
Abstract
Cronobacter spp. are well known to be resistant to osmotic and dry stresses and can persist in a variety of foods. C. sakazakii and C. malonaticus are currently considered to be the primary pathogenic species that cause severe diseases. There are few studies on the prevalence of Cronobacter spp. in infant rice cereal. Information on the diversity, pathogenicity, and virulence of Cronobacter species obtained from foods is still relatively scarce and fragmentary. In this article, a total of 67 Cronobacter spp. strains were isolated from infant rice cereal in Nanchang, China. Forty-seven strains of C. sakazakii and 11 strains of C. malonaticus were identified by polymerase chain reaction (PCR). The resistance to 10 antibiotics was analyzed, showing that all isolates were sensitive except two strains. PCR detection for the five virulence genes (cpa, aut, hly, inv, and sip) and whole genome sequencing were performed. Co-existence of CSA or CMA with ampH was found in C. sakazakii and C. malonaticus strains. Compared with C. sakazakii, C. malonaticus lacked various virulence genes including yeeJ, stjC, and nanAKT genes. Another important observation was the presence of csg in the C. malonaticus genomes, while it was not found in the C. sakazakii. Notably, STs 1, 7, and 4, which are frequently associated with clinical infections, were observed. C. sakazakii and C. malonaticus strains induced higher apoptosis rate in Caco-2 cell in comparison with other Cronobacter species.
Introduction
Cronobacter species are opportunistic pathogens that can contaminate a wide range of foods. The genus Cronobacter consists of seven species: C. sakazakii, C. turicensis, C. malonaticus, C. universalis, C. muytjensii, C. condimenti, and C. dublinensis (Hu et al., 2017). There are significant differences in pathogenicity and distribution among the seven Cronobacter species. C. sakazakii and C. malonaticus are the most common Cronobacter species associated with infections. They can cause infections in all age groups but threaten infants and other susceptible individuals (Forsythe, 2018). In infants and immunocompromised individuals, C. sakazakii can cause sepsis, meningitis, and necrotizing enterocolitis (Phair et al., 2022), while C. malonaticus is considered to be linked to urosepsis and bacteremia (Li et al., 2023).
Understanding the mechanisms underlying the virulence of Cronobacter spp. is essential for the development of effective prevention and treatment strategies. It was suggested that C. sakazakii sequence type 4 (ST4) is predominantly associated with illnesses including neonatal meningitis in infants and children (Mousavi et al., 2023). Virulence factors contributing to Cronobacter pathogenicity consist of genes that function in tissue adhesion, invasion, and host cell damage (Singh et al., 2015). C. sakazakii is the only Cronobacter species possessing the nanAKT gene cassette, which encoded for exogenous sialic acid utilization and may play a role in pathogenesis (Parra-Flores et al., 2023). The known virulence factors included flagellar proteins, outer membrane proteins (ompA), hemolysins (hly), invasion (inv), plasminogen activator (cpa), autotransporter (aut), a transcriptional regulator (sdiA), siderophore-interacting protein (sip), toxin biosynthetic processing factors, etc. (Mousavi et al., 2023). Some authors reported that the cpa gene, which is related to serum resistance and invasion, was not found in the C. malonaticus strain (Parra-Flores et al., 2023). It has been shown that curli biosynthesis gene csg is found in C. dublinensis, C. malonaticus, C. turicensis, and C. universalis but not in C. sakazakii and C. muytjensii (Jang et al., 2020). To date, the pathogenesis and virulence factors of Cronobacter spp. have not been fully elucidated and require further investigation. Information on the diversity, pathogenicity, and virulence of C. malonaticus isolated from various sources remains limited.
Cronobacter species exhibit unusual resistance to heat, dry, and acid stress growth conditions, so that they can survive for extended periods in various processing environments, including flour, spices, and dairy products (Jaradat et al., 2014). In recent years, dairy companies have frequently recalled products contaminated with C. sakazakii. In February 2022, Abbott voluntarily recalled powdered infant formula (PIF), and in March 2023, Nestle recalled PIF because of contamination with C. sakazakii (Schultz et al., 2023). Monitoring Cronobacter spp. isolates in infant foods is important (Abebe, 2020).
Some Chinese authors studied the prevalence and antimicrobial resistance (AMR) phenotype of Cronobacter cultured from PIF and supplementary food (Gan et al., 2022). A total of 1055 Cronobacter spp. isolates were recovered from 12,105 samples of PIF and supplementary food collected from 29 provinces between 2018 and 2019 in China (Gan et al., 2022). Among these, 1048 isolates were from infant rice cereal, and 7 were from PIF. The infant rice cereal showed a relatively high detection rate compared with PIF (Gan et al., 2022). Infant rice cereals are the most popular first complementary food for baby in China. However, information on Cronobacter spp. contamination in infant rice cereal in China is still limited.
C. sakazakii has shown resistance to various antibiotics, as well as the presence of antibiotic resistance genes, plasmids, and mobile genetic elements (Holýet al., 2023). However, limited data are available on the antibiotic resistance profiles and genomic characteristics of C. malonaticus in infant foods. C. sakazakii and C. malonaticus are close and indistinguishable based on the 16S rDNA sequence analysis (Joseph et al., 2012). Therefore, the aim of this study was to isolate Cronobacter spp. from infant rice cereal in NanChang, China, to identify C. sakazakii and C. malonaticus strains and to verify the antibiotic susceptibility, virulence characteristics, and genomic specificity of the representative strains by detecting virulence genes, whole-genome sequencing (WGS), and apoptosis induction of Caco-2 cell.
Materials and Methods
Isolation of Cronobacter spp. strains
A total of 67 Cronobacter spp. strains were isolated from infant rice cereal samples from 2018 to 2023 according to the Chinese standard GB4789.40—2016. A pre-enrichment with 100 g of samples and 900 mL buffered peptone water (Qingdao Hope Bio-Technology Co., Ltd., China) was incubated at 37°C for 18 h. Afterward, 1 mL aliquot of pre-enrichment samples was transferred to 10 mL of modified lauryl sulfate tryptose broth-vancomycin medium (mLST-Vm) and incubated at 44°C for 24 h. A loop of enriched mLST-Vm culture was streaked onto modified C. sakazakii chromogenic medium (CHROM Agar, Qingdao Hope Bio-Technology Co., Ltd., China) and incubated at 37 °C for 24 h. Typical colonies were selected (two to five colonies with blue in color) and transferred to tryptic soy agar (Qingdao Hope Bio-Technology Co., Ltd., China) incubated at 25°C for 48 h. After incubation, the colonies that changed color to yellow were purified, prepared glycerol stocks, and stored at −20°C.
Identification of Cronobacter spp. strains
For genus identification, the suspected strains were checked by using Cronobacter Genus Biochemical Identification Kit (Beijing Land Bridge Technology co., Ltd.). The total DNA of positive strains was then extracted with Bacterial Genomic Extraction Kit (TianGen Biotech Co., Ltd., China) and stored at −20°C until use. The confirmation of presumptive Cronobacter spp. strains was further achieved using polymerase chain reaction (PCR) based on 16SrDNA. For species identification, the gyrB of C. sakazakii and the group_32895 of C. malonaticus were used as target genes to identify the two species based on the reported species-specific primers(Huang et al., 2013; Shang et al., 2021). The nanA of C. sakazakii and the mmcR of C. malonaticus were also used as target genes to screen the two species (Supplementary Table S1). All of the PCR reactions were conducted using a 25 μL system. The mixture included 0.15 μL rTaqTM DNA olymerase (Takara, Japan), 2.5 μL 10×Easy Taq Buffer, 2 μL dNTP (2.5 mM), 1 μL of each primer (10 μmol/L), 1 μL of bacterial DNA template, and distilled water to the final volume of 25 μL. The PCR primers were listed in Table 1, and the PCR reactions were performed as described by the references. The type strains C. sakazakii ATCC51329 and C. malonaticus CICC24177 were used as positive controls. The strains that were positive for the genus identification and negative for the species identification were named as Cronobacter genus strains.
T
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed using a broth microdilution method with the Mueller–Hinton medium in 96-well culture plates according to the 2022 Clinical and Laboratory Standards Institute guidance (CLSI, 2022). The 10 antibiotic drugs were assessed: ampicillin, ciprofloxacin, ceftriaxone, cefotaxime, polymyxin B, nitrofurantoin, tetracycline, rifampicin, imipenem, and gentamicin. Antibiotic drugs were purchased from Shanghai Yuanye Bio-Technology Co., Ltd. Escherichia coli ATCC25922 was used as the quality control strain. The minimum inhibitory concentration results were interpreted as susceptible, intermediate, or resistant based on the 2022 Clinical and Laboratory Standards Institute breakpoints.
Detection of virulence genes
The five virulence genes (cpa, aut, hly, inv, and sip) were screened by PCR as previously described (Holý et al., 2019). The PCR mixture was the same as the methodology described above. The PCR primers and reaction conditions were listed in Supplementary Table S1. All PCR reactions were performed in duplicate.
Whole-genome sequencing
A total of 13 C. sakazakii and C. malonaticus strains with the most and the least virulence genes, as well as those that are drug-resistant and show intermediate resistance, were selected for WGS sequencing. In 10 strains, second-generation sequencing was performed, and 3 strains were subjected to third-generation sequencing because of cost consideration.
The draft genome sequence analysis of 10 strains was carried out using the Illumina NovaSeq PE150 (Novogene Co. Ltd., Tianjin, China). Sequencing libraries were generated using NEBNext®Ultra™ DNA Library Prep Kit (NEB, USA). Raw sequence reads were assembled with SOAP denovo 2.04, SPAdes 3.13.0, Abyss, and GapCloser 1.12.
Three strains were subjected to Shanghai Majorbio Bio-Pharm Technology Co., Ltd., to perform third-generation sequencing by using the Nanopore and the Illumina HiSeq X-10 platforms. Illumina sequencing libraries were prepared from the sheared DNA fragments using the NEXTFLEX Rapid DNA-Seq Kit. The Nanopore library was prepared and sequenced on Oxford Nanopore PromethION (Oxford Nanopore, Oxford, UK). Raw sequence reads were de novo assembled with Unicycler v0.4.8, SPAdes 3.13.0, and GapCloser 1.1.
Gene prediction was performed using Glimmer version 3.02, RAST2.0, and GeneMarkS version 4.3. AMR genes were identified using ResFinder 4.1 and CARD (https://card.mcmaster.ca/). Virulence genes were predicted using VFDB (http://www.mgc.ac.cn/VFs/main.htm) and TADB (http://bioinfo-mml.sjtu.edu.cn/TADB/). MLST typing was performed using PubMLST (http://www.pubmlst.org/Cronobacter/), and the results were listed in Table 2.
T
The apoptosis of Caco-2 induced by Cronobacter spp.
Human Caco-2 cells were purchased from BOSTER Biological Technology Co. Ltd. (Wuhan, China) and cultured in DMEM with 20% fetal bovine serum and 1% streptomycin/penicillin. All cells were grown at 37°C in 5% CO2 incubator. A total of 16 Cronobacter spp. strains including 13 sequenced strains and 3 Cronobacter genus strains were cultured at 37°C under shaking in LB broth until culture OD600 reached about 0.5. Then, 10 mL of bacterial enrichment cultures was centrifuged at 9000 rpm for 10 min. The supernatants were filtered through a membrane filter (pore size, 0.22 μm) and then used for apoptosis induction. The concentration of Caco-2 cells was adjusted to 105 cells/mL by fresh DMEM. Each well of the 6-well cell culture plate was added with 1 mL DMEM and 1 mL diluted Caco-2 suspension, and incubated in a CO2 incubator for up to 48 h. On the third day, the medium was discarded, and cells were washed with phosphate buffered saline (PBS) solution. Then, 2 mL fresh medium and 150 μL of bacterial filtrate samples were added to 6-well plates. After incubation with bacterial supernatant for periods of up to 24 h, cell monolayers were detached with trypsin, washed in PBS, and collected by centrifugation at 1000 rpm for 5 min. Cells were resuspended in 100 μL 1× binding buffer and incubated for 15 min in the dark with 5 μL Annexin V-FITC and 5 μL propidium iodide (PI) to monitor apoptosis. The Annexin V-FITC/propidium iodide kit was purchased from Beilunbio Co. Ltd. (Dalian, China). The cell apoptosis was quantified by flow cytometry (FongCyte™, Chanllenbio Co. Ltd., Beijing, China).
Results
Identification of Cronobacter genus and species
Cronobacter spp. strains were isolated from infant rice cereal samples, and the identification of Cronobacter genus was performed by commercial biochemical identification kit and PCR targeting the 16SrDNA gene. A total of 67 strains were positive for the both methods and finally identified as Cronobacter genus. Species identification of these strains was performed with an initial screening by the PCR for the gyrB and nanA of C. sakazakii and the group_32895 and mmcR of C. malonaticus. Among the 67 Cronobacter strains, 46 were positive for both gyrB and nanA, and 11 were positive for both group_32895 and mmcR via PCR. There was difference in the identification of C. sakazakii isolates by PCR. One strain J28 was identified as C. sakazakii by nanA PCR and WGS, but it was not positive for gyrB PCR.
Out of the 67 Cronobacter spp. strains, 47 strains (70.15%) were finally classified as C. sakazakii, and 11 strains were classified as C. malonaticus (16.41%). Nine strains were classified as Cronobacter genus and belonged to the other Cronobacter species.
The profile of antimicrobial susceptibility testing
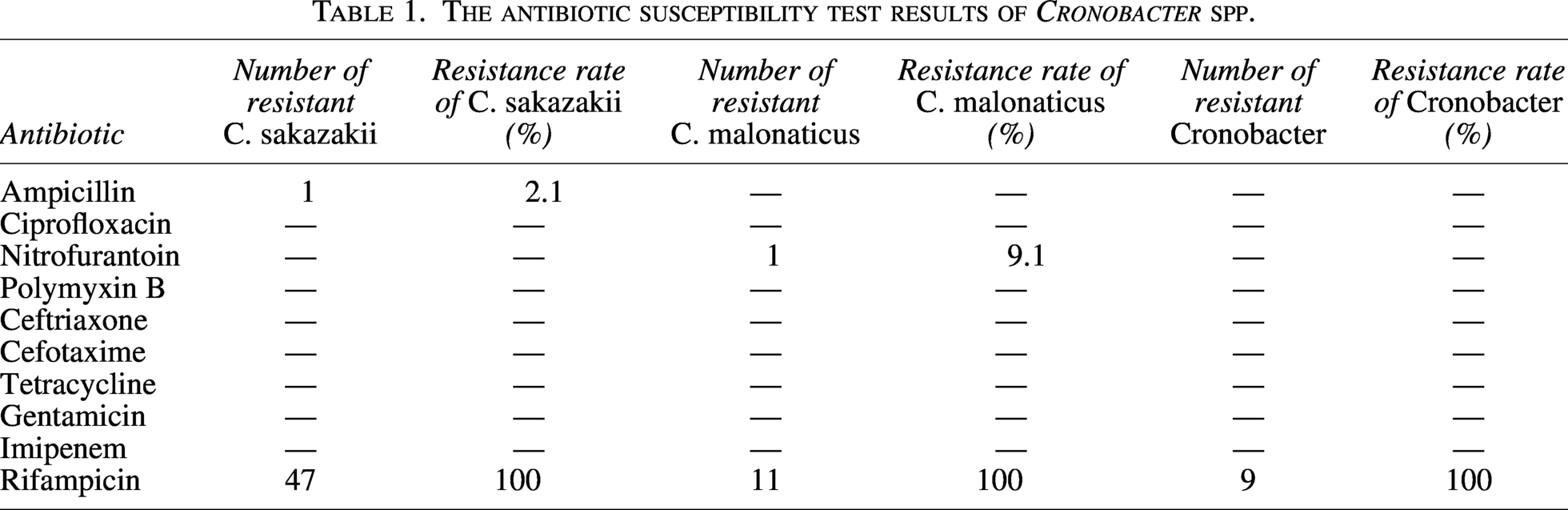
We selected 10 antibiotics from 9 groups to examine the susceptibility, intermediate resistance, and resistance profiles of 67 Cronobacter spp. isolates (Table 1). All the strains were resistant to rifampicin. However, they were all sensitive to ciprofloxacin, polymyxin B, tetracycline, gentamicin, and imipenem. One C. sakazakii isolate (J28) was resistant to ampicillin. One C. malonaticus isolate (J41) was resistant to nitrofurantoin. The nine Cronobacter genus isolates were sensitive to all antibiotics except rifampicin.
PCR detection of virulence genes in Cronobacter spp. strains
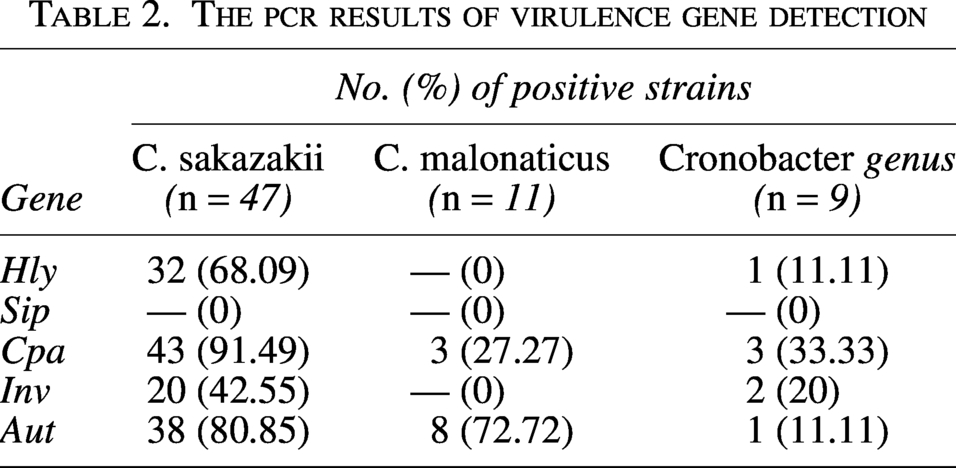
The 67 Cronobacter spp. isolates showed differences in the presence of virulence genes, as shown in Table 2. Regarding the species, the cpa gene was the most frequently detected in C. sakazakii isolates (43/47, 91.49%) followed by the aut gene (38/47, 80.85%), the hly gene (32/47, 68.09%), and the inv gene (20/47, 42.55%).
Eight strains of C. malonaticus were positive for aut gene (72.72%), and three strains were positive for cpa gene (27.27%). The virulence genes were detected in other Cronobacter species as follows: cpa (3/9, 33.33%), inv (2/9, 22.22%), aut (1/9, 11.11%), and hly (1/9, 11.11%). The gene sip has not been found in any species of the Cronobacter spp. isolates (Table 2).
Most of the C. sakazakii isolates carried multiple virulence genes. For example, 15 C. sakazakii isolates carried hly, cpa, inv, and aut genes (31.91%), simultaneously. The proportion of C. sakazakii carrying more than two virulence genes was as high as 91.49%, and only one strain of C. Sakazakii carried none of any test virulence genes, with a detection rate of 2.12%. However, C. malonaticus carried fewer virulence genes. Only two C. malonaticus isolates (18.18%) carried two virulence genes (cpa and aut). The sporadic distribution of the virulence genes was observed in the nine Cronobacter genus strains, but none of the strains carried more than two virulence genes.
WGS analysis
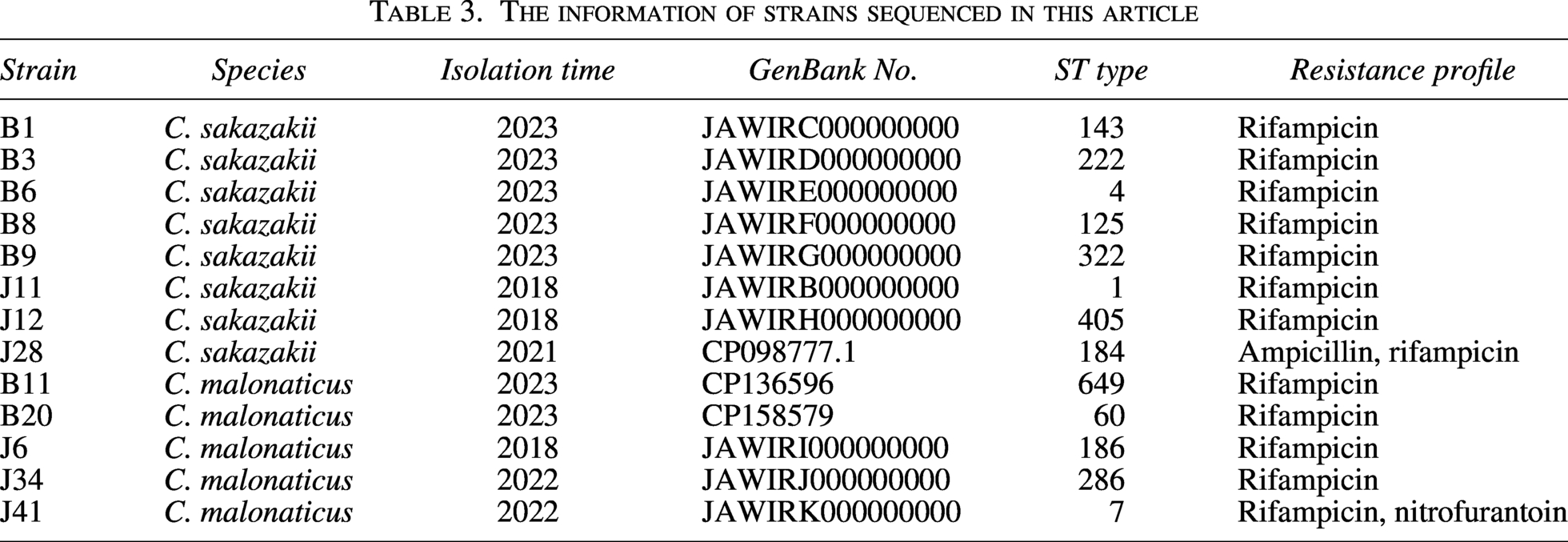
Initial experiments using PCR had identified 58 Cronobacter spp. isolates as either C. sakazakii or C. malonaticus. Due to cost consideration, 13 strains including 8 C. sakazakii and 5 C. malonaticus strains were selected to perform WGS. The sequences of the seven housekeeping genes (atpD, fusA, glnS, gltB, gyrB, infB, and ppsA) were extracted and compared with the Cronobacter MLST database. The 13 strains were clustered into 13 STs, shown in Table 3. Notably, C. sakazakii strain B6 was assigned to ST4. C. sakazakii strain J11 and C. malonaticus strain J41 were assigned to ST1 and ST7, respectively.
T
C. sakazakii J28 was resistant to ampicillin. WGS data showed that it contained ampH and ampCSA-1. The β-lactamase gene ampH was present in all sequenced C. sakazakii and C. malonaticus strains (Fig. 1A). Co-existence of the variants of β-lactamase resistance genes blaCSA in C. sakazakii and blaCMA in C. malonaticus with ampH was found. All the C. sakazakii and C. malonaticus strains exhibited the same efflux genes (emrR, emrB, marA, marR, rsmA, CRP, adeF, acrA, kpnE, kpnF, qacG, and msbA), antibiotic target alteration genes (glpT, EF-Tu, vanG, PBP3, msrA/B, and srmA), and fosfomycin resistance gene fosA. The C. sakazakii strain J11 carried mcr-9.1, which confers resistance to colistin (Fig. 1A). However, it showed susceptibility to polymyxin B (Table 3).
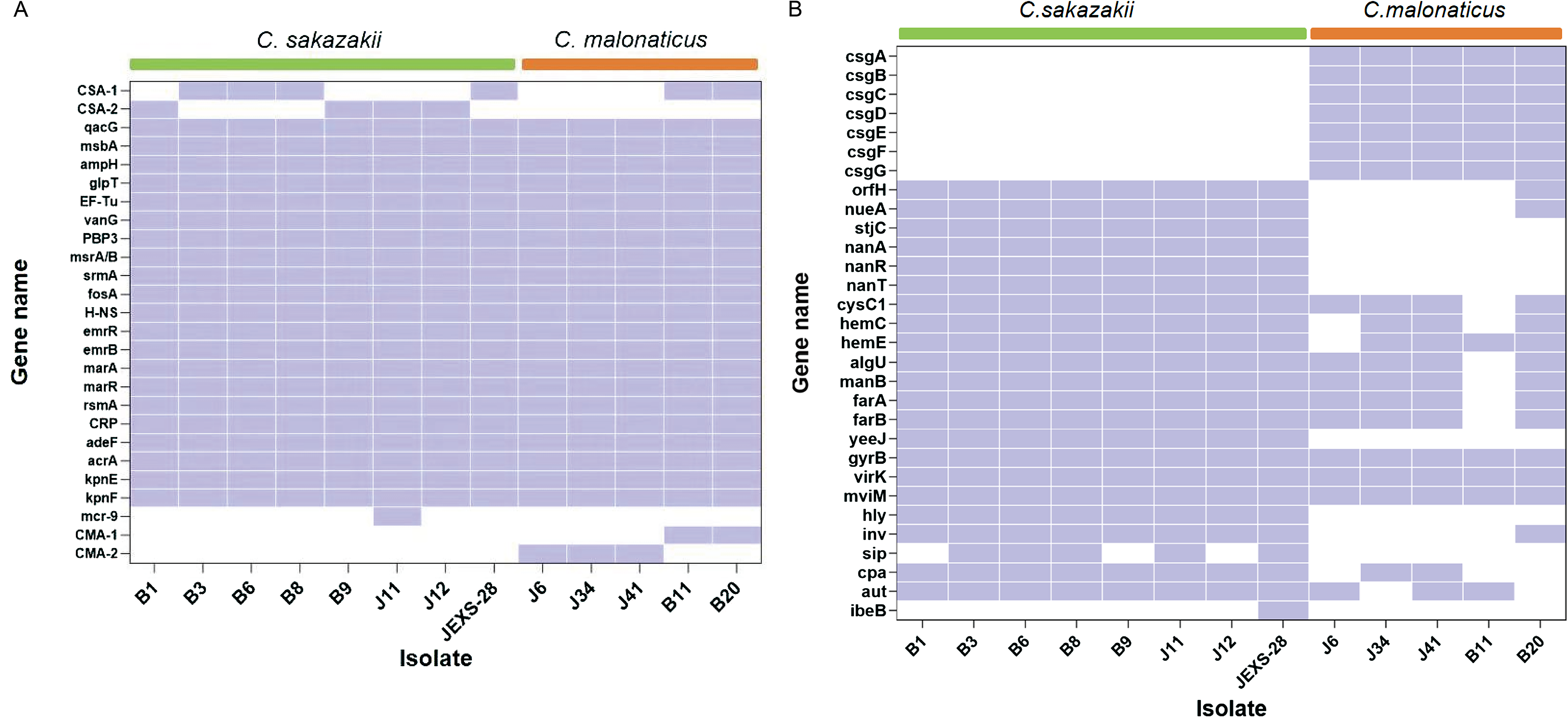
Comparison of differential resistance and virulence genes between C. sakazakii and C. malonaticus.
The nanART gene cluster was specific to C. sakazakii strains (Fig. 1B). The virulence genes including stjC and yeeJ were also presented in C. sakazakii strains; however, these genes were not identified in C. malonaticus (Fig. 1B). The curli assembly/transport genes csgA/B/C/D/E/F/G were present in all C. malonaticus strains, while it was not found in C. sakazakii. The ibeB gene that was probably associated with the invasion of human brain microvascular endothelial cells (HBMECs) was only predicted in C. sakazakii J11 (Fig. 1B).
The apoptosis induction
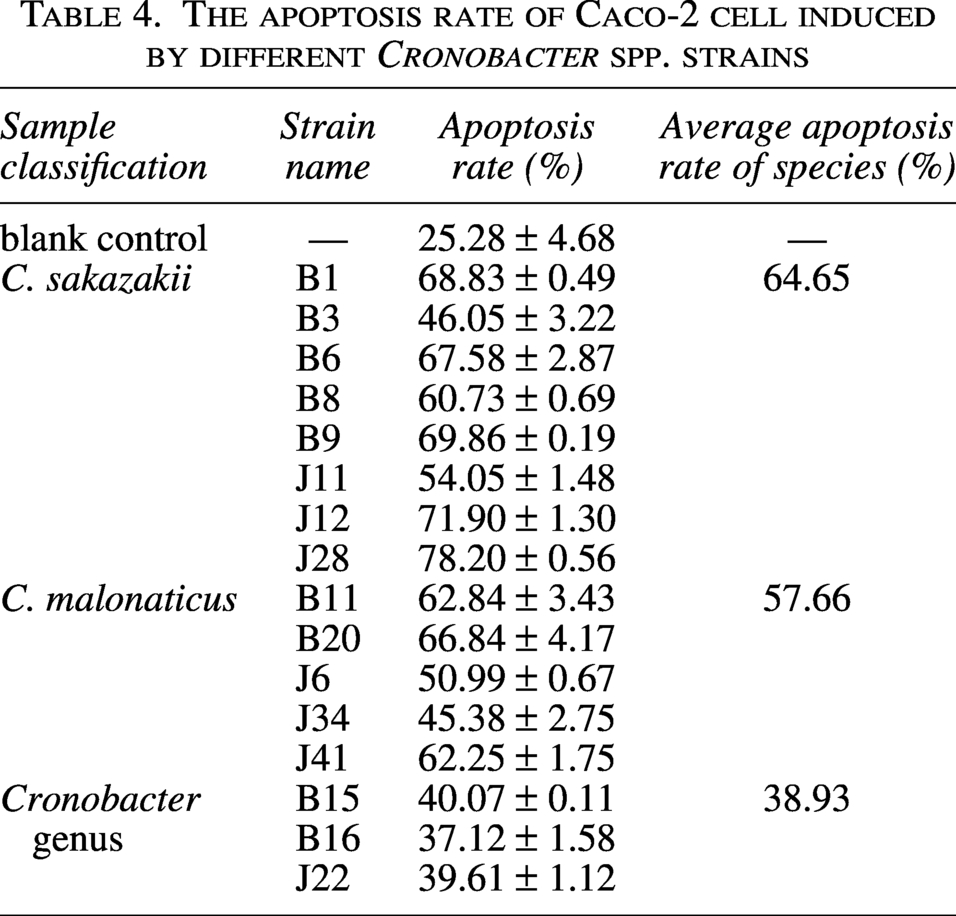
Eight C. sakazakii, five C. malonaticus, and three Cronobacter genus strains infected Caco-2 cells for 24 h. The apoptotic Caco-2 cells were stained with Annexin V/FITC and PI and analyzed using a flow cytometer.
The eight C. sakazakii and five C. malonaticus strains induced high levels of apoptosis (between 46.05% and 78.20% for C. sakazakii and 45.38% and 66.84% for C. malonaticus) in Caco-2 cells. The three Cronobacter genus strains caused lower levels of apoptosis (between 37.12% and 40.07%) in Caco-2 cells.
Discussion
At present, fast detection and identification of Cronobacter isolates to the species level by PCR-based method remains a complex task. Several target genes, such as 16S rDNA, rpoB (Vlach et al., 2017), fusA (Li et al., 2017), ompA (Yan and Fanning, 2015), cgcA (Silva et al., 2019), gyrB (Huang et al., 2013), etc., have been used for the differentiation of Cronobacter at the species level. However, the reliable specific marker genes for identification of Cronobacter species are still deficient. C. sakazakii and C. malonaticus could be detected at both the genus and species level. However, the other species of Cronobacter could be detected accurately only at the genus level (Wang et al., 2022). Most infections are caused by C. sakazakii and C. malonaticus. This article only identified C. sakazakii and C. malonaticus at species level because of the limitation of current methods and the importance of different Cronobacter species. Among 67 Cronobacter strains, 47 (70.15%) C. sakazakii strains and 11 (16.41%) C. malonaticus strains were finally identified in our article. Li et al. reported the similar results (Li et al., 2023a). They isolated 136 Cronobacter spp. strains from wet rice and flour products in Guangdong province (Li et al., 2023a). They found that C. sakazakii accounted for 52.21% (71/136) of the total Cronobacter spp. population, while C. malonaticus accounted for 41.91% (57/136).
C. sakazakii is intrinsically resistant to rifampicin (Stock and Wiedemann, 2002). In our article, Cronobacter showed susceptibility to most antibiotics except for rifampicin. Li et al. studied 154 Cronobacter spp. isolates and found that all strains were sensitive to cefepime, tobramycin, ciprofloxacin, and imipenem, in which the resistance to cephalothin was the highest (64.94%) (Li et al., 2023b) . Gan et al. isolated 1055 Cronobacter spp. strains from 12,105 samples of PIF and supplementary food collected from 29 provinces between 2018 and 2019 in China (Gan et al., 2022). Only 11 (1.0%) isolates were resistant. Though Cronobacter spp. isolates are generally susceptible to the antimicrobial agents and multidrug resistance Cronobacter spp. isolates were seldom identified, in recent years the antibiotic resistance of C. sakazakii has increased owing to the acquisition of resistance genes (Shi et al., 2018).
The production of chromosomal AmpC b-lactamases, including CSA-1 and CSA-2 in C. sakazakii, and CMA-1 and CMA-2 in C. malonaticus, confers the resistance exclusively to the first-generation cephalosporins. The blaCSA and blaCMA genes were first described in 2014 and are associated with β-lactamase class C resistance; they are not inducible and are regarded as cephalosporinases (Müller et al., 2014). In our article, the C. sakazakii and C. malonaticus strains co-harbored CSA or CMA with β-lactamase ampH. In addition to it, all sequenced C. sakazakii and C. malonaticus strains carried fosA gene. To the best of our knowledge, this finding has not been reported previously. Parra-Flores et al. isolated four C. sakazakii and one C. malonaticus strains from different PIF batches from Chile (Parra-Flores et al., 2022). All the C. sakazakii exhibited blaCSA-1, and the C. malonaticus strain showed blaCMA-1. The co-existence of ampH was also observed. However, fosA was not found in the Cronobacter spp. isolates (Parra-Flores et al., 2022).
C. sakazakii strain J11 carried mcr-9.1. However, it showed susceptibility to polymyxin B. It has been reported that the expression of mcr-9 gene requires promoter, inducer, etc. Although some bacteria carry this gene, they lack related factors required for gene expression, so they cannot show resistance to polymyxin (Carroll et al., 2019).
This article is focused on the comparison of virulence genes between C. sakazakii and C. malonaticus. In our article, PCR screening of five virulence genes showed that none of the C. malonaticus strains harbored the hly and inv gene, but they were present in some C. sakazakii strains tested. The gene sip has not been found in any strains. The finding was quite different from Holy’s report (Holý et al., 2019). They analyzed 25 strains of C. sakazakii and 11 strains of C. malonaticus isolated from different clinical materials. They found that sip was detected from most samples (97%, 35/36). Another notable aspect was the presence of the hly gene in 100% of C. malonaticus strains and the inv gene in 54.44% of C. malonaticus strains (Holý et al., 2019). The difference of property should be related to separation source of strains.
Compared with C. Sakazakii, C. malonaticus lacked some genes that play a role in the stage of adhesion and invasion of host cells, such as virulence factor yeeJ and stjC (Fig. 1B). Plasminogen activator gene cpa has been demonstrated to confer resistance to bactericidal serum and facilitate invasion (Chen et al., 2023). It was found in most of C. sakazakii strains and a few C. malonaticus strains by PCR (Table 2). The PCR detection rate of inv gene was 42.55% in C. sakazakii (20/47) and 22.22% in C. malonaticus (2/9) (Table 2). Studies have shown that inv not only promotes biofilm formation but also contains an LysM domain that interacts with peptidoglycan to help it locate in the outer membrane (Martinez-Gil et al., 2017). Chandrapala et al. constructed an Inv-deletion mutant strain of Cronobacter sakazakii ATCC29544 and found that the deletion of this gene significantly reduced the efficiency of bacterial entry into host cells, and also proved that inv gene could cooperate with ompA in vitro and in vivo to promote the pathogenicity of C. Sakazakii (Chandrapala et al., 2014).
It has been reported that the nanART gene cluster required for the use of exogenous sialic acid is unique to C. Sakazakii, and this metabolism may be the reason why C. Sakazakii is dominant in neonatal and infant infections (Joseph et al., 2013). In addition, curli fimbriae, a major component of biofilms that plays a key role in biofilm structure and adhesion to various surfaces, is encoded by clusters of csg genes (DeBenedictis et al., 2016). The gene csg was only predicted in C. malonaticus strains in our article. Hu found that csgA was found in C. dublinensis, C. malonaticus, C. turicensis, and C. universalis but not in C. sakazakii and C. muytjensii (Hu, 2018). The expression of adherence factors such as fimbriae or pili and outer membrane protein adhesins is a common trait possessed by all Cronobacter spp. strains. However, little information is known about the function of csg in the mechanisms of intestinal adherence among C. dublinensis, C. malonaticus, C. turicensis, and C. universalis. C. malonaticus B11 lacked some virulence genes, while B20 carried more virulence genes (Fig. 1B). Both of them performed third-generation sequencing by using the Nanopore platform.
MLST typing of 13 strains was conducted, and it was found that they belonged to different ST types (Table 3). Moreover, three important STs were defined: ST4 in the C. sakazakii strain B6, ST1 in the C. sakazakii strain J11, and ST7 in the C. malonaticus strain J41. These STs were believed to be frequently associated with human infections (Mashoufi et al., 2017). ST1 and ST4 are clinically relevant strains and the main STs in pathogenic C. sakazakii isolates (Samadpour et al., 2024). C. sakazakii ST4 is responsible for a large proportion of the documented severe neonatal infections, especially neonatal meningitis (Joseph and Forsythe, 2011). ST7 was also the main ST in C. malonaticus isolates (Alsonosi et al., 2019). Of note, ST7 is the most common ST isolated from adults with Cronobacter infections (Alsonosi et al., 2019). The WGS result showed inconsistent results with the PCR detection result. For example, inv was not found in C. malonaticus by PCR detection (Table 2). However, B20 carried inv gene by WGS analysis (Fig. 1B). Primer limitation may cause false negative results. WGS combined with initial PCR screening could offer more accurate and overall results.
Cronobacter spp. can induce apoptosis of host intestinal cells (Liu et al., 2012). This article aimed to explore the virulence differences of different species of Cronobacter by inducing apoptosis of Caco-2 cells. The two species, C. sakazakii and C. malonaticus, showed higher apoptosis induction abilities than that of other Cronobacter species. Studies have shown that neonatal death is only related to C. sakazakii and C. malonaticus, and only a small number of C. turicensis have been found in clinical infections (Huang et al., 1999). In this article, the test Cronobacter spp. strains demonstrated the diversity of the apoptosis induction rate and contained different putative virulence genes. Huang et al. confirmed that ibeB deletion mutants could not invade HBMEC (Huang et al., 1999). Wang et al. constructed an ibeB gene expression mutant strain and confirmed that compared with the wild-type strain, the inactivation of ibeB resulted in reduced invasion ability to DF-1 cells, and the ibeB deletion strain was found to have reduced toxicity in animal model experiments (Wang et al., 2012). Notably, C. sakazakii J11 contained ibeB gene, while it demonstrated lower apoptosis induction rate than most of C. sakazakii strains (Table 4).
The apoptosis rate of Caco-2 cell induced by different Cronobacter spp. strains
Conclusion
The continuous dynamic monitoring of Cronobacter spp. on infant rice cereal is necessary, and further research into the underlying virulence mechanisms is required, due to its ability to persist in extremely dried foods. The limitation of this study still exists. The sequencing samples were insufficient, thus not providing more information for comparative genomic analysis.
To our knowledge, this is the first time to investigate the difference of pathogenic potential between C. sakazakii and C. malonaticus strains isolated from infant rice cereal in China. Based on WGS, the findings of our article will allow for a better understanding of Cronobacter spp. virulence. As the information of virulence factors determining the pathogenicity of Cronobacter species is still limited, it is important to understand which gene products are responsible for the pathogenicity of different Cronobacter species and how the expression of these virulence factors is regulated.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by State Key Laboratory of Microbial Technology Open Projects Fund of Shandong University (Project NO. M2023-08), Wuhan Science and Technology Bureau (No. 2023020201010208), and Jiangxi Quality Supervision and Administration Bureau (Grant No. GSJK202317).
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.