
Abstract
Despite advances in medical management and device therapy, chronic heart failure (CHF) remains a condition of high mortality and poor quality of life. Patients with CHF endure frequent admissions to hospital, with exacerbations of breathlessness or recurrent acute myocardial infarction and have a high incidence of sudden death. A high intake of marine polyunsaturated fatty acids (PUFAs) is associated with lower cardiovascular mortality in the general population, and diabetics, and can reduce cardiovascular deaths post-infarction. Many of the effects of PUFAs could be of benefit in CHF patients. They can improve endothelial function, reduce vascular tone, reduce platelet aggregability, improve myocardial relaxation, stabilize myocardial cells by prolonging the refractory period, and lead to increased appetite and weight gain. They also have potentially important immune-modulating effects, reducing cytokine production and release and altering prostaglandin metabolism. In this review article we discuss the potential benefits of PUFA supplementation in CHF patients using data from clinical trials and in vitro experiments.
Introduction
Heart failure is common [1]. It is a leading cause of death in industrialized countries [2], and the single most common reason for medical admissions to hospital [3]. Recent therapeutic advances have led to benefits on symptoms and prognosis [4].
Recurrent acute myocardial infarction, worsening chronic heart failure (CHF) and sudden (presumed arrhythmic) death are common causes of deterioration leading to hospital admission or death [5]. Conventional therapies are only partially successful leaving patients with a poor prognosis, persistently high readmission rates [6] and reduced quality of life [7].
The association between a diet high in marine polyunsaturated fatty acids (PUFAs) and low cardiovascular mortality in an Eskimo population was first observed by Sinclair in the 1940s [8]. Recent randomized post-infarction studies have suggested that there is a therapeutic benefit of these agents [9, 10] with a significant reduction in cardiovascular death. Subanalysis of the GISSI-prevenzione study, which compared a combination of two fish oils with placebo in more than 11 000 post-infarction patients on otherwise conventional treatment, revealed a significant reduction in sudden death by 3 months (45% over 3 years) and a significant reduction in total mortality by 3 months [11].
Marine PUFAs (fish oils) have also shown promise in patients with cancer cachexia in whom they can reduce markers of inflammation and lead to weight gain [12, 13].
The aim of this article is to discuss the possible benefits of PUFA supplementation in CHF patients using data from clinical trials and in vitro experiments.
Methods
We identified all articles listed on Medline containing the words ‘fish oils', ‘polyunsaturated fatty acids', ‘eicosapentanoic acid' and ‘docosahexanoic acid' and their abbreviations (PUFA, EPA and DHA). We considered the human and animal data for cardiovascular, cellular and metabolic activity. The data of most potential relevance to chronic heart failure was summarized and divided into sections each concentrating on a particular aspect of the syndrome of CHF.
Fatty acid metabolism
A fatty acid is comprised of a long hydrocarbon chain with a single terminal carboxyl group. Fat is mostly comprised of triglycerides, which are combinations of three fatty acids bound together with a glycerol molecule. The particular fatty acids determine whether the fat is solid or liquid at room temperature. Fats are an important source of calories, and their fatty acid constituents are intermediates in the synthesis of phospholipids and eicosanoids (prostaglandins and leukotrienes). Fatty acids may be saturated (no double carbon bonds, such as stearic acid), mono-unsaturated (single carbon double bond, oleic acid) or polyunsaturated (two or more double bonds, linoleic acid). There are several methods used to name these molecules. The commonest fatty acids are recognized by their ‘trivial names'. Fatty acids can also be named by the ‘omega system', which indicates the number of carbon atoms, the number of double bonds, and the position of the double bond closest to the terminal carbon, the carbon atom furthest from the carboxyl group (omega carbon). The major unsaturated fatty acids fall into omega-6 and omega-3, or n-6 and n-3 groups. For example, linoleic acid is a polyunsaturated fatty acid with two double bonds, 18 carbon atoms, and the first double bond is six carbons away from the omega carbon (n-6). Linolenic acid on the other hand is also 18 carbon atoms long, but has three double bonds, and the first double bond is three carbons away from the ‘noncarboxyl end' (n-3). For simplicity, we shall group those of importance by the position of the double bond (n-3 or n-6).
Fatty acids can be broken down or desaturated to other fatty acids by fatty acyl CoA. However, the specificity of distance from the carboxyl group and the requirement for desaturation of at least a 16-carbon chain means that n-3 and n-6 fatty acids cannot be synthesized in humans. This characteristic and their importance as the base agents in metabolism makes them ‘essential' fatty acids and they must therefore be supplied in the diet.
In the typical Western diet, 20–25-fold more n-6 fats than n-3 fats are consumed [14]. Linoleic acid (LA), which is present in high concentrations in soy, corn, safflower and sunflower oils, is the major n-6 fatty acid. There is a low intake of the n-3 homologue of LA, α-linolenic acid (ALA), which is present in leafy green vegetables and in flaxseed and canola oils. Once ingested, these 18-carbon fatty acids are desaturated and elongated to 20-carbon fatty acids. LA is converted to arachidonic acid (AA) and ALA is converted albeit inefficiently to eicosapentanoic acid (EPA). There is little dietary intake of AA and EPA, which are present in meat and fish. The dietary ratios of n-6 and n-3 fatty acids determine the cellular ratios of LA and ALA and also the relative quantities of AA and EPA produced. Inuit Eskimos have a higher relative intake of the two principal n-3 fatty acids EPA and docosahexanoic acid (DHA) due to a higher intake of fish and fish-eating mammals than western populations [15].
The enzymatic conversion of the essential fatty acids occurs by the same pathway (Fig. 1). Due to competition for the cycloxygenase enzyme (COX), the addition of EPA to the cellular environment significantly alters the relative concentrations of the interconversion products, reducing the relative concentrations of the products of AA metabolism [16–20].
Direct cardiovascular and haemodynamic effects of fish oils
Blood pressure
Marine oils in relatively high doses can lower blood pressure in patients with hypertension and hypercholesterolaemia [21–23], probably due to changes in the physiochemical properties of cell membranes and reduced vascular tone [24, 25]. Blood pressure lowering seems to be unrelated to sympathetic activity or vascular reactivity to adrenergic neurotransmitters [26, 27], and is augmented by salt restriction [28], suggesting an effect on the renin–angiotensin system [29]. Recent work in rats has demonstrated altered T-cell calcium signalling as a possible mechanism [30]. Fish oils also increase nitric oxide release in porcine coronary arteries [31].
Lipid profile and ischaemic heart disease
Fish oils have a beneficial effect on lipid profile lowering triglyceride levels [32, 33], and potentiating the effects of statins [34], and although there were some concerns about impaired glycaemic control in diabetics [35–37] this has not been borne out in a large randomized trial [38] or meta-analyses [32, 33]. High fatty fish intake is associated with a reduced mortality from cardiovascular disease [38–44]. The GISSI-prevenzione study examined the benefits of adding DHA and EPA to conventional therapy in 11 323 patients less than 3 months after a myocardial infarction, and demonstrated an important reduction in total mortality [9]. After 4 months of follow-up the active therapy group had a significant reduction in sudden cardiac death, and after 6 months this group had a lower overall cardiac mortality. These are similar results to data from the DART trial where 2033 men patients consumed fatty fish twice a week following an index myocardial infarction [10]. Data looking at the long-term (10-year) benefit of this dietary modification were less impressive, although many patients had not maintained the altered diet [45]. Despite data suggesting reduced mortality, and improvements in vascular tone, and evidence from work in rats suggesting reduced markers of myocardial ischaemia [46], it is unclear whether fish oils improve subjective or objective markers of myocardial ischaemia in patients with stable symptomatic coronary artery disease [47].
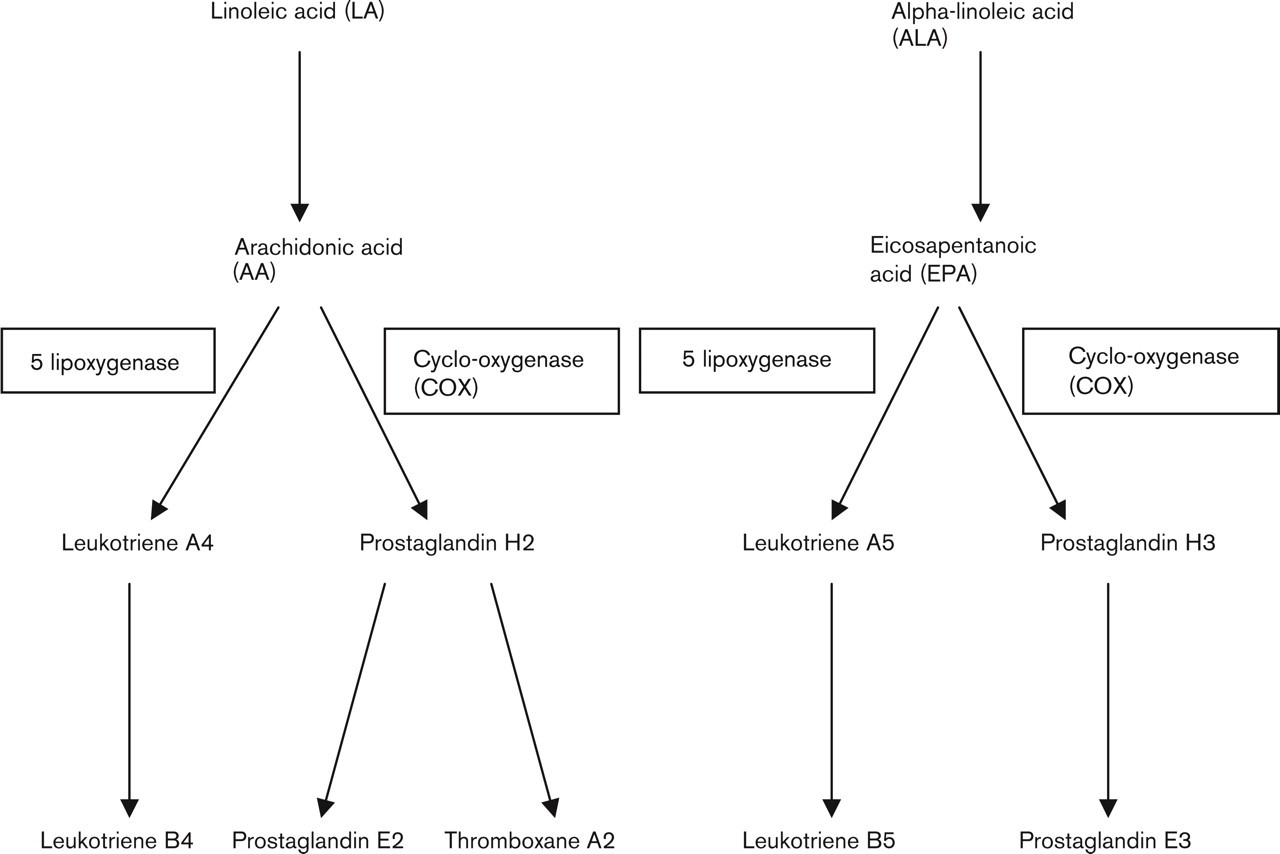
Fatty acid metabolism with the alternative products of n-3 fatty acids. See text for abbreviations.
Cardiac function and ischaemia
Marine oils reduce heart rate [48, 49], and improve diastolic left ventricular function in marmosets and humans [49–51]. This may be due to increased elastic properties of myocardial cells [50] and increased nitric oxide production [52] and release [31].
EPA and DHA but not n-6 fatty acids protect myocardial cells against hypoxia-reoxygenation-induced injury [53], perhaps by inhibiting neutrophil infiltration into infarcted or ischaemic myocardium [54]. Contractile performance of infarcted myocardium can be preserved [55].
Renal function
Fish oil supplementation can increase renal blood flow, reduce renal vascular resistance and increase glomerular filtration rate (GFR) [56] possibly due to altered (prostaglandin E) PGE metabolism. Fish oils can preserve renal function and lower blood pressure in heart transplant recipients [57].
Anti-inflammatory and immunomodulatory effects
Cytokines
Tumour necrosis factor-alpha (TNFα) is raised in heart failure [58, 59], particularly in those with weight loss [60, 61] and higher levels predict a worse prognosis [62]. It directly stimulates loss of appetite via altered beta-3 and leptin receptor activity [63, 64]. IL-6 is also elevated [59].
Cytokines also stimulate collagenase production [65, 66], increase the expression of adhesion molecules and encourage migration and proliferation of smooth muscle cells in the vascular wall [67].
There is an inverse exponential relation between mono-nuclear cell EPA content and cytokine production. Whether the cellular source of EPA is the diet or endogenous synthesis from dietary ALA, cytokine synthesis decreases as cellular EPA concentrations increase [68, 69]. Increased dietary n-3 fats suppress the production of both TNF-α and IL-1β [68, 70–72]. This is due to an inhibitory effect of EPA on lipopolysaccharidestimulated TNF gene transcription and protein elaboration in monocytes [73]. ALA supplementation can reduce IL-1β and TNF-α production by more than 30% after 4 weeks [68]. EPA and DHA also inhibit IL-6 production in human endothelial cells [74] and reduce the concentrations of TNF-α and IL-1β in the arterial wall [75], thereby potentially reducing smooth muscle cell migration and proliferation.
Prostaglandins
EPA competitively inhibits AA metabolism by competing for COX enzyme sites and can therefore alter the production of inflammatory mediators (Fig. 1) by the preferential formation of prostaglandins of lower pro-inflammatory potency (for example PGE3) and overall to significantly reduced inflammatory prostaglandin production [76, 77]. EPA also competes with AA for 5-lipoxygenase leading to the formation of leukotriene B5 (LTB5), which has little inflammatory activity compared with LTB4 [78, 79]. Thus, increasing dietary n-3 fats shifts the balance of the eicosanoids produced to a less inflammatory mixture. However, the formation of PGE itself may be anti-inflammatory suggesting that direct competition may not be the only anti-inflammatory mechanism encouraged by fish oils [80].
There are concerns that moderate doses of fish oils > 2 g per day could affect the immune response [81], predominately T-cell responses [82], but even lower doses (150mg DHA+30mg EPA) can modify the cellular immune response in elderly humans [83]. These changes might put elderly patients with concomitant chronic disease at increased risk of acute infections. This matter is not resolved, however, as there are conflicting data about the effects of ALA (a source of DHA and EPA) on cytokine production. One study on healthy elderly human subjects suggests that ALA leads to reduced IL-6 production by stimulated mononuclear cells with no reduction in their production of TNF-α, interleukin (IL) 1beta, IL-2, IL-4, IL-10 or interferon-gamma [84]. There is also no effect on the numbers of or the functional activities of neutrophils, monocytes or lymphocytes with moderate fish oil supplementation in healthy adults [85, 86]. Other studies have shown a reduction in cytokines as mentioned above [67, 70–72]. In view of these conflicting results further research is required to examine the effects on elderly patients.
Haemostasis
Patients with CHF are at increased risk of thromboembolism. Ingestion of n-3 fatty acids leads to suppression of TXA2 synthesis by platelets and by blood mononuclear cells [87, 88]. Decreased platelet aggregation has been demonstrated with EPA supplementation in some [89– 91] but not all studies [92–94].
EPA can reduce the expression of vascular cell adhesion molecule-1 (VCAM-1), endothelial leukocyte adhesion molecule-1 (E-selectin or ELAM-1) and intercellular adhesion molecule-1 (ICAM-1) [95]. These adhesion molecules are elevated in CHF and plasma levels are related to the severity of the condition [96, 97]. EPA and DHA suppress adherence of monocytes to activated endothelial cells also by affecting endothelial platelet aggregating factor (PAF) generation [98].
Endothelin 1
Endothelin 1 (ET-1) is a powerful vasoconstrictor [99] and higher levels of ET-1 predict a poor prognosis in CHF patients. EPA suppresses endothelin-1 (ET-1) production in human coronary artery smooth muscle cells [100].
Cachexia
Patients with severe CHF often lose weight. This is a consequence of the inflammatory processes, reduced intake and increased catabolism. Weight loss or frank cachexia is commonly seen in heart failure [101], the prevalence increasing with worsening symptoms [102, 103]. Cachexia worsens the already poor prognosis by a factor of 2.6 [102]. More subtle loss of muscle bulk occurs early in the disease [104].
There may be multiple aetiologies to the weight loss [105]. There is a general predominance of catabolic processes over anabolic in heart failure [106]. Catabolic steroids are elevated in untreated patients [107] and in treated patients there is an increase in catabolic relative to anabolic steroids [60, 108]. EPA is effective in attenuating the increased protein catabolism in cancer cachexia [109–111]. EPA also inhibits lipolysis [112].
Arrhythmias
Patients with chronic heart failure have a high incidence of sudden death [113]. One of the major effects seen in the GISSI-prevenzione post-infarction study was a reduction in sudden death by 45% [9]. There is an inverse association between blood levels of fish oils and sudden death [114]. Fish oils have been shown to have an anti-arrhythmic effect in vitro and in vivo [115–117], even when administered acutely [118–121]. They can prevent ischaemia-induced ventricular fibrillation [121]. The anti-arrhythmic effect is related to their ability to reduce the electrical excitability and automaticity of cardiac myocytes [118, 131]. They increase the electrical stimulus required to elicit an action potential by approximately 50% and prolong the relative refractory time by approximately 150% [122].
Fish oils can prevent or terminate the tachyarrhythmias induced by isoproterenol, or its intracellular messenger, cAMP [119]. Despite this, cardiomyocytes exposed to DHA have a greater chronotropic response to beta-agonists than those without this fish oil [123], possibly due to increased cAMP efficiency.
Secondary prevention trials of myocardial infarction survivors have suggested that high intakes of fish [10] and ALA [124] reduce the incidence of fatal cardiac arrhythmias. In survivors of acute myocardial infarction with left ventricular dysfunction, there was a correlation between platelet DHA, the patients' intake of fish and heart rate variability [125]. Human studies have demonstrated a reduction in ventricular extrasystoles [126], and increased heart rate variability with fish oil supplementation [127, 128]. Increased dietary intake of marine oils has been linked with a lower incidence of primary cardiac arrest [129]. By reducing sodium influx [130], and shortening the action potential [131], fish oils can shift the threshold for excitation to more positive potentials, and prolong the relative refractory period.
N-3 polyunsaturated marine oils prevent ischaemia-induced malignant cardiac arrhythmias in animal studies [115, 117, 132–138] The presence of fish oils accelerates the electrical depression during hypoxia but allow a faster recovery upon reoxygenation. By increasing the ratio of EPA to AA within cellular membranes there is an increase in the Ca2+-Mg2+-ATPase activity within myocardial membranes [139]. These cellular alterations reduce the severity of ventricular arrhythmias following the ischaemia by altering cardiac sarcoplasmic reticulum Ca-ATPase function, and thereby inhibiting the rapid accumulation of intracellular Ca2+ [140].
Despite the observed effects of fish oils on arrhythmic mortality in post-infarct patients, there are conflicting data in patients with automatic implantable cardioverter defibrillators (AICD) [141]. In a population of 200 patients over 2 years there was a non-significant trend to an increased risk of ventricular arrhythmia in patients randomized to n-3 fatty acids. Subgroup analysis suggested that patients who had received an AICD for sustained VT had more arrhythmias on fish oil, whilst patients enrolled with VF tended to have fewer. There were only two sudden deaths, both on fish oil but there was a strong trend for reduced overall mortality (P = 0.09) with fish oil. This trial suggests that if n-3 fatty acids do reduce mortality, they may do so by a mechanism other than reduction in ventricular arrhythmias. As the substrate for the induction of VT and VF may be different, a specific effect on VF suppression cannot be excluded. This is a small study though, with a small number of events and it is possible that the effects seen are due to chance.
Heart failure data
There are no published data examining the effects of fish oil supplementation in patients with CHF. One study examining cachexic heart failure in dogs demonstrated that fish oil supplementation decreased Il-1 concentrations and improved cachexia [142].
Patients with chronic heart failure die of ischaemic events [143], arrhythmias and deterioration of their heart failure. They are at greater risk of death if they are cachexic and have high circulating markers of inflammation. Fish oils lower blood pressure, reduce ischaemia, reduce platelet aggregation and prevent arrhythmias. They reduce inflammatory mediator production and lead to weight gain. Fish oil supplements have previously been limited by poor tolerability, but newer preparations are well tolerated allowing for high-dose supplementation and long-term use [144].
Summary
Fish oils might be of benefit in patients with both ischaemic and non-ischaemic CHF through several mechanisms. They have anti-ischaemic, anti-arrhythmic and antithrombotic activity. In patients with cachexia, whose outlook is particularly poor, they might have important anticachectic and anticytokine effects. The clinical implications of the changes in cellular immune responses that fish oils might cause remain unclear, but patients with CHF are already at increased risk of acute infections. Any trial involving elderly patients would have to proceed cautiously with interim analyses looking for an increased incidence of infection in those receiving fish oil supplementation. The present review has focused on systolic heart failure. It is possible that patients with diastolic heart failure might benefit from these agents through similar mechanisms.
The time has come for a randomized, controlled trial in CHF patients who have potentially much to gain from these agents and the results of the GISSI-CHF trial [141] are eagerly awaited.
Footnotes
Acknowledgements
The authors are grateful to Ms Julie Hannah of the Dietetic Department, Hull Royal Infirmary for advice and suggestions during the preparation of this manuscript.