
Abstract
Periodontitis is a chronic inflammatory disease characterized by persistent inflammation, redox imbalance, and mitochondrial dysfunction, leading to progressive impairment of periodontal cellular function and tissue integrity. Current periodontal therapies primarily focus on controlling inflammation and promoting tissue regeneration. However, these approaches do not adequately restore the function of resident cells compromised by an inflammatory microenvironment. Restoration of cellular homeostasis prior to inducing regeneration may enhance treatment outcomes for effective periodontal repair. This study aimed to investigate the role of CPNE7 in periodontal homeostasis and to evaluate whether selcopintide (SCPT), a CPNE7-derived peptide, restores cellular function under inflammatory periodontal conditions and exerts therapeutic effects in a mouse ligature-induced periodontitis model. In aged periodontal tissues, CPNE7 expression was reduced, accompanied by increased oxidative stress and decreased Nrf2 expression. Cpne7 knockout mice also exhibited periodontal structural abnormalities and showed aggravated alveolar bone loss and inflammatory responses following periodontitis induction. SCPT reduced excessive reactive oxygen species accumulation induced by inflammatory stimuli and restored redox balance, thereby supporting cellular homeostasis. In addition, SCPT improved mitochondrial function under inflammatory conditions, as evidenced by restored mitochondrial morphology and bioenergetic function, while attenuating inflammatory signaling and apoptosis. These protective effects were associated with activation of the Nrf2 antioxidant pathway. Importantly, topical administration of SCPT significantly attenuated periodontal destruction and alveolar bone loss in the ligature-induced periodontitis model, demonstrating its therapeutic efficacy in vivo. Collectively, these findings indicate that CPNE7 is an important contributor to periodontal tissue homeostasis and that SCPT restores redox balance, mitochondrial function, and inflammatory stability, highlighting CPNE7-derived peptides as a promising therapeutic strategy for periodontitis.
Keywords
Introduction
Periodontitis is a chronic inflammatory disease that compromises the structural and functional integrity of periodontal tissues, including the periodontal ligament (PDL), cementum, and alveolar bone (Nanci and Bosshardt 2006; Cho et al 2021). Persistent inflammation causes a destructive microenvironment that impairs resident periodontal cells, such as PDL fibroblasts, cementoblasts, and osteoblasts, thereby diminishing their regenerative capacity and contributing to progressive tissue breakdown (Chapple and Matthews 2007; Graves 2008; Meyle and Chapple 2015). A hallmark of periodontitis is the excessive accumulation of reactive oxygen species (ROS), which induces sustained oxidative stress and mitochondrial dysfunction, further exacerbating cellular impairment (Chapple and Matthews 2007; Deng et al 2024; Wadan et al 2025).
Excessive ROS, impaired antioxidant defense, and mitochondrial dysfunction converge to disrupt cellular homeostasis in inflamed periodontal tissues (Dahiya et al 2013; Deng et al 2024). Previous studies have described periodontal homeostasis as a state in which balanced host–microbiota interactions prevent excessive inflammation and tissue destruction (Hajishengallis and Lamont 2012). We extend this concept to the cellular level, defining cellular homeostasis as the capacity of resident periodontal cells to maintain redox balance, preserve mitochondrial integrity, and regulate stress responses required for tissue maintenance and regeneration.
Mitochondrial dysfunction is considered a potent pathogenic mechanism in periodontitis (Deng et al 2024). As the primary intracellular source of ROS, damaged mitochondria amplify oxidative stress, disrupt metabolic activity, and accelerate tissue destruction (Murphy 2009; Brand et al 2013). Periodontitis-associated mitochondria exhibit impaired biogenesis, altered dynamics, and defective mitophagy, contributing to loss of mitochondrial integrity (Jiang et al 2023). Among various antioxidant defense systems that regulate ROS, nuclear factor erythroid 2–related factor 2 (Nrf2) is known as a key regulator that induces cytoprotective gene expression (Zhang 2025). Under basal conditions, Nrf2 is sequestered by Keap1 and degraded, whereas oxidative stress enables its nuclear translocation and activation of antioxidant response element–dependent transcription (Taguchi et al 2011).
Although conventional periodontal therapies remove pathogens and damaged tissue, they do not directly restore the intrinsic metabolic dysfunction of chronically inflamed periodontal cells (Sculean et al 2008; Deas et al 2016; Graziani et al 2017). When cellular resilience remains compromised, regenerative outcomes may be inherently limited (Dabra et al 2012). Therapeutic strategies that reestablish cellular homeostasis before regenerative intervention may improve periodontal repair. According to previous studies, copine-7 (CPNE7) can contribute to cellular homeostasis under oxidative and metabolic stress (Hwang et al 2023; Lee et al 2024). In Cpne7 knockout (KO) mice, premature aging phenotypes have been observed in the dental pulp complex, including reduced pulp space, diminished cell numbers, increased apoptosis and DNA damage, and mitochondrial dysfunction under systemic stress (Hwang et al 2023; Lee et al 2024). In contrast, treatment with recombinant CPNE7 or its derivative peptide selcopintide (SCPT) has been reported to attenuate these pathological changes (Park et al 2021; Hwang et al 2023; Lee et al 2024). SCPT is a 10-amino acid sequence derived from the linker region between the C2 and von Willebrand factor type A domains of CPNE7, selected for its basic amino acids that confer cell-penetrating properties while retaining the biological activity of the full-length protein. SCPT has shown regenerative potential in periodontal defect models (Bai et al 2022; Jung et al 2025; Ko et al 2025). However, these findings derive from mechanically induced defects, and mechanistic insights under inflammatory conditions remain limited (Kim et al 2025).
In this study, we investigated the therapeutic mechanisms of SCPT in periodontitis by examining its ability to restore impaired cellular homeostasis under inflammatory conditions. Rather than targeting ROS or inflammatory mediators alone, we focused on the integrated regulation of redox balance, mitochondrial function, and stress-response signaling in resident periodontal cells. While previous studies primarily focused on structural regeneration, the present study addresses the mechanistic restoration of cellular homeostasis under inflammatory stress. We used wild-type (WT), Cpne7-KO, and overexpressing transgenic (Tg) mice to determine whether CPNE7 modulates susceptibility to periodontitis and evaluated the protective effects of SCPT under inflammatory conditions.
Materials and Methods
Animals
The study complied with the ARRIVE 2.0 (Animal Research: Reporting of In Vivo Experiments) guidelines. Details are provided in the Appendix.
Results
CPNE7 Expression Is Required for the Maintenance of Periodontal Integrity and Redox Homeostasis
To investigate CPNE7 expression in periodontal tissues during aging, we performed immunohistochemistry (IHC). In 3-mo-old mice, CPNE7 was prominently expressed in the PDL, cementum, and alveolar bone matrix. However, its expression progressively decreased with aging (Fig. 1A). In parallel, Nrf2 expression also decreased in periodontal tissues during aging (Appendix Fig. 1A). To assess the role of CPNE7 in maintaining periodontal integrity, we analyzed periodontal tissues from 24-mo-old WT, Cpne7-KO, and -Tg mice. KO mice exhibited narrow PDL space, disorganized fiber alignment, and cementum resorption, whereas Tg mice showed preserved fiber architecture and newly formed cementum (Fig. 1B). To assess the relevance of CPNE7-derived SCPT in cementum-related cells, we examined its effects on the cementoblast cell line OCCM-30. Treatment with SCPT enhanced cementogenic differentiation, as evidenced by increased Alizarin Red S staining and alkaline phosphatase expression (Appendix Fig. 2A, B). Given that ROS accumulation contributes to tissue damage, ROS levels were assessed in PDL tissues from 6- and 24-mo-old mice. KO mice exhibited higher ROS levels than WT and Tg mice in both age groups (Fig. 1C). These data indicate that CPNE7 is an important regulator of periodontal tissue structure and redox homeostasis during aging.
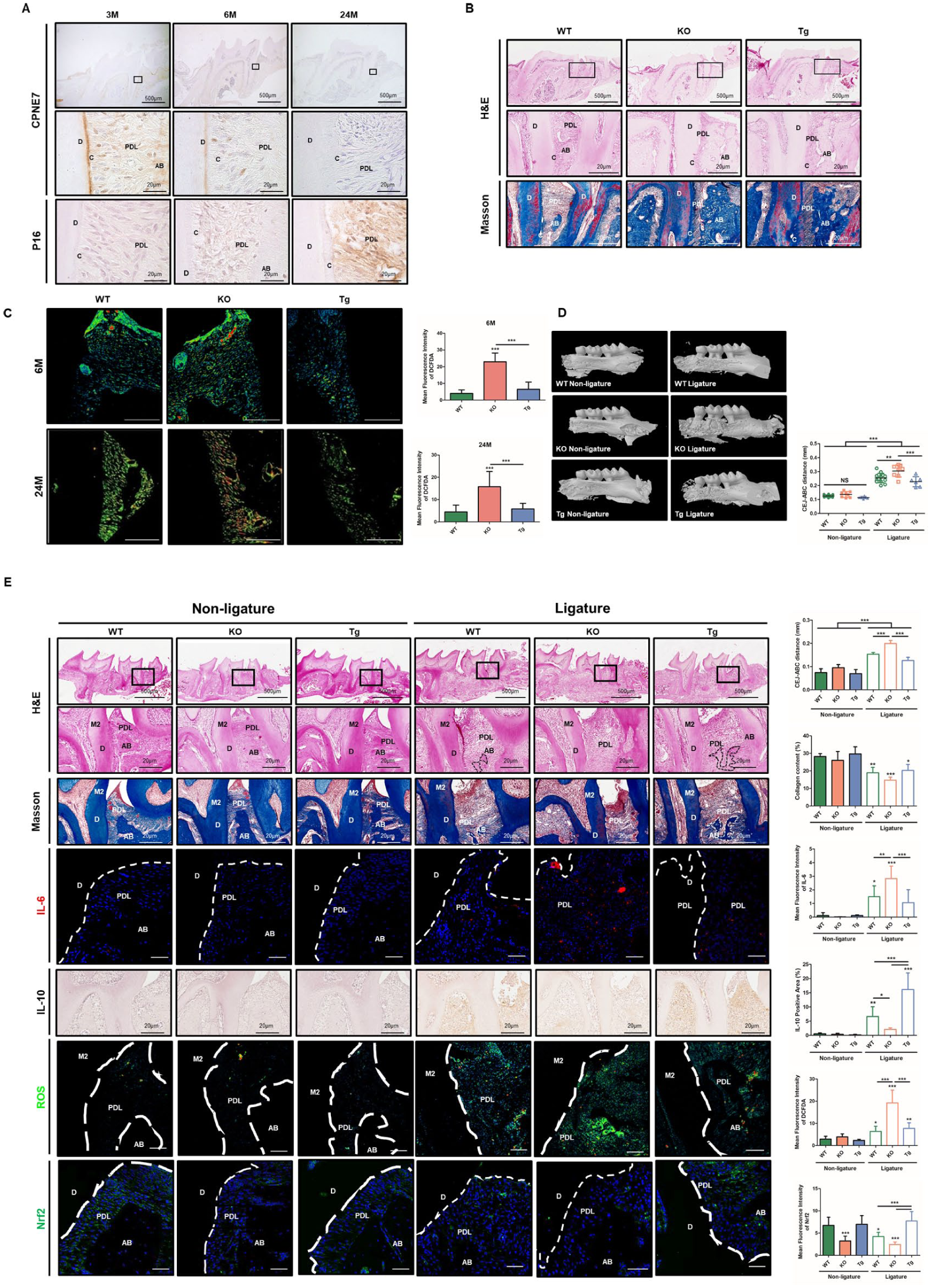
CPNE7 contributes to periodontal homeostasis and modulates periodontitis severity. (
CPNE7 Modulates the Severity of Periodontitis
Given that CPNE7 contributes to the maintenance of periodontal structure and function during aging, we next examined whether CPNE7 affects periodontal tissue integrity under inflammatory conditions. Experimental periodontitis was induced by ligature placement for 7 d, followed by micro-computed tomography (CT) and histological analysis. Under non-ligature conditions, alveolar bone levels were comparable among WT, KO, and Tg mice (Fig. 1D). However, after ligature application, KO mice exhibited aggravated bone loss, whereas Tg mice showed significantly reduced bone resorption relative to WT and KO. Consistent with these findings, hematoxylin and eosin (H&E) and Masson’s trichrome staining confirmed severe alveolar bone loss in KO mice and attenuated periodontal tissue destruction in Tg mice (Fig. 1E). To further investigate the underlying regulatory mechanisms, IHC was performed. KO mice demonstrated elevated IL-6, reduced IL-10, increased ROS accumulation, and diminished Nrf2 expression. In contrast, Tg mice exhibited enhanced anti-inflammatory and antioxidant responses, with increased IL-10, reduced ROS, and elevated Nrf2 expression. These data indicate that CPNE7 modulates the severity of periodontitis by regulating inflammatory responses and redox homeostasis.
SCPT Modulates Redox Balance by Activating Nrf2 under Inflammatory Conditions
In the periodontitis microenvironment, excessive ROS production disrupts cellular redox balance, leading to oxidative stress, tissue destruction, and impaired regenerative capacity (Chapple and Matthews 2007). Therefore, we investigated whether SCPT modulates ROS accumulation and maintains redox balance in human periodontal ligament cells (hPDLCs). To evaluate the antioxidant effects of SCPT, hPDLCs were treated with SCPT (100 μg/mL) and Porphyromonas gingivalis lipopolysaccharide (LPS, 5 μg/mL) for 48 h. Furthermore, to examine the preventive and therapeutic effects of SCPT, hPDLCs were pretreated with either SCPT or LPS for 24 h prior to subsequent LPS or SCPT exposure, respectively. Intracellular ROS levels increased after LPS stimulation (Fig. 2A). However, both pre- and post-SCPT treatments reduced ROS accumulation. Because mitochondria are the primary source of intracellular ROS, we next examined mitochondrial-specific ROS (mtROS) using MitoSOX staining. MitoSOX staining revealed that LPS-induced mtROS was attenuated by both pre- and post-SCPT treatments (Fig. 2B). Given SCPT’s ability to modulate intracellular and mitochondrial ROS, we examined whether the Nrf2 pathway mediates its antioxidant effects. Upon activation, Nrf2 dissociates from Keap1 and translocates into the nucleus to induce the expression of antioxidant genes. Indeed, SCPT treatment increased nuclear translocation and phosphorylation of Nrf2 in hPDLCs, accompanied by a reduction in Keap1 expression, collectively indicating activation of the Nrf2-Keap1 pathway (Fig. 2C, D). To determine whether SCPT-induced Nrf2 activation leads to antioxidant responses, we examined the expression of canonical Nrf2 target genes, including NQO1, HO-1, and SOD1, as well as the mitochondrial antioxidant enzyme SOD2. SCPT treatment increased HO-1 and SOD2 expression; however, NQO1 and SOD1 were upregulated following LPS stimulation, indicating that SCPT can strengthen basal antioxidant capacity and selectively enhance antioxidant defenses under inflammatory conditions (Fig. 2E). To examine whether the antioxidant effect of SCPT depends on Nrf2 activity, we treated hPDLCs with ML385 (5 μM), an inhibitor of Nrf2 transcriptional activity. SCPT increased Nrf2 phosphorylation (Appendix Fig. 3A). However, NQO1 induction by SCPT was not observed in the presence of ML385. Consistently, the reduction in intracellular ROS by SCPT was abolished when ML385 was coadministered (Appendix Fig. 3B). These results suggest that SCPT modulates cellular redox homeostasis through Nrf2 activation and the induction of antioxidant genes.
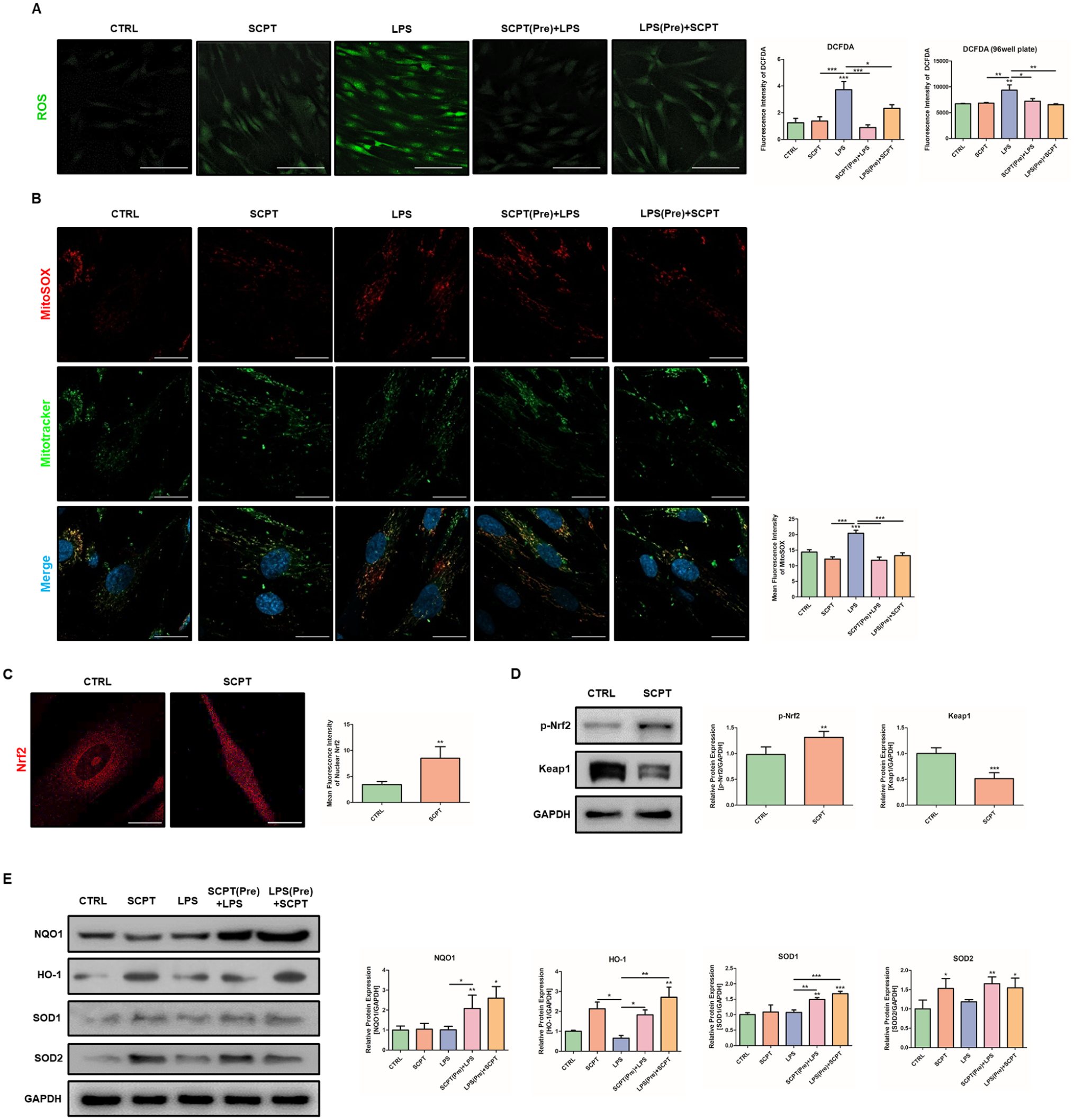
Selcopintide (SCPT) modulates redox balance through Nrf2 activation under inflammatory conditions. Human periodontal ligament cells (hPDLCs) were treated with SCPT and/or lipopolysaccharide (LPS) for 48 h. In pretreatment experiments, cells in the SCPT pretreatment group were treated with SCPT for 24 h, followed by LPS treatment for an additional 24 h. Cells in the LPS pretreatment group were treated with LPS for 24 h, followed by SCPT treatment for an additional 24 h. (
SCPT Restores Mitochondrial Function under Inflammatory Conditions
To examine whether SCPT reverses LPS-induced mitochondrial dysfunction, mitochondrial morphology was assessed using MitoTracker staining. LPS increased fragmented and rounded mitochondria, whereas pre- and post-treatment with SCPT reduced fragmentation and restored branched network structures (Fig. 3A). We next evaluated mitochondrial biogenesis-related markers. SCPT alone did not alter PGC-1α expression, but in the presence of LPS, SCPT increased PGC-1α levels. TOM20 and COX IV expression were also restored by SCPT under inflammatory conditions (Fig. 3B). Consistently, LPS-induced alterations in mtDNA copy number were restored by SCPT treatment (Fig. 3C). LPS reduced mitochondrial membrane potential (MMP), confirming mitochondrial damage. Both pre- and posttreatment with SCPT prevented this loss of MMP (Fig. 3D). Finally, mitochondrial respiration was evaluated using Seahorse analysis. LPS decreased basal respiration, maximal respiration, and ATP production, indicating impaired mitochondrial function (Fig. 3E). However, despite the recovery of mitochondrial morphology and function, SCPT did not alter the expression of dynamics-regulatory proteins across time points (6, 12, 24, and 48 h) following LPS stimulation (Appendix Fig. 4A). These findings suggest that SCPT-mediated mitochondrial recovery may occur without overt modulation of canonical fusion–fission regulatory proteins, as structural and functional restoration can precede measurable changes in these regulators. To determine whether the mitochondrial protective effects of SCPT are associated with Nrf2 activity, hPDLCs were cotreated with the Nrf2 inhibitor ML385. The restoration of mitochondrial morphology and membrane potential by SCPT under LPS stimulation was not observed in the presence of ML385 (Appendix Fig. 5A, B). Similarly, the improvements in basal respiration, maximal respiration, and ATP production were not maintained when Nrf2 activity was inhibited (Appendix Fig. 5C). These findings indicate that SCPT-mediated mitochondrial recovery under inflammatory stress is associated with Nrf2 activity.
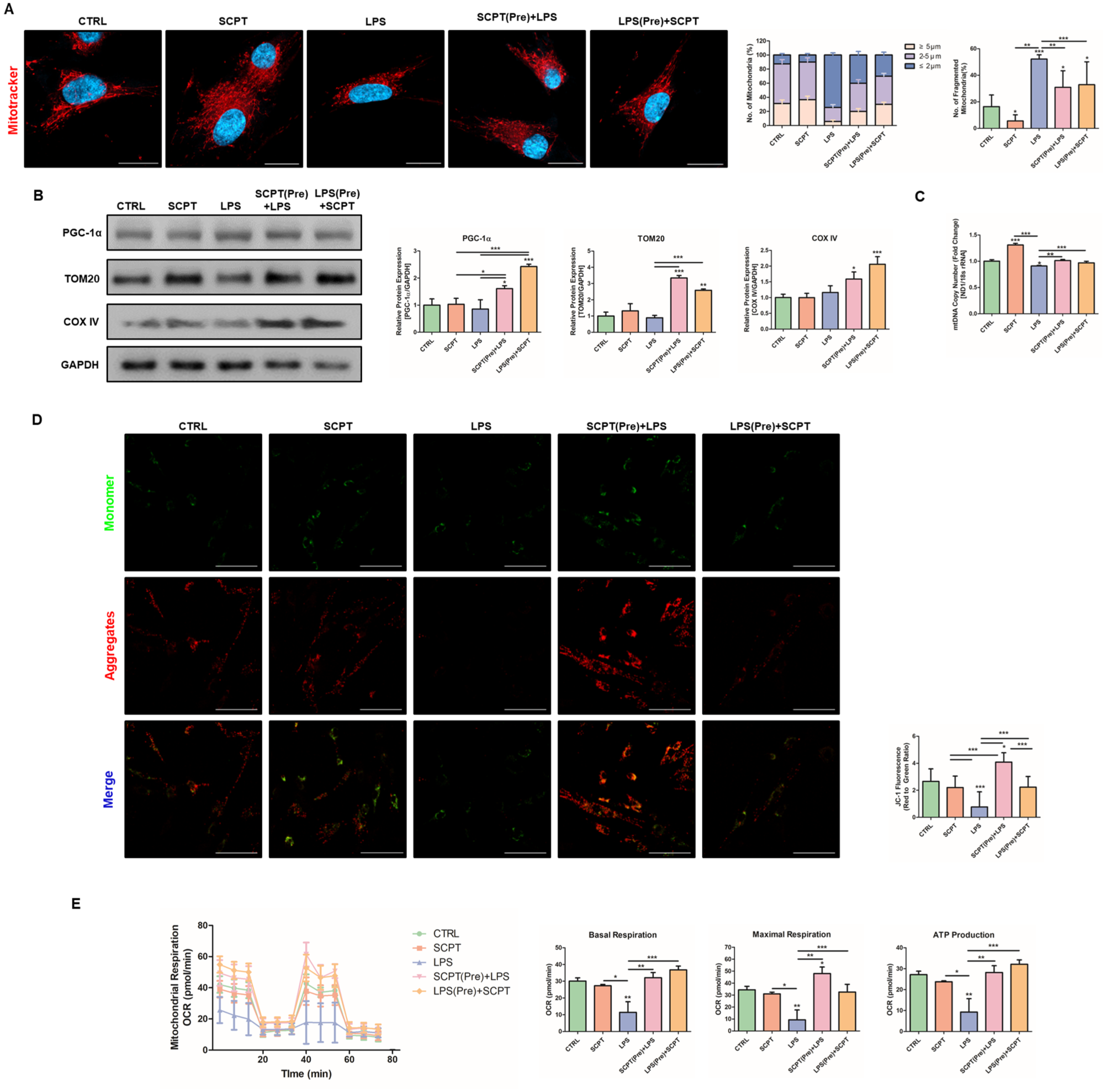
Selcopintide (SCPT) restores mitochondrial function under inflammatory conditions. Human periodontal ligament cells (hPDLCs) were treated with SCPT and/or lipopolysaccharide (LPS) for a total of 48 h. In pretreatment experiments, cells in the SCPT pretreatment group were treated with SCPT for 24 h, followed by LPS treatment for an additional 24 h. Cells in the LPS pretreatment group were treated with LPS for 24 h, followed by SCPT treatment for an additional 24 h. (
SCPT Suppresses LPS-Induced Inflammatory Responses and Apoptotic Signaling
Excessive inflammation and apoptosis are features of periodontitis (Meyle and Chapple 2015). Therefore, we next examined whether SCPT attenuates LPS-induced inflammatory and apoptotic responses in hPDLCs. NF-κB is a central mediator of LPS-induced inflammatory signaling, and its activation involves nuclear translocation. To determine whether SCPT modulates this pathway, we first examined NF-κB localization by immunofluorescence. LPS stimulation increased the nuclear translocation of NF-κB, but not by SCPT pretreatment (Fig. 4A). Consistently, SCPT decreased phosphorylated NF-κB expression levels (Fig. 4B). We next evaluated the expression of inflammatory cytokines. LPS increased the messenger RNA (mRNA) levels of TNF-α, IL-1β, and IL-6, whereas SCPT treatment suppressed their induction (Fig. 4C). To assess the effects of SCPT on apoptosis, Western blot and flow cytometry were performed. LPS stimulation increased the proapoptotic protein Bax and decreased the antiapoptotic protein Bcl-2, both of which were reversed following SCPT treatment (Fig. 4D). Likewise, mRNA expression of Bax was decreased by SCPT (Fig. 4E). Consistently, LPS increased the proportion of apoptotic cells, while SCPT reduced apoptosis under inflammatory conditions (Fig. 4F). In addition, LC3 expression was increased in SCPT combined with LPS treatment groups, suggesting that SCPT enhanced autophagic activity under inflammatory stress (Fig. 4G). To determine whether this autophagic response involves mitochondrial turnover, mitophagy was assessed using a mitophagy detection kit. Under inflammatory conditions, mitophagy intensity was increased by SCPT treatment (Fig. 4H). These findings indicate that SCPT suppresses NF-κB activation, reduces inflammatory cytokine expression, attenuates apoptosis, and may enhance mitophagy under inflammatory stress.
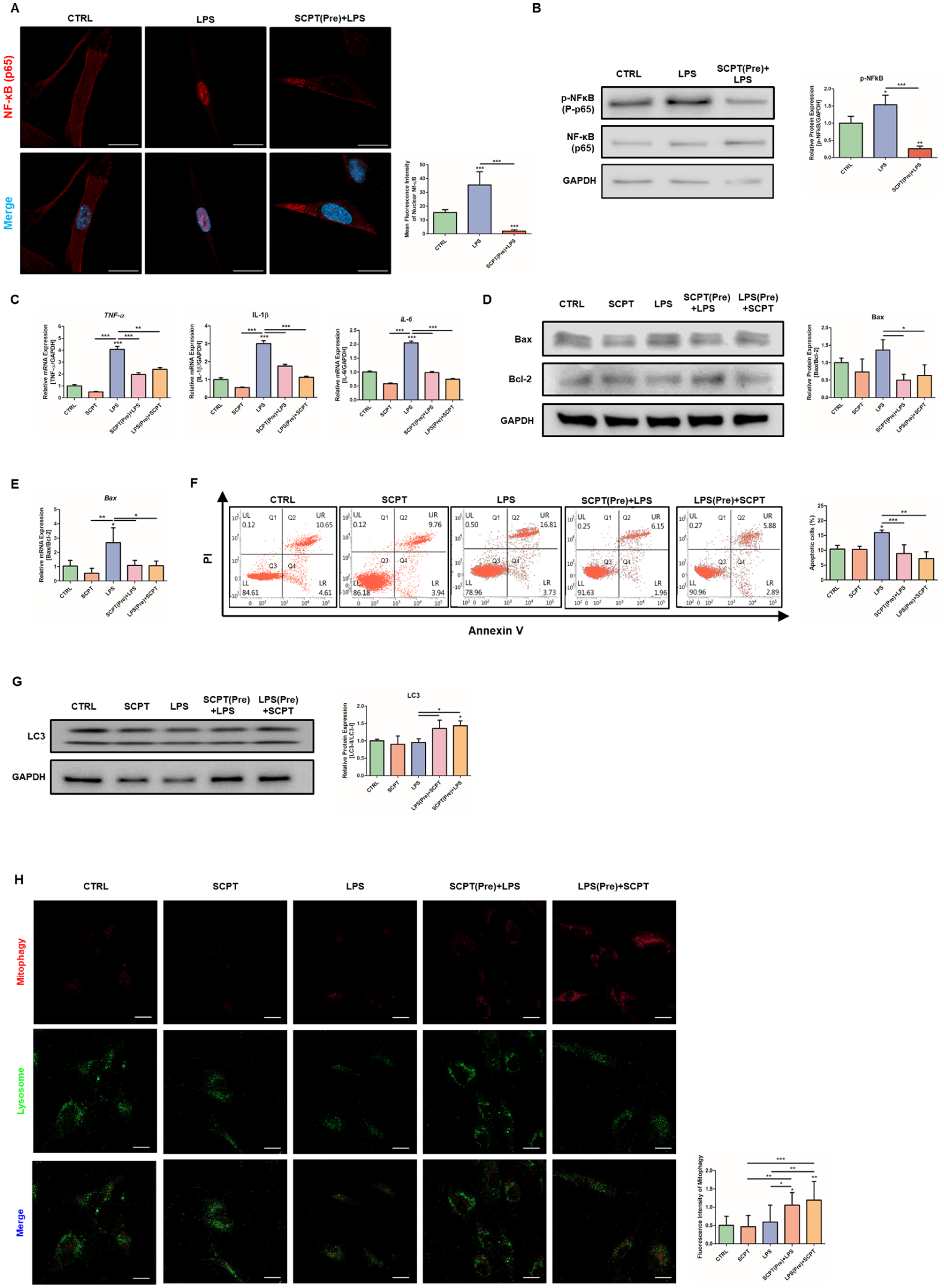
Selcopinetide (SCPT) suppresses lipopolysaccharied (LPS)-induced inflammatory responses and apoptotic signaling. Human periodontal ligament cells (hPDLCs) were treated with SCPT and/or LPS for 48 h. In pretreatment experiments, cells in the SCPT pretreatment group were treated with SCPT for 24 h, followed by LPS treatment for an additional 24 h. Cells in the LPS pretreatment group were treated with LPS for 24 h, followed by SCPT treatment for an additional 24 h. (
Therapeutic Effects of SCPT in a Periodontitis Mouse Model
Given the effects of SCPT observed in vitro, we next investigated whether SCPT exerts therapeutic effects in the ligature-induced periodontitis model. SCPT (50 μg/20 μL) or vehicle control (1.5% carboxymethyl cellulose, CMC) was applied topically for 7 d (Fig. 5A). Micro-CT analysis revealed that SCPT treatment attenuated alveolar bone loss compared with the ligature-only group (Fig. 5B). Histological analysis demonstrated that SCPT reduced bone resorption and PDL destruction, as shown by H&E and Masson staining (Fig. 5C). Tartrate-resistant acid phosphatase (TRAP) and TdT-dUTP terminal nick-end labeling staining showed that the numbers of osteoclasts and apoptotic cells were reduced in SCPT-treated tissues (Fig. 5D). To examine whether the reduction in osteoclast numbers was due to a direct effect on osteoclast differentiation, osteoclast differentiation assays were performed. SCPT treatment and CPNE7 expression did not significantly affect osteoclast differentiation in vitro or osteoclast-related markers in aged periodontal tissues (Appendix Fig. 6A–C). We next examined whether SCPT influences osteoblast differentiation. BSP expression, reduced in the ligature group, was restored by SCPT treatment in vivo (Appendix Fig. 7A). However, SCPT did not significantly affect mineralized nodule formation or osteogenic marker expression in MC3T3-E1 cells under osteogenic conditions (Appendix Fig. 7B, C). These results suggest that SCPT does not directly promote osteoblast differentiation in vitro. Inflammatory cytokine staining indicated that SCPT decreased IL-6 expression while increasing the anti-inflammatory cytokine IL-10 (Fig. 5E). To determine whether the anti-inflammatory effects of SCPT extend to immune cells, neutrophil activity was assessed by staining for neutrophil elastase (NE) and citrullinated histone H3 (CitH3). Neutrophil infiltration was increased in the ligature-only group and reduced following SCPT treatment (Appendix Fig. 8A). The number of NETosis-positive cells was also elevated in the ligature group and decreased following SCPT treatment. We examined whether the antioxidant mechanisms observed in vitro were present in vivo. ROS levels were decreased in the SCPT-treated group (Fig. 5F). Nrf2 expression, which was reduced in the ligature-only group, was restored following SCPT treatment. In parallel, the downstream antioxidant genes were upregulated in the SCPT-treated group. These results indicate that SCPT attenuates oxidative stress and inflammation in periodontal tissues in vivo, accompanied by Nrf2-dependent antioxidant pathway activation.
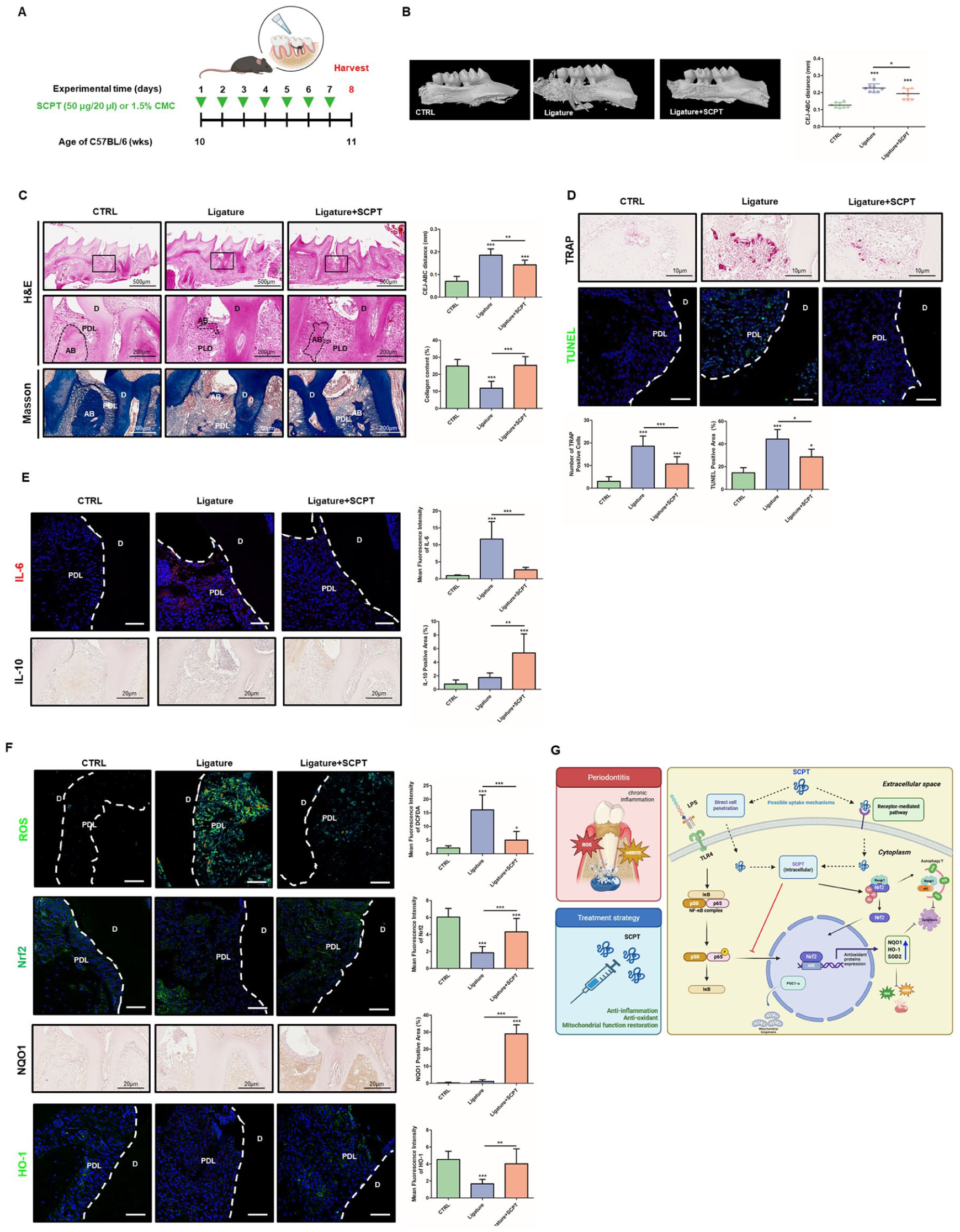
Therapeutic effects of selcopintide (SCPT) in a periodontitis mouse model. Mice at 10 wk old underwent ligature-induced periodontitis for 7 d and were sacrificed on day 8. SCPT or vehicle was topically applied once daily. (
Discussion
Periodontitis is a chronic inflammatory disease whose prevalence increases with age. Increasing evidence suggests that redox imbalance and mitochondrial dysfunction contribute to periodontal tissue deterioration, and aging further exacerbates this vulnerability through declining antioxidant defenses (Deng et al 2024). Current therapeutic strategies focus on pathogen elimination, inflammation control, and regenerative procedures to restore alveolar bone (Slots 2017; Chavda and Levin 2018; Kwon et al 2021). However, these approaches do not directly address intrinsic dysfunction of periodontal cells exposed to inflammatory stress. When cellular metabolic capacity and regenerative potential are compromised, suppression of inflammation alone may not be sufficient to restore tissue integrity (Eming et al 2017).
In the present study, we demonstrated that CPNE7 is an important regulator of periodontal tissue homeostasis under aging- and inflammation-associated stress, and that SCPT exerts a protective effect by restoring redox balance and mitochondrial function. Although previous studies have shown that SCPT promotes periodontal regeneration in artificial defect models (Bai et al 2022; Lee et al 2024; Jung et al 2025; Ko et al 2025), the underlying intracellular mechanisms of SCPT under pathological conditions remain unclear. In fact, CPNE7 expression was reduced, accompanied by increased oxidative stress and decreased Nrf2 expression. Similarly, Cpne7 deficiency led to PDL disorganization, increased cementum resorption, and ROS accumulation. Moreover, in the Cpne7 KO periodontitis model, alveolar bone loss and inflammatory responses were further aggravated, suggesting that CPNE7 contributes to periodontal homeostasis under stress conditions, and is closely associated with the Nrf2-mediated antioxidant pathway.
In this study, we stimulated hPDLCs with LPS to induce oxidative stress and mitochondrial dysfunction, characterized by increased intracellular and mitochondrial ROS, and impaired bioenergetics, consistent with previous reports demonstrating mitochondrial alterations in inflammatory periodontal cells (Li et al 2016). In contrast, SCPT treatment reduced ROS and mtROS and restored mitochondrial function. SCPT also increased SOD2 and HO-1, downstream antioxidant genes of the Nrf2 pathway, even in the absence of inflammatory stimulation. This suggests enhanced basal antioxidant capacity and improved resistance to oxidative stress, consistent with the established role of Nrf2 in antioxidant defense (Tonelli et al 2018). Notably, mitochondrial recovery occurred without detectable changes in mitochondrial dynamics proteins, whereas mitophagy activity increased following SCPT treatment. Previous studies have shown that mitochondrial recovery is driven by mitophagy-dependent clearance of damaged mitochondria (Twig and Shirihai 2011). Taken together, our results indicate that SCPT restores mitochondrial function by promoting mitochondrial quality control rather than altering mitochondrial dynamics under stress conditions.
In addition, local administration of SCPT attenuated alveolar bone loss, preserved PDL architecture, and reduced osteoclast numbers and apoptosis in the periodontitis model. Inflammatory cytokine profiles shifted toward a less inflammatory state, accompanied by reduced ROS levels and restoration of Nrf2 signaling in periodontal tissues. Consistently, ligature tissues exhibited reduced neutrophil infiltration and NETosis together with altered macrophage inflammatory cytokine expression following SCPT treatment. Although SCPT did not directly regulate osteoclast or osteoblast differentiation, reduced alveolar bone loss was still observed. These findings suggest that the bone-protective effect of SCPT is mediated indirectly through modulation of the inflammatory and oxidative microenvironment. This interpretation is consistent with previous studies demonstrating that regulation of the immune microenvironment can mitigate periodontal bone loss without directly targeting bone cells (Yao et al 2025).
In summary, CPNE7 contributes to periodontal homeostasis, and SCPT restores redox balance, mitochondrial function, and inflammatory regulation under stress conditions. These findings support the therapeutic potential of SCPT as a host-modulatory strategy for periodontitis. However, several limitations should be considered before clinical application. Although SCPT was found to restore cellular homeostasis through Nrf2-mediated signaling cascades, the mechanism underlying its internalization into hPDLCs remains unclear. Two possible pathways are suggested. First, since CPNE7 can bind to the cell surface receptor nucleolin (Seo et al 2017), SCPT may be internalized into hPDLCs via the receptor-mediated pathway. Alternatively, SCPT may enter hPDLCs via direct membrane translocation, as CPNE7-derived peptides exhibit cell-penetrating activity (Lee et al 2019). Further studies are required to clarify the precise internalization mechanism of SCPT. While SCPT-mediated protection was associated with Nrf2 activation and increased mitophagy, the upstream regulation of the Keap1-Nrf2 axis and the signaling pathways governing mitophagy are not yet fully understood. Cellular responses to SCPT may also vary depending on cell type, as previous studies have suggested stronger effects in certain mesenchymal lineage cells (Lee et al 2024). Moreover, the periodontitis model used in this study primarily reflects acute inflammatory tissue destruction and does not clearly distinguish inhibition of tissue breakdown from true periodontal regeneration. Therefore, further studies are needed to define the molecular mode of action, evaluate cell type-specific responses, and determine whether SCPT can promote periodontal regeneration in post-treatment models following ligature removal. Although we have confirmed the safety and therapeutic efficacy of SCPT delivered via an injectable hydrogel in clinically relevant canine periodontitis models (Kim et al 2025), further studies are required to bridge the translational gap between preclinical models and human application. Specifically, clinical trials are needed to establish safety, stability, optimal dosing, and long-term periodontal regenerative efficacy. Despite these limitations, SCPT may represent a promising therapeutic strategy for restoring periodontal homeostasis by alleviating cellular stress and stabilizing the periodontal microenvironment.
Author Contributions
H. Jang, J. Im, J.-C. Park, S.H. Han, contributed to conception and design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript; D.-S. Lee, contributed to conception and design, data acquisition and analysis, drafted and critically revised the manuscript; D.H. Park, C. Son, S.H. Ki, S. Park, S.M. Roh, J.-S. Seo, J.M. Park, contributed to design, data acquisition and analysis, critically revised the manuscript; O.-J. Park, contributed to design, data acquisition and interpretation, critically revised the manuscript; S. Jang, contributed to design, data analysis, critically revised the manuscript. All authors gave final approval and agreed to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345261450582 – Supplemental material for Restoring Cellular Homeostasis as a Therapeutic Strategy for Periodontitis
Supplemental material, sj-docx-1-jdr-10.1177_00220345261450582 for Restoring Cellular Homeostasis as a Therapeutic Strategy for Periodontitis by H. Jang, J. Im, D.-S. Lee, D.H. Park, C. Son, S.H. Ki, S. Park, S.M. Roh, J.-S. Seo, O.-J. Park, J.M. Park, S. Jang, J.-C. Park and S.H. Han in Journal of Dental Research
Footnotes
A supplemental appendix to this article is available online.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (Grant No. HI22C1684), National Research Foundation of Korea, which is funded by the Korean government (Grant No. RS-2022-NR0675350, RS-2022-00164722, RS-2025-00518511, and RS-2025-23324123).
ORCID iDs
Data Availability Statement
All data supporting the findings of this study are provided in the article and the Appendix files, with numerical source data underlying the figures publicly available in Figshare (DOI: 10.6084/m9.figshare.30995728).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.