
Abstract
Background
In some cases of hemiplegia, the initiation of yawning is associated with involuntary raising of the paralysed arm. Reports are scarce in the literature, probably because the phenomenon has largely been overlooked.
Methods
We studied six patients from two neurologic units, and compared them with published cases from the last 200 years. Brain imaging typically shows a small vascular lesion most often located in the internal capsule.
Results
After comparison with experimental models in cats, we suggest that damage to the cortico-neocerebellar tract of the extrapyramidal system disinhibits the spino-archeocere-bellar tract, enabling a motor stimulation of the arm by the lateral reticular nucleus, which harmonises both central respiratory and locomotor rhythms.
Conclusions
When phylogenetically primitive structures are disinhibited, they regain autonomy in the homeostatic process associating the massive inspiration of yawning – a form of behaviour that stimulates vigilance – with a motor control mechanism that is active during locomotion. For this phenomenon, we coined the term ‘parakinesia brachialis oscitans’.
Introduction
In 1844, the following case was reported in ‘The nervous system of the human body as explained in a series of papers read before the Royal Society of London’ by the Scottish physician John Abercrombie (1780–1844), an author well known for his exact observations and insights on the associated pathology: ‘I had some time ago under my care, a man affected with hemiplegia of the left side, the palsy complete, without the least attempt at motion, except under the following circumstances: he was very much affected with yawning, and every time he yawned the paralytic arm raised up, with a firm steady motion, until it was at right angles with his body (as he lay in bed on his back), the forearm a little bent inwards, so that his hand was above his forehead at its greatest elevation. The arm was raised steadily during inspiration, and when the expiration began it seemed to drop down by its own weight with considerable force. He continued to be liable to the affection for a considerable time, and it ceased gradually as he began to recover the natural motion of the limb (1,2)’.
Using recent case reports collected from two different sites in France and Switzerland, we aim to present this curious phenomenon, which involves the combination of involuntary motion in a paralysed upper limb with yawning. We studied six patients with this clinical phenomenon, for which we coined the term ‘parakinesia brachialis oscitans’, and reviewed historical reports and experimental data, in order to delineate a pathophysiological explanation.
Case reports
Case 1 (Montreux)
A 49-year-old woman with no history of vascular disease and few recognised risk factors, including a long history of migraine with aura and cigarette smoking, developed a fluctuating hemiplegia on the left side, shortly after waking up. Weakness predominated in the upper limb, with moderate dysarthria. Sensory testing was unremarkable, and no associated neurologic dysfunction was found on examination. Diffusion-weighted magnetic resonance imaging (MRI) showed a small infarct in the posterior limb of the right internal capsule, associated with marked leukoaraiosis. Blood pressure was normal, as were cardiac investigations and extra/intracranial MR angiography. A positive Notch3 diagnostic mutation of cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy (CADA-SIL) was found (her father suffered from dementia and multiple strokes).
After 2 days, the patient reported that while she was unable to move the left upper limb voluntarily, it rose every time she yawned, up to the level of her chest, when she was in a sitting position. This lasted for a few seconds and was not associated with specific movements of the hand and fingers. We ourselves witnessed this movement, which disappeared as motor recovery developed over the following 2 weeks.
Case 2 (Montreux)
A 73-year-old man, treated for high blood pressure and cholesterol, developed sensorimotor hemiparesis on the right side, without cognitive changes or aphasia. Initially, he was completely paralysed in the upper limb, but during episodes of yawning, the same limb would move about 30 cm upwards, remain still for a couple of seconds and then slowly return to its primary position. This was mainly noted when the patient was sitting in a wheelchair, and disappeared after 1 week, while the patient recovered some voluntary motor function in the upper limb, with ataxia. Magnetic resonance imaging showed a small infarct in the anterior limb of the left internal capsule, suggesting lacunar infarction. An occlusion of the V4 segment of the left vertebral artery was present, but no large artery disease was found in the anterior circulation, and cardiac tests showed no embolic source.
Case 3 (Poitiers)
A right-handed, 71-year-old man was treated for diabetes and hypercholesterolaemia. In November 1996, he presented with an isolated motor deficit in the left leg, resolving in 24 h. The assessment of this transient ischaemic attack revealed 40–50% bilateral internal carotid artery stenosis. A few days later, MRI of the brain showed no residual lesions. Transoesophageal echocardiogram was normal. The patient was prescribed aspirin 250 mg/day In September 1999, he experienced a sudden vertigo, rapidly followed by severe left hemiplegia without sensory impairment. Brain CT scan was normal upon admission. Doppler examination was unchanged. A transoesophageal echocardiogram revealed a left intra-atrial thrombus. A second CT scan showed a hypodensity in the right centrum semiovale and lenticular nucleus with leukoaraiosis. The diagnosis was embolic occlusion of the lenticulostriate branches of the right middle cerebral artery (MCA). The patient was put on heparin, with a vitamin K antagonist as relay treatment.
One year later, the patient still had severe left spastic hemiplegia. He reported the involuntary lifting of his totally paralysed left arm during yawning. In 2–3 s, the arm rose around 10 cm, with adduction and flexion of the elbow, until the patient opened his mouth. The arm fell inert once the patient closed his mouth. This could occur while he was sitting, standing or lying down. He had observed this phenomenon from the onset of his hemiplegia. This involuntary movement of the paralysed arm during yawning was still present 3 years after his stroke.
Case 4 (Poitiers)
A right-handed, 53-year-old man with a history of heavy smoking, hypercholesterolaemia and Factor V Leiden mutation, with two episodes of deep vein thrombosis, while on an oral anticoagulant, had, in May and June 2001, two transient ischaemic attacks with left brachiofacial deficits. During hospitalisation in June 2001, he suddenly suffered a severe left hemiplegia due to a middle cerebral artery infarction (MCA) with MCA occlusion and dolichoectatic carotid siphon and an infarct involving the right caudate nucleus and centrum semiovale. In the following hours, the patient was drowsy and yawned deeply in an abnormally frequent and repeated way. At the beginning of each yawn, the left arm rose involuntarily with flexion of the elbow and adduction. This gesture lasted for the duration of the yawn. The muscular force was so great that the patient's wife, seated next to him, was unable to counter it. He never recovered and, 1 year after his stroke, his arm rose at every yawn.
Case 5 (Poitiers)
A right-handed, 75-year-old, alcoholic man presented on 7 November 2001 with right hemiparesis resolving in an hour. Brain CT scan was normal. He received one injection of Enoxaparin 40 mg per day. On 10 November 2001, a brachiocephalic right hemiplegia developed, associated with Broca's aphasia. Doppler examination revealed a bilateral 50% carotid stenosis. Vertebral arteries were normal. Echocardiography showed a dilated left atrium, concentric left ventricular hypertrophy and moderate mitral insufficiency. Echocardiography showed normal sinus rhythm. Brain CT scan revealed an infarct in the left internal capsule and lenticular nucleus. From the onset of the patient's stroke, an increased frequency of yawning was noted. With each yawn, the right arm rose with adduction and elbow flexion, falling as soon as the patient closed his mouth.
Case 6 (Poitiers)
A right-handed, 35-year-old woman suffered, 2 months after delivery, an unusual, diffuse, intense headache resistant to analgesics. Two days later, she was found lying on the ground, paralysed and aphasic. Upon admission, there was a massive complete right hemiplegia with aphasia and drowsiness. Blood pressure was 12/7 and cardiac rhythm was sinusal. CT scan on admission was normal. Doppler examination was normal on the right. On the left, a complete occlusion of the internal carotid artery, with no distal flow, was shown. Brain CT scan in the following days showed a total MCA territory infarct. Angiography showed signs of fibromuscular dysplasia and left carotid dissection. From the initial phase of complete flaccid hemiplegia, every time the patient yawned, her completely paralysed right arm rose with elbow flexion and moved adductively towards her face, before falling heavily when she closed her mouth.
Discussion
Although probably unaware of Abercrombie's earlier publication, Dr Liegey of Rambervilliers in France reported, in 1851, the following case published in the Gazette médicate de Strasbourg. ‘A man recovering from repetitive attacks of hemiplegia was overcome by very disagreeable yawning which occurred every day at the same time. No less remarkable, whenever this yawning occurred, it was accompanied by a convulsive movement which raised the patient's previously paralyzed arm. The movement produced a painful sensation in the limb and could only be stopped by the patient strongly grasping the arm with the opposite hand (3).’
Hence, since the early 19th century, a few cases of movements of a paralysed arm during yawning have been reported (1,–19). The aetiology is variable: ischaemic or haemorrhagic stroke, bulbar form of amyotrophic lateral sclerosis, brainstem tuberculoma. Based on these cases and the literature review, this hemiplegia-associated movement shows no lateral preference and may appear from the accident's onset during the flaccid phase, or later, during the spastic phase. The reports mention either abduction or adduction of the arm. While the paralysis leaves the arm immobilised in semi-extension alongside the patient's body, we have observed the arm to move outwards away from the thorax (abduction), followed by elbow flexion bringing the hand towards the sternum (adduction). This movement is strictly concomitant with the yawn; the arm falls inert once the yawn ends. No simultaneous movement is observed in the leg. The arm's movement is totally involuntary and uncontrollable. It coincides with the paralysis and tends to disappear with recovery.
The case reports from 1988 onwards indicate two main locations closely linked to this clinical picture. The first is a lesion in the MCA territory, particularly the lenticulostriate branches, leading to infarction in the internal capsule and lenticular nucleus (paleostriatum) or caudate nucleus (neostriatum) (20). The second is a pontomedullary lesion, as caused by tuberculoma in a 14-year-old girl with Millard-Gubler syndrome (9), and also reported in a series of bulbar amyotrophic lateral sclerosis cases (18) and in a case of basilar artery thrombosis resulting in right-sided pontine infarction (19).
Hemiplegia and associated movements
At the end of the 19th century, movements associated with hemiplegia were already debated. Vulpian (21) used the term synkinesis for ‘movements which take place involuntarily in one part of the body as voluntary or reflex movements are carried out elsewhere’. In pyramidal syndromes in general and hemiplegias in particular, synkinesis produces involuntary movements in paralysed muscles concomitant with voluntary movements in healthy muscles. Classically, there are several types: global synkinesis, which is merely an exaggeration of the contraction on the paralysed side during an effort on the healthy side; coordination synkinesis, when the voluntary contraction of the healthy muscle results in the involuntary contraction of the paralysed synergistic muscles; and imitation synkinesis, when a voluntary movement on the healthy side triggers an involuntary attempt at the same movement on the paralysed side.
de Buck (22) also described movements associated with hemiplegia: ‘Surprisingly enough, they extend even to the extremities of organs not under voluntary control. These processes accompany not only voluntary movements, but also reflex movements – yawning, sneezing, etc.’. A. Souques, writing about hemiplegia in Pratique Médico-Chirurgicale (23), re-examined this distinction: in addition to synkinesis, which only occurs in the spastic phase, ‘associated reflex movements coincide with a cough, a yawn, primarily affecting the upper limb’ and may be observed during the initial flaccid phase. Working in Turin, Bertolotti (9) made the same distinction. And Brissaud wrote in his lecture dated 1 December 1893: ‘Spasmodic yawning is also seen with a certain frequency in hemiplegics. Not merely a superficial symptom amongst the spasmodic elements of epilepsy (Fere) or hysteria (Charcot, Gilles de la Tourette, Guinon and Huet), it indeed seems to arise directly from organic lesions …’ (24). We agree with the disciples of J.-M. Charcot on this point, and do not support using the term synkinesis to describe this involuntary raising of the arm during yawning.
In vertebrates, ethologists describe pandiculation or Rekel-Syndrome as the association of yawning with a stretching of the trunk and four limbs (25). Bertolotti (9) and Blin et al. (17), each reporting cases similar to those described above, use the term pandiculation or hemipandiculation. Neither the description of the arm's involuntary, non-stretching movement, nor the absence of stretching in the leg on the same side supports the use of this term.
de Buck (22) wrote: ‘Parakinesia appears to differ from synkinesia by the fact that the relationship between the idea and the act in the latter is perfectly preserved, involving only a simple diffusion of the nervous influx at the motor centres, owing to the need for a much stronger signal to produce a useful effect; in parakinesia, the relationship between the idea and the movement appears disrupted’. We have therefore proposed the term parakinesia brachialis oscitans. Parakinesia is an abnormal movement that acts as a parasite, caricature or replacement of a normal movement, in this instance of the arm (brachial). Oscitans comes from the Latin oscitantis, meaning ‘which yawns’ and mentioned in early writings on fever (26).
Yawning
Yawning is phylogenetically ancient, resembling what ethologists refer to as maintenance behaviour. Highly stereotypical, it is observed in cold-blooded and warm-blooded vertebrates, from reptiles with rudimentary, ‘archaic’ brains to human primates, in water, air and land environments. The ethology, neurophysiology and neuropsychology literature associates yawning with wake/sleep rhythm fluctuations, eating and sexuality, where it externalises a group of vigilance-stimulating mechanisms and attests to the hypothalamus' central role in homeostasis (27,–29). Yawning is recognisable in ultrasound images from the 14th week of pregnancy, and like the appearance of oromandibular movements and swallowing, it signals functional maturation of the brainstem and basal ganglia, whereas the extension to the frontoparietotemporal cortex continues through the 24th week (30). No brain structure has ever been identified as the yawning centre. Clinical and pharmacological evidence suggests that the hypothalamus – particularly the paraventricular nucleus – plays a key role in yawning, as well as medullary and pontine regions, with connections towards the frontal region in human primates and towards the cervical spine. The muscles that contract during yawning are supplied by cranial nerves V, VII, IX, X, XI and XII, cervical nerves C1–C4 (phrenic nerve) and the dorsal nerves innervating the intercostals, which play an accessory role in breathing (31).
Provine et al. (32) showed that yawning has no effect on arterial oxygenation. Hence, the old paradigm, which from the time of Hippocrates to the mid-20th century interpreted this behaviour as a reflex to increase brain oxygen supply, must now be replaced with a neuromuscular paradigm involving subcortical control. The hypothalamic paraventricular nucleus contains the cell bodies of oxytocin neurons projecting to the hippocampus and to the reticular formation and locus ceruleus in the brainstem. When these neurons are stimulated by dopamine, excitatory amino acids or oxytocin itself, they trigger yawning by releasing oxytocin in these various subcortical structures. This oxytocinergic activation is inhibited by opioids, which prevent yawning. The activation or inhibition of these oxytocin neurons is correlated with the activity of paraventricular nitric oxide synthase. Other neuronal systems are involved as modulators. Serotonin, oestrogens, testosterone and hypocretin play a role either in the paraventricular nucleus or in the motor nuclei of the brainstem and spinal cord, with the final executive pathway controlled by acetylcholine (33,–35).
Attempt at a pathophysiological explanation
The release of subcortical structures from cortical inhibition is classically proposed to explain certain automatic or reflex activities occurring experimentally in decorticate animals, or following stroke in humans. This is the case for palatal myoclonus, or palatal tremor (36), caused by destruction of the central tegmental tract, allowing uncontrolled activity in the olivary body and, in turn, the palate's rhythmic movement. This amounts to a re-emergence of the structure's phylogenetically branchial function.
Another example is the syndrome of Foix et al. (37), or biopercular syndrome, where the automatic activities of emotional expression, swallowing and yawning remain intact even though there is total faciopharyngoglossomasticatory paralysis. The automatic activity depends on the brainstem and is preserved, whereas the voluntary control of the prerolandic cortex and the supplementary motor area is interrupted by a biopercular infarct. One MRI case study showed this automatic-voluntary dissociation as a corticobulbar disconnection (38).
A final example is the locked-in syndrome, in which there is total paralysis and patients may only retain voluntary control over eye movement. However, involuntary facial expressions of pain are preserved, along with yawning and loud crying. The corticospinal or the pyramidal tract is completely interrupted at the pons. The preserved activity of extrapyramidal and cerebellar structures and of archaic spinal automatisms explains the automatic-voluntary dissociation (39).
Could such a mechanism of cortical-subcortical dissociation explain the synchronisation between the raising of the arm and the respiratory cycle of the yawn, due to diaphragmatic extension?
The study of the relationship between respiratory cycles and movement was initiated by Marey in 1892 (40) through his invention of chronophotography, a precursor to the cinema. He had already shown that the breathing cycle of a running dog, a galloping horse and a flying gull was based on stride or wingbeat. This synchronicity allows optimal adaptation between oxygenation and muscular work, but the resulting piston effect of the diaphragm's movement also plays an important role in bodily and aerodynamic equilibrium. Energy transfers between trunk muscles and those used for breathing improve the energetic yield of running (41,–44). Humans maintain partial coupling through arm-swinging during walking, a sign of extrapyramidal activity (45, 46).
Respiratory automatisms are the result of several medullary and pontine nuclei working in concert. Experimental data suggest that respiratory rhythm generation depends on pacemaker neurons. These neurons are found in a small region known as the pre-Bötzinger complex, ventral to the nucleus ambiguus and to the emergence of the roots of cranial nerve XII. The pacemaker-generated rhythm is modulated by networks of respiratory interneurons, responsible for its final spatiotemporal organisation. Inspiratory and expiratory neurons have been described according to the respiratory phase concurrent with their firing time. The rhythmic signal from these neuronal interactions is then distributed, according to a precise spatiotemporal pattern, to medullary and spinal motor neurons, resulting in the motor innervation of the upper airway and respiratory muscles (47, 48).
Adjacent to these nuclei, the lateral reticular nucleus projects to the cerebellum and plays a role in the sensory-motor coordination of the limbs. This corresponds to the spinoreticulocerebellar pathway. Experiments in the cat have shown that when the nuclei in the respiratory rhythm complex are firing, the lateral reticular nucleus itself has a rhythmical and synchronous activity, particularly when limb extensor muscles are contracted during stretching (49,–52). The dual role of the intercostals and the diaphragm in posture and locomotion, on the one hand, and in breathing, on the other (not to mention phonation), requires not only somatomotor coordination but also neurovegetative coordination, especially of cardiovascular functions. In the cat, Schomburg et al. (53) showed sympathetic modulation of cardiolocomotor and respiratory activity with a spinopontine feedback loop extending to the hypothalamus.
Cardiorespiratory adaptation to effort involves the autonomic nervous system, particularly the hypothalamus and pituitary gland, with regulation of blood pH, PaCO2 but also satiety and vigilance (54, 55). These homeostatic mechanisms, dependent on anatomically similar subcortical structures and regulated by identical neurotransmitters, are linked to those controlling the rhythms of wake/sleep, satiety and sexuality, i.e. those involved in triggering yawning (29, 56).
The extrapyramidal motor system has a key influence on the spinal, brainstem and cerebellar motor circuits as well as the motor cortex itself. Its neurons connect the cerebral cortex to the cerebellum, forming the corticopontocerebellar tract of the neocerebellum, whose phylogenetic development mirrored that of bipedality (57). The cerebellum receives a copy of all motor impulses from the cerebral cortex through this tract (58). This corticopontine pathway passes through the internal capsule on each side of the pyramidal tract. The lesion responsible for parakinesia brachialis oscitans is most often found at this level, involving damage to the first neuron and interrupting the corticospinal and corticonuclear pathways, but also the extrapyramidal corticostriate, corticorubral, corticonigral and corticoreticular pathways (Fig. 1). All six reported cases share this characteristic.
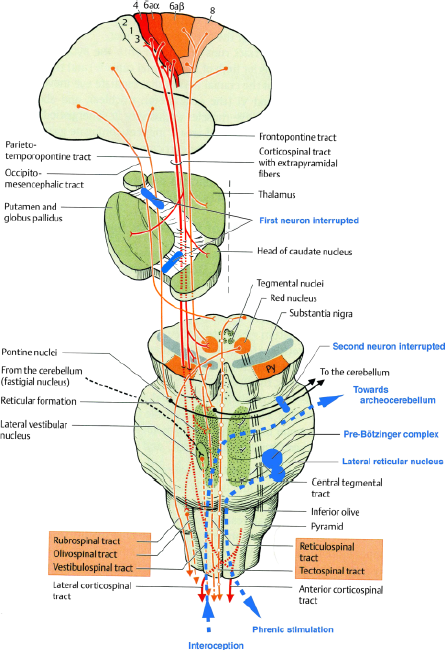
Schematic representation of the extrapyramidal tract and the localisations of lesions responsible for parakinesia brachialis oscitans. (Adapted from Duus' Topical Diagnosis in Neurology, Thieme Ed., 2005).
At the level of the pons, the fibres form synapses in the pontine nuclei whose axons constitute the second neuron projecting to the cerebellum (middle cerebellar peduncle). The cases reported by Bertolotti (9), Louwerse (18) and Topper et al. (19) show that a lesion at this level may also cause parakinesia brachialis oscitans (Fig. 1).
The paleocerebellum receives signals from the spine via the dorsal and ventral spinocerebellar tracts. The two pathways transmit proprioceptive impulses from the peripheral system, i.e. from muscle spindles and Golgi tendon organs in the muscles of the limbs, trunk and diaphragm. Efferents from the paleocerebellum, after being relayed by the emboliform nuclei and the fastigial nucleus, project to the red nucleus via the superior cerebellar peduncle. In this way, the paleocerebellum controls antigravity muscles, determines the appropriate muscle tone for walking, participates in the synergistic coordination of agonist and antagonist muscles necessary for this task, all while coordinating the motor control of the diaphragm. It also sends out ascending signals that project to the centromedian nucleus of the thalamus, then to the caudate nucleus and the putamen, thereby providing a connection with the extrapyramidal system (59).
Like normal walking, pandiculation – the generalised stretching of the trunk, limbs and diaphragm – requires the uninterrupted functional activity of the corticoneocerebellospinal and the spinoarcheocerebellothalamic pathways, coupling the pyramidal and extrapyramidal control of the entire musculature.
Interruption of the corticospinal tract, whatever the aetiology, causes paralysis in the arm, preventing voluntary motion. In certain cases, the corticoneocerebellospinal pathway is also interrupted. However, the proprioceptive loop conducting signals between the motor anterior spinal horn, the paleocerebellum and the lateral reticular nucleus (ventral spinocerebellar tract) remains functional. During yawning, the strong contraction of respiratory muscles represents a proprioceptive signal. An antidromic stimulation from the respiratory nuclei to the anterior spinal horn has been experimentally demonstrated in the cat (50). It might be speculated, albeit boldly, that parakinesia brachialis oscitans is a movement of the arm (akin to the swinging coupled with respiration during walking) secondary to an incoming motor signal in the anterior spinal horn from C4 to C8, originating in the lateral reticular nucleus and travelling through the extrapyramidal pathways of the archeocerebellum (54).
This mechanism is consistent with the absence of movement in the leg (corticospinal interruption) and with the possibility of occurrence in the flaccid as well as the spastic phase. A preserved corticoneocerebellar pathway would prevent such a movement. Therefore, the requisite condition for parakinesia brachialis oscitans would be corticonuclear, corticospinal and corticoneocerebellar interruption, whereas the spinoarcheocerebellar pathway would remain functional. Assuming that yawning is the exteriorisation of a homeostatic mechanism regulating the vigilance systems in the hypothalamus, repetitive yawning, which is frequently observed during stroke, would appear to stimulate arousal mechanisms. When a lesion in the extrapyramidal system prevents the modulating activity of the corticoneocerebellar pathway, structures that are phylogenetically more primitive may allow recovering movement in the arm during the diaphragm's maximal ampliation, as occurs during yawning.
The central nervous system in vertebrates follows a common organisational pattern and shows gradually increasing complexity with higher and higher levels of independence and functionality. The American neuropsychiatrist P. MacLean (60) proposed a model of the nervous system's functional organisation based on the study of its phylogenesis. At the base of this model is the ancestral ‘reptilian’ brain (brainstem and basal ganglia), where yawning originates. The next level is the ‘paleomammalian’ brain (limbic system) shared by all mammals. This is the synaptic and hormonal interface, where emotive yawning in monkeys is localised. Finally, a ‘neomammalian’ brain comprises the top layer, characterised by cortical development in humans, particularly of the frontal lobes, where ‘contagious’ yawning occurs (28). If these functional levels become disconnected, as happens in certain stroke localisations, functions may reappear that are normally inhibited by a phylogenetically more recent and functionally more sophisticated structure. In this way, human pathology reveals that the coordination and regulation of body temperature, breathing, locomotion and vigilance has been perfected over time, with ever-increasing complexity and precision, from reptiles to primates.
Footnotes
Acknowledgements
We would like to thank the following individuals for their help with proofreading and their suggestions: Prof. M. G. Bousser (Neurology Department, Hospital Lariboisiere, Paris), Prof. K. Ezure (Tokyo Metropolitan Institute for Neuroscience, Japan) and Dr P. Wicks (MRC Centre for Neurodegeneration Research, King's College London, GB).