteriophage successfully entered the interior of the periplasmic membrane with the help of polymerase. The phage-resistant strains can reduce the inhibitory effect of phage and achieve an equilibrium symbiotic state (Qu, 2018; Yang et al., 2019).
Bacteriophage VC3 could tolerant a range of temperature and pH, and it could be an effective biological control agent for pathogenic bacteria in aquatic products. Bacteriophage VC3 was able to maintain activity at low temperatures, which is advantage for the application of bacteriophage VC3 to the control of pathogenic bacteria in seafood.
Conclusions
The bacteriophage VC3 of V. cholerae was isolated from the environmental samples, which consisted of a head and a tail, and belonged to the bacteriophage of the family of long-tailed virus and genus Vibrio. The full length of the genome was 27,645 bp. Genome consisted of a circular dsDNA with a G + C content of 40.37%. Bacteriophage VC3 had a latency period of 20 min and a lysis capacity of 73 PFU/cell. It was able to perform well at low temperatures and neutral pH. In addition, it showed lysis activity against a variety of pathogenic bacteria, and could effectively crack and inhibit V. cholerae biofilms. Bacteriophage VC3 showed significant inhibition of V. cholerae in fish fillets and shrimp meat. VC3 is a promising bacteriophage with great potential of inhibiting virulent pathogen V. cholerae in aquatic foods.
Authors’ Contributions
Q.H. designed and performed the experiments and wrote the article. Y.B., X.B., and X.K. managed software and data curation. Y.H., R.S., S.W., and L.Z. analyzed the data. M.L. and S.W. supervised the study and reviewed the article. All authors have read and approved the final article.
Footnotes
Funding Information
This study was supported by the National Key R&D Program of China (2022YFC2805101), the National Natural Science Foundation of China (32172154), the Priority Academic Program Development of Jiangsu Higher Education Institutions, and Post-graduate Research & Practice Innovation Program of Jiangsu Province (SJCX22-1658).
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Disclosure Statement
The authors confirm that there is no conflict of interests regarding this article.
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.