
Abstract
Objective
Identify and compare phenotypic properties of osteoblasts from patients with otosclerosis (OSO), normal bones (HOB), and normal stapes (NSO) to determine a possible cause for OSO hypermineralization and assess any effects of the bisphosphonate, alendronate.
Study Design
OSO (n = 11), NSO (n = 4), and HOB (n = 13) cultures were assayed for proliferation, adhesion, mineralization, and gene expression with and without 10–10M-10–8M alendronate.
Setting
Academic hospital.
Methods
Cultures were matched for age, sex, and passage number. Cell attachment and proliferation + alendronate were determined by Coulter counting cells and assaying tritiated thymidine uptake, respectively. At 7, 14, and 21 days of culture + alendronate, calcium content and gene expression by quantitative reverse transcription-polymerase chain reaction (qRT-PCR) were determined.
Results
OSO had significantly more cells adhere but less proliferation than NSO or HOB. Calcification was significantly increased in OSO compared to HOB and NSO. NSO and HOB had similar cell adhesion and proliferation rates. A dose-dependent effect of alendronate on OSO adhesion, proliferation, and mineralization was found, resulting in levels equal to NSO and HOB. All cultures expressed osteoblast-specific genes such as RUNX2, alkaline phosphatase, type I collagen, and osteocalcin. However, osteopontin was dramatically reduced, 9.4-fold at 14 days, in OSO compared to NSO. Receptor activator of nuclear factor κB ligand/osteoprotegerin (RANKL/OPG), important in bone resorption, was elevated in OSO with decreased levels of OPG levels. Alendronate had little effect on gene expression in HOB but in OSO increased osteopontin levels and decreased RANKL/OPG.
Conclusions
OSO cultures displayed properties of hypermineralization due to decreased osteopontin (OPN) and also had increased RANKL/OPG, which were normalized by alendronate.
Introduction
Otosclerosis is a focal disease of the human temporal bone. 1 It is among the most common causes of acquired hearing loss in the United States, as it affects 0.1% to 1% of the general population. 2
Histologically, otosclerotic lesions in and around the stapes have 2 main phases: an early spongiotic phase (otospongiosis) and a late or sclerotic phase.3,4 Spongiotic bone has large marrow areas filled with fibrocytes and osteoclasts or acellullar cavitating areas, but most cells are from the osteoblast lineage.3,5 Remodeling occurs continuously within these foci, resulting in dense mineralized bone (sclerosis).6,7 Osteoblasts secrete osteoprotegerin (OPG), 8 which blocks receptor activator of nuclear factor κB ligand (RANKL) expressed by preosteoblasts. When RANKL binds to its receptor (RANK), osteoclast differentiation is triggered, leading to bone resorption. Therefore, stapes osteoblasts play a central role in all phases of otosclerosis. The developmental origin of the stapes bone is from the cranial mesenchyme of the second arch9-11 with osteoblasts being mesenchyme derived.
To investigate the pathophysiology of otosclerosis, various animal models have been proposed but do not adequately reproduce otosclerotic characteristics12,13 Cell culture has shown some promise as a model. Dost et al 14 was the first to culture stapes osteoblasts. Since that work, differences in sulfate uptake in otosclerotic stapes and renin-angiotensin-aldosterone system in otosclerotic patients were found,15-18 but despite these findings, no significant differences were observed in basic in vitro characteristics when otosclerotic stapes osteoblasts were compared to control tissue taken from the external auditory canal. 19 The inability to find differences may reflect the choice of control since both experimental and control tissue were from patients with otosclerosis. In addition, we have described the importance of controlling for age and sex on osteoblast properties in vitro. 20
Treatment of otosclerosis is primarily surgical, namely, stapedectomy. Bisphosphonates, the current treatment for osteoporosis, are the most promising pharmaceutical treatment for otosclerosis. 3 In a 2-year prospective double-blind study, investigators found a trend toward stabilization/improvement in hearing thresholds in otosclerotic patients treated with etidronate disodium. 21 A more recent clinical study supports the use of bisphosphonates to slow the progression of sensorineural hearing loss in otosclerotic patients. 22 In a retrospective study, patients diagnosed with otosclerosis were treated with etidronate, which appeared effective in the treatment of dizziness, hearing loss, and tinnitus. 23 OPG-knock-out mice, a proposed animal model for otosclerosis, treated with bisphosphonates had reduced otic capsule remodeling and hearing loss. 24 Bisphosphonates, however, are known for their antiresorptive properties, and it is not clear what mechanisms mediate all their beneficial effects in otosclerosis.
Given the central role of the osteoblasts in the pathophysiology of otosclerosis, we have described and characterized an in vitro model of otosclerotic osteoblasts to evaluate the effect of bisphosphonates.
Material and Methods
Cell Culture
Discarded stapes, including suprastructure and footplate fragments, from patients with otosclerosis (OSO; n = 11; 29F, 30F, 34F, 38F, 40M, 44F, 51F, 52M, 53M, 54M, 54F, describing age and sex) undergoing stapedectomy and normal stapes (NSO; n = 4; 40M, 42F, 50F, 60M) from patients undergoing translabyrinthine surgery were collected. No otic capsule samples were obtained (see appendix at www.otojournal.org). No identifiers were recorded other than age and sex. Stapedectomies were performed at the University of Connecticut Health Center, Yale-New Haven Hospital, and Hartford Hospital. Institutional Review Boards determined that our work did not constitute human subject research. Each stapes specimen was age- and sex-matched to a long bone specimen as close as possible in age and always same sex (n = 13; 29F, 30F, 34F, 40M, 41F, 46F, 46F, 52F, 52F, 52M, 53M, 54M, 54M). These human osteoblasts were labeled HOB as designated in the bone literature and were from discarded bone of healthy adults without history of hearing loss who underwent elective orthopedic procedures.
Specimens were morselized with a rongeur and cultured within 12 hours of collection in proliferation media; Dulbecco’s modified Eagle’s medium-F-12 (DMEM/F12 medium), 10% fetal bovine serum (FBS, Atlanta Biologicals, Inc., Lawrenceville, Georgia), 100U/ml penicillin-G and 100 μg/ml streptomycin (Invitrogen, Inc, Carlsbad, California). Once cells grew out, bone fragments were removed ( Figure 1 ). Cells were passaged once to evenly distribute them and then plated at a density of 10,000 cells/cm2 in 24-well plates (Costar, Corning Inc, Corning, New York) for subsequent experiments. Only passages 2 through 5 were used.
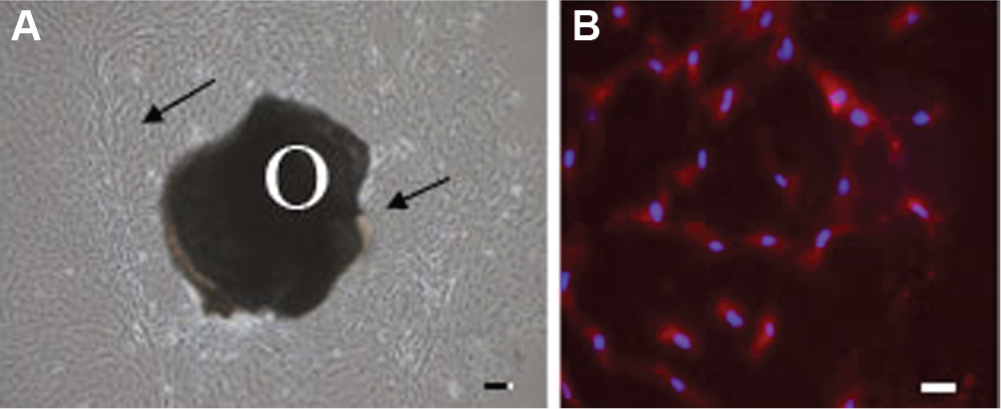
(A) Phase-contrast micrograph of cell outgrowth (arrows) from an otosclerotic footplate fragment (O) at 7 days. (B) Immunofluorescence micrograph demonstrating alkaline phosphatase staining at 8 days. Nuclei were stained blue with 1 μg/ml 4′,6-diamidino-2-phenylindole (DAPI). Bars = (A) 100 μm and (B) 10 μm.
For alendronate experiments, media with or without 10–10M-10–8M alendronate was added at 24 hours (Sigma, St. Louis, Missouri).
Alkaline Phosphatase Immunocytochemistry
Alkaline phosphatase (ALP) was examined in cell cultures that had been expanded as described previously but plated in 8-well chambers slides (Lab-Tek, Scotts Valley, California), cultured in proliferation medium for 3 days, and then switched to differentiation medium described in the following for 5 days. Slides were rinsed with PBS after each step. Cells were fixed, permeabilized with 0.1% Triton, and blocked with 4% bovine serum albumin (BSA). Cells were incubated with primary antibody at 4oC overnight, as well as control cells with blocking solution alone. The primary antibody was goat anti-rabbit polyclonal IgG to ALP (Santa Cruz Biotechnology, Inc, Santa Cruz, California, sc-30203), diluted 1:100. Then slides were incubated for 1 hour with 1:100 tetramethyl rhodamine isothiocyanate (TRITC)-labeled goat anti-rabbit IgG (sc-3841).
Adhesion Assay
After 7 days growth with and without alendronate, cells were replated. After 4 hours, cells were rinsed with phosphate buffered saline (PBS) to remove nonadherent cells, trypsinized, resuspended, and counted in a Beckman Coulter Z2 Counter (Beckman Coulter, Hialeah, Florida).
Proliferation Assay
After 68 hours, 5 μCi/ml of [3H]-thymidine (Perkin Elmer, Boston, Massachusetts) were added to each well. Four hours later, cells were washed with PBS and lysed with 0.5 N NaOH. Radioactivity was measured in lysates using a liquid scintillation counter (Packard Instrument Co, Downers Grove, Illinois); disintegrations per minute(dpm)/cell number at 72 hours of culture.
Mineralization
Cells were grown for 7 days in proliferation medium, which was changed to differentiation medium: alpha-MEM, 10% FBS, antibiotics, 2 mM β-glycerol phosphate with 100 μg/ml ascorbic acid. This time point was considered time 0. Calcium content was quantified on days 7, 14, and 21. Cells were treated with 5% trichloroacetic acid for 1 hour twice. Calcium content data (μg/ml) were determined in the combined supernatants using a standardized Calcium Kit (Eagle Diagnostics, De Soto, Texas) in a microtiter plate reader at 570 nm. To verify mineralization by staining calcium in the matrix, cells were fixed with 10% formalin, stained with 2% solution of alizarin red (Sigma), pH 4.2, washed, dried, and photographed.
Quantitative Reverse Transcription-Polymerase Chain Reaction
RNA was extracted at 7, 14, and 21 days in mineralization medium. Cells were pelleted and RNA was extracted using the RNeasy kit (Qiagen, Valencia, California). Cells were homogenized using QIA shredder columns (Qiagen). cDNA was synthesized using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, California) in a BioRad iCycler (Hercules, California). Ten μl RNA was added to the iCycler, which was programmed to 25°C for 10 minutes, 37°C for 120 minutes, and 85°C for 5 minutes. Resulting cDNA was used according to BioRad’s SYBR Green protocol in Applied Biosystems 7900HT. All samples were tested in triplicate and repeated in at least 4 independent experiments with separate patient samples. All samples were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Values in gene expression in OSO and HOB were expressed relative to NSO. Primer sequences are shown in Table 1 .
Forward and Reverse Primers Used in Quantitative Reverse Transcription-Polymerase Chain Reaction Experiments.

Abbreviations: RANK, receptor activator of nuclear factor kappa-B; RANKL, RANK ligand; GADPH, glyceraldehyde 3-phosphate dehydrogenase.
Western Blots
After 14 days in mineralization medium ±10–9M alendronate, cells were prepared for Western blot analysis. 25 Incubation with the first antibodies followed manufacturer’s instructions with rabbit anti-goat polyclonal IgG to osteopontin (OPN), GAPDH, RANKL, or OPG (sc-10591, sc-20358, sc-7627, sc-8468, respectively, Santa Cruz Biotechnology, Inc). The second antibody was 1:10,000 rabbit anti-goat HRP-conjugated antibody (sc-2768, Santa Cruz Biotechnology, Inc). Protein bands were visualized with ECL Western Blotting kit (Amersham, Piscataway, New Jersey). Band densities were assessed by Quantity One software (BioRad) and GAPDH was used to normalize OPG, OPN, and RANKL levels.
Statistics
Each experiment consisted of 3 groups: OSO, NSO, and HOB. Since surgeries for OSO and NSO samples were known ahead of time, HOBs matching in sex and similar age could be planned. Four samples (separate cell culture wells) per patient were analyzed in each assay, and at least 4 patients per group were analyzed in separate experiments for the adhesion, proliferation, and mineralization assays. The combined results for each patient had an N of 16 samples, on which statistical tests were performed. For quantitative reverse transcription-polymerase chain reaction (qRT-PCR), 1 RNA sample was extracted from 4 cell culture wells per patient to obtain enough RNA. Four patients were analyzed for each gene. One cDNA sample was synthesized from the RNA from each patient and then run in triplicate for each gene, resulting in an N of 4 for each gene. Statistical comparisons were carried out by nonparametric ANOVA (Kruskal Wallis) followed by Bonferroni posttest. All data are means ± standard deviations. Western blots were performed on 3 patients per group with comparable results. One representative Western blot is shown.
Results
Cells began to emerge from bone fragments in 1 to 2 weeks ( Figure 1A ). Approximately 6 million cells are obtained from the stapes of 1 patient. Immunofluorescence staining for the osteoblast marker, alkaline phosphatase, confirmed that 90% to 95% of the cells plated as described in Methods expressed high levels of alkaline phosphatase ( Figure 1B ).
To study adhesion and proliferation, control cultures from the stapes of normal patients (NSO) and normal human osteoblasts (HOB) were compared to cells from otosclerotic patients (OSO). A significant bone type effect was present in both adhesion and proliferation (ANOVA P < .001 for both, Figure 2 ). OSO adhered significantly more than NSO (P = .001) or HOB (P = .001). NSO and HOB showed no significant differences between each other. OSO exhibited significantly lower proliferation than NSO and HOB (P = .001), which were similar ( Figure 2B ).
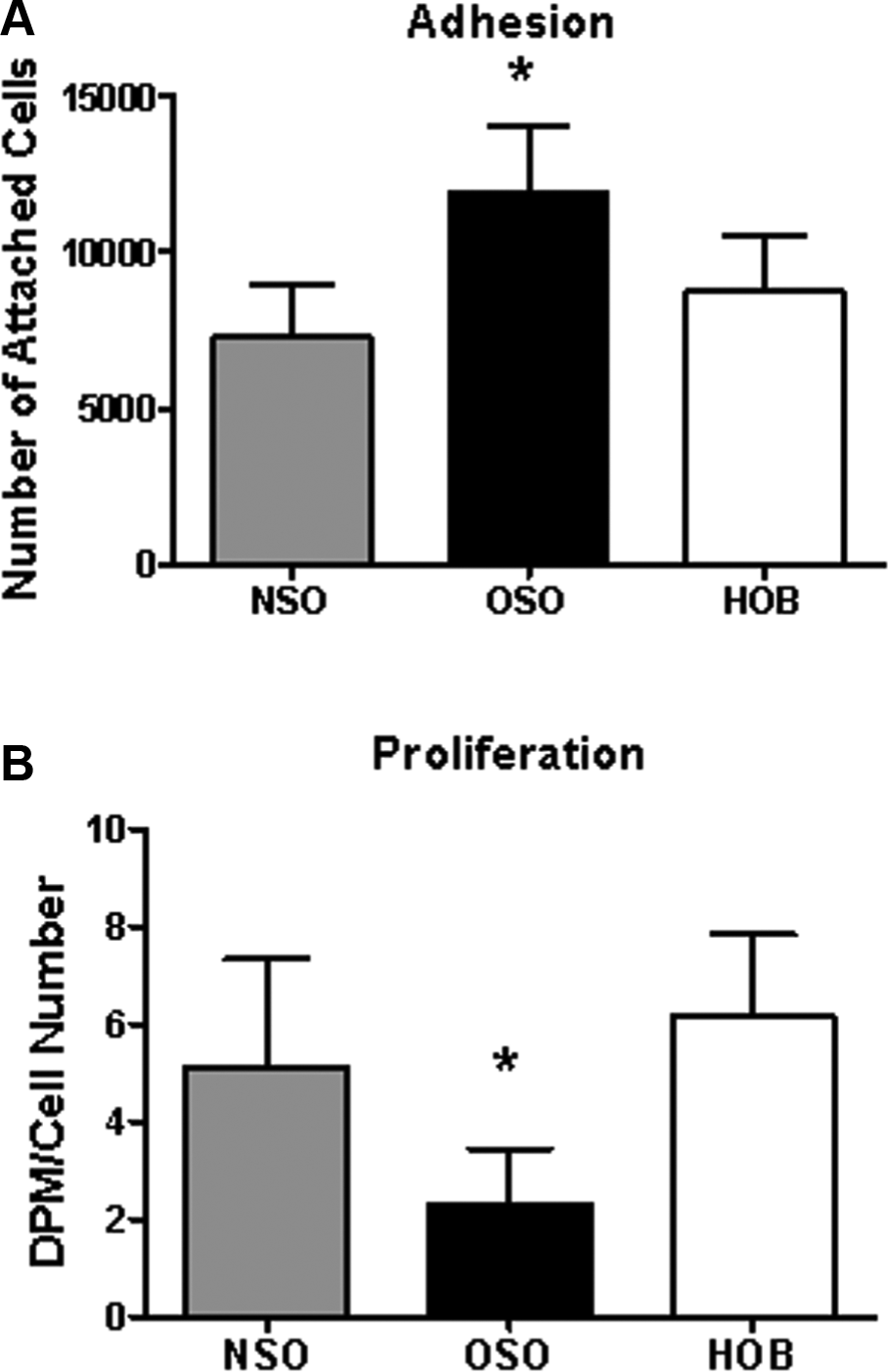
(A) NSO and HOB adhesion were not significantly different while OSO had significantly more cells attached (*P < .001). (B) Proliferation was comparable for NSO and HOB but OSO had a significantly lower proliferation rate (*P < .001). DPM, disintegrations/minute of [3H]-thymidine; NSO, normal patients; HOB, normal human osteoblasts; OSO, otosclerotic patients.
Mineralization was quantified on days 7, 14, and 21 ( Figure 3A ). ANOVA showed main effects of bone type (P = .03) and day (P < .001). Calcification significantly increased with time in all groups from 7 days to 21 days (P < .01). When comparing 7 versus 14 days, calcium content increased with time; however, it was not a statistically significant increase. Variability between patients was too great for significant differences within each group from 7 to 14 days, namely, 7 day NSO versus 14 day NSO, 7 day OSO versus 14 day OSO, and so on. However, at 14 days, OSO had significantly increased mineralization compared to NSO (P = .01) by 7.9-fold. At 21 days, OSO remained significantly different from NSO (P = .04). HOB mineralization was always intermediate in amount between OSO and NSO for all time points and was not significantly different from NSO calcification. Alizarin red staining confirmed increased calcification in OSO compared to NSO and HOB ( Figure 3B ). These results demonstrate more mineralization in OSO.
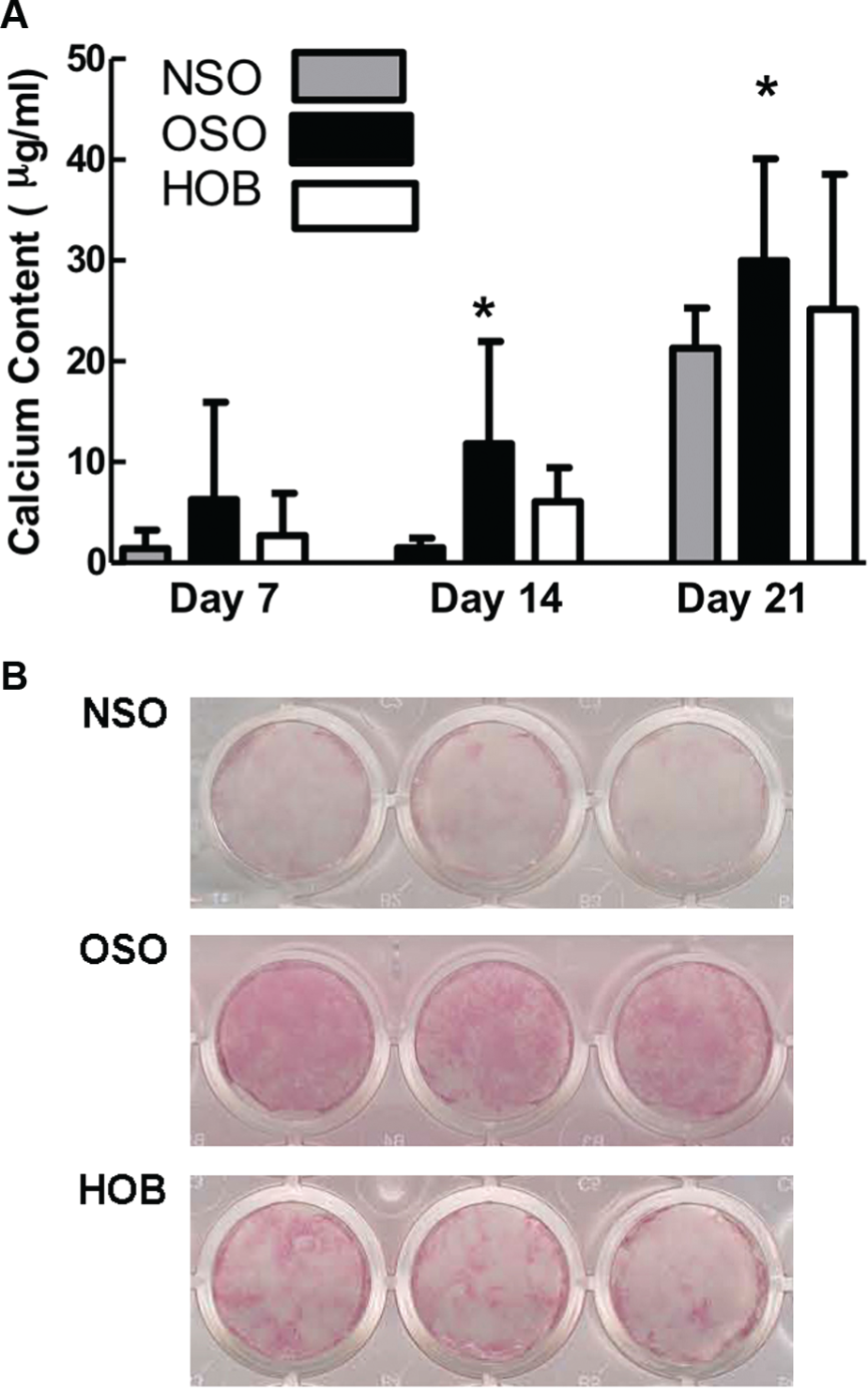
(A) Mineralization increased significantly with time. OSO was always significantly higher than NSO (*P < .01, 14 days; *P = .04, 21 days) but not to HOB at 21 days. (B) Alizarin red confirmed increased staining of the calcified cell layer in OSO compared to NSO and HOB in 3 tissue culture wells each. OSO, otosclerotic patients; NSO, normal patients; HOB, normal human osteoblasts.
Quantitative RT-PCR demonstrated that OSO, NSO, and HOB were osteoblast cultures, expressing bone-specific genes such as Runt-related transcription factor 2 (RUNX2), alkaline phosphatase, type I collagen, and osteocalcin ( Figure 4 ). RUNX2 gene expression was similar for all cultures. Since RUNX2 is a transcription factor that regulates cell commitment to the osteoblast lineage, these gene expression profiles suggest all cells were committed similarly to become osteoblasts. Alkaline phosphatase, reflective of osteoblast differentiation, was significantly higher in OSO compared to NSO on 7 and 21 days (P < .01). Type I collagen was significantly higher in OSO and HOB than NSO at 7 days (P = .04), but otherwise levels were similar. Osteocalcin was 24.1-fold greater in OSO than NSO at 14 days. At 21 days, osteocalcin increased to higher levels in NSO than OSO and HOB, which had decreased from 14 days. However, the absolute expression of osteocalcin was low, typical of human osteoblast cultures.26,27 One of the most striking findings was that osteopontin was significantly reduced in OSO compared to NSO at all time points with a maximal decrease of 9.4-fold at 14 days (P = .01). In addition, OSO had decreased levels of osteopontin compared to HOB at all time points. For confirmation, Western blots were performed to measure protein levels of osteopontin ( Figure 5 ). Although Western blots are semi-quantitative, osteopontin in OSO was decreased 56% compared to HOB.
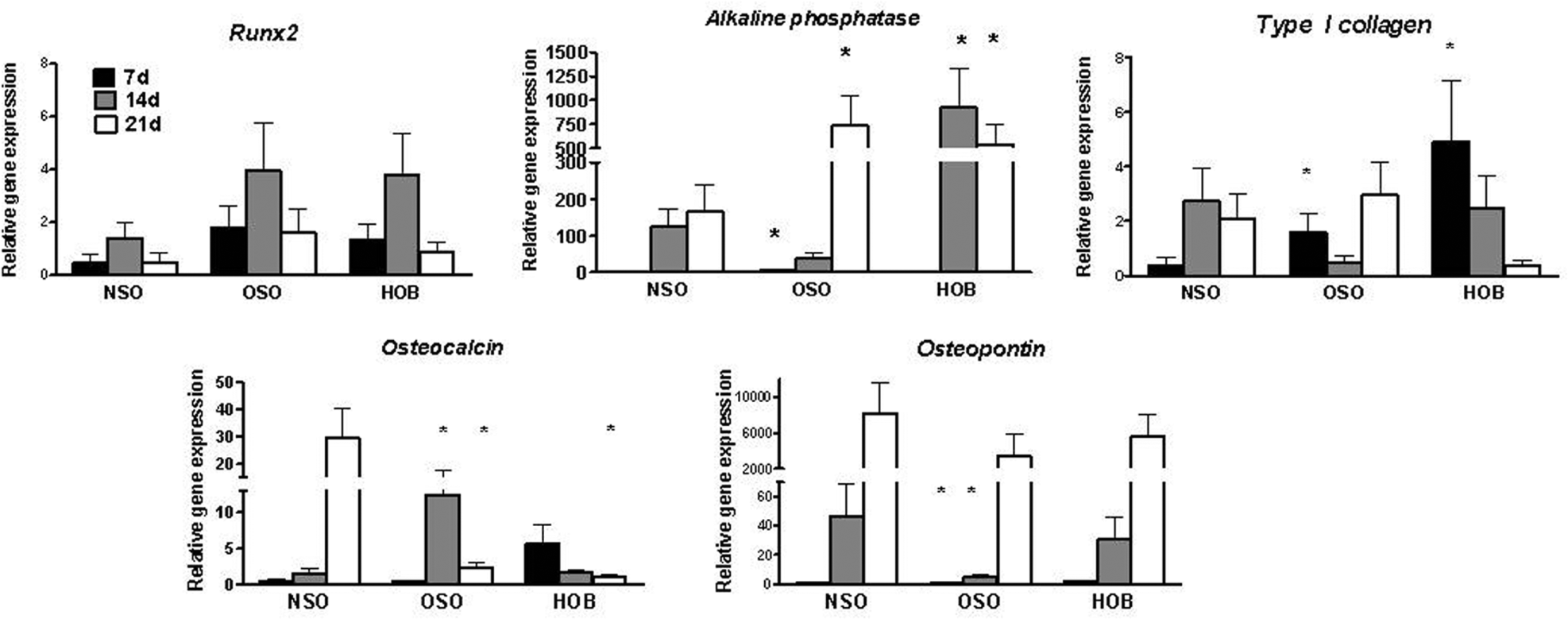
Message levels for RUNX2, alkaline phosphatase, type I collagen, osteocalcin, and osteopontin were determined by quantitative reverse transcription-polymerase chain reaction and normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) at 7, 14, and 21 days of culture. Data are shown as fold-difference relative to control NSO (normal patients). *P ≤ .04.
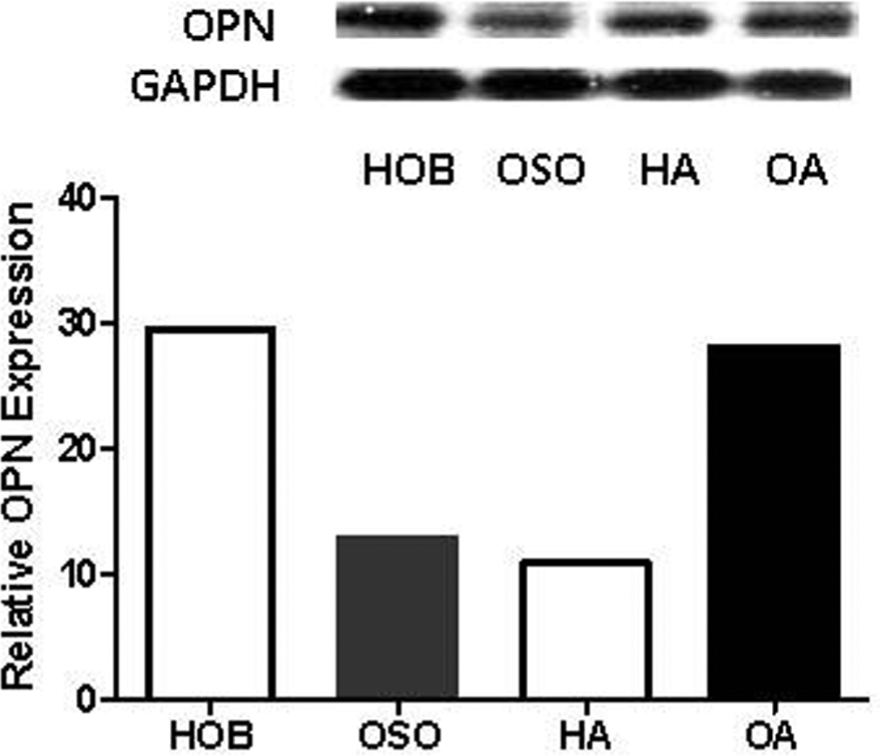
Western blot of osteopontin (OPN) at 14 days ± alendronate. OPN protein in OSO were decreased compared to HOB, and 10–9M alendronate increased OPN in OSO (OA) and decreased HOB OPN (HA). Densitometric analysis of bands normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) are graphed. OSO, otosclerotic patients; HOB, normal human osteoblasts.
Levels of RANKL and OPG gene expression were determined as an index of osteoclastogenic stimulus and bone resorption. 28 In NSO, OSO, and HOB, RANKL and OPG were low at 7 days, increased at 14 days and reduced to negligible levels by 21 days (data not shown). RANK was also measured, but no significant RANK expression was found. Compared to NSO, OSO and HOB had significantly lower OPG and higher RANKL expression at 7 days (ANOVA P = .001) ( Figure 6 ), suggesting increased bone resorption capacity.
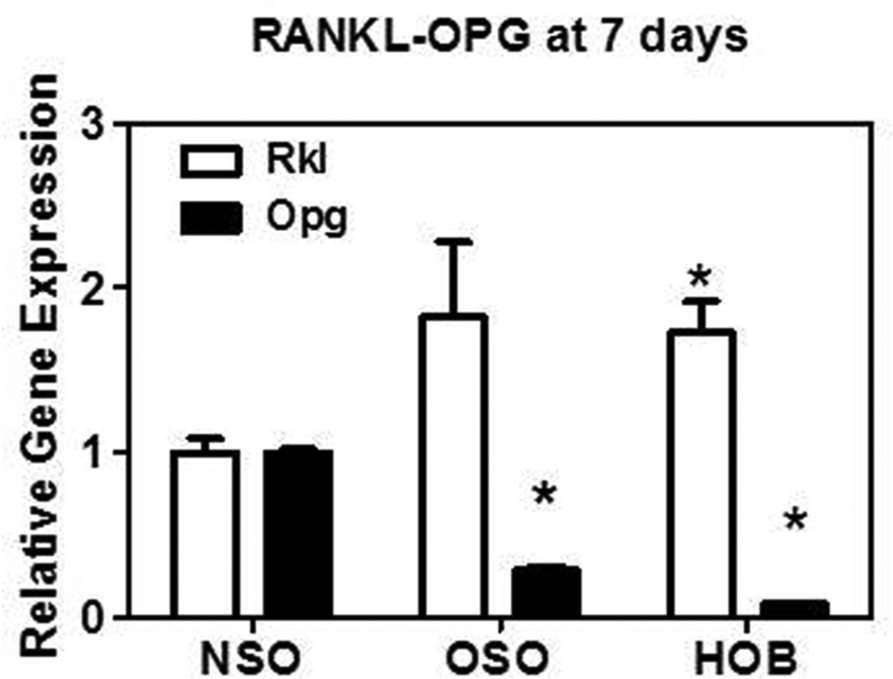
Quantitative RT-PCR. In OSO, OPG but not RANKL levels were reduced compared to NSO. In HOB, OPG and RANKL were similar to OSO. RANKL in HOB was 2-fold more than in NSO. *P < .001. RT-PCR, reverse transcription-polymerase chain reaction; OSO, otosclerotic patients; OPG, osteoprotegerin; RANKL, receptor activator of nuclear factor κB ligand; NSO, normal patients; HOB, normal human osteoblasts.
The effect of alendronate on OSO and HOB was investigated. Control OSO (no alendronate) had significantly higher cell adhesion (P = .01) than control HOB ( Figure 7A ), confirming Figure 2A . When alendronate was added to HOB, adhesion decreased in a dose-dependent manner, with a significant reduction at 10–9 M-10–8M alendronate (P = .03). When alendronate was added to OSO, cell attachment was significantly decreased at all 3 doses (P = .03) and no longer significantly different from HOB control (P = .9). Since the lowest concentration of alendronate has no significant effect on HOB cell attachment but had a significant effect on OSO, it appears that OSO were more responsive to alendronate than HOB.
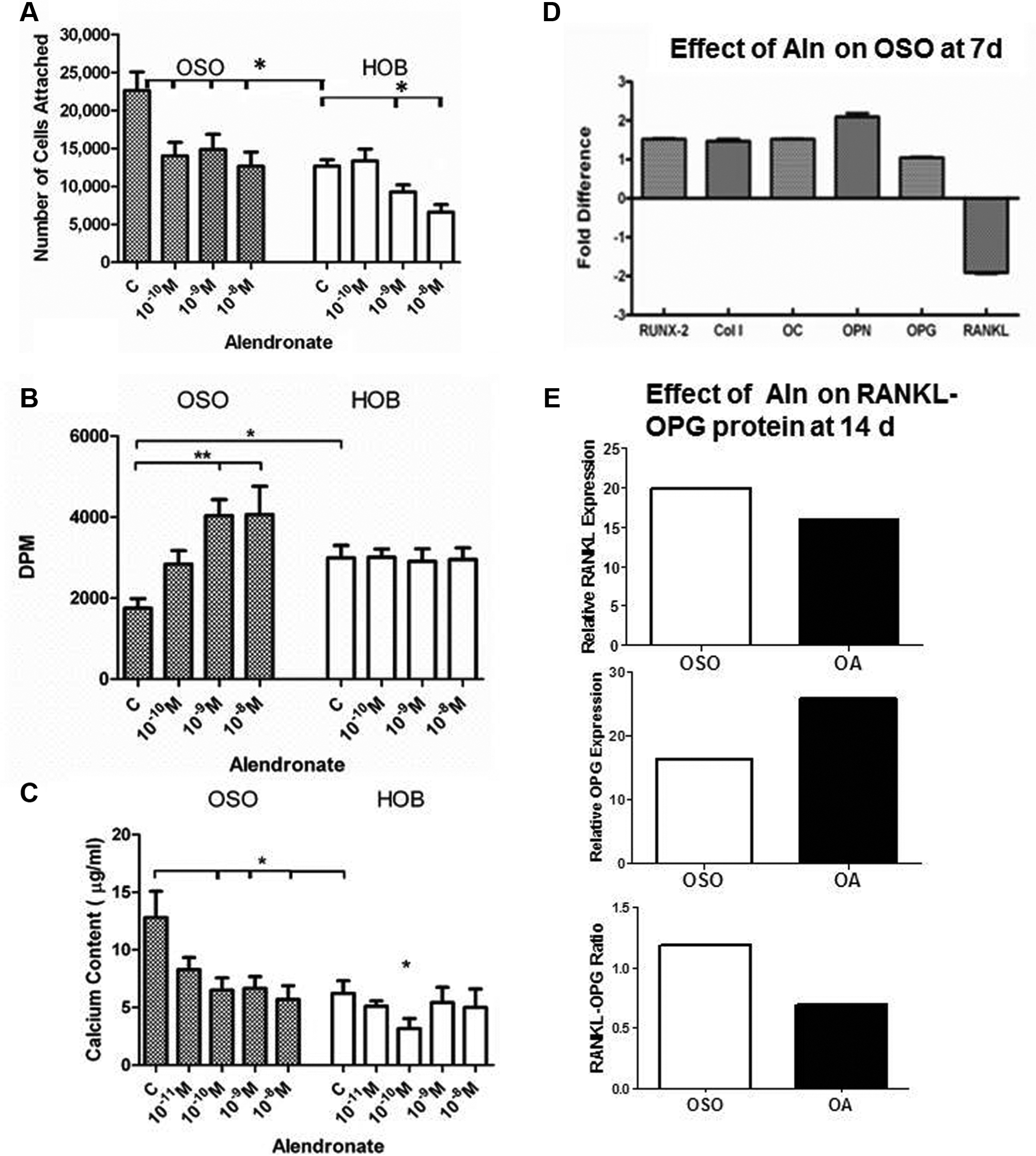
Alendronate (10–10M-10–8M) brought OSO (A) adhesion, (B) proliferation, and (C) calcium content to HOB levels with little effect on HOBs. Vehicle-treated cells are labeled as control (c). (D) qRT-PCR graphed as OSOuntreated/OSOtreated demonstrated that alendronate (10–9M) had no effect on RUNX2, collagen (Col I), or osteocalcin (OC) but affected OPN and RANKL. (E) Graphs of Western blots of OPG and RANKL at 14 days ± 10–9M alendronate in OSO, normalized to GAPDH. Alendronate increased OPG and decreased RANKL (OA), thereby decreasing the RANKL/OPG ratio, an index of bone resorption. OSO, otosclerotic patients; HOB, normal human osteoblasts; qRT-PCR, quantitative reverse transcription-polymerase chain reaction; OPN, osteopontin; RANKL, receptor activator of nuclear factor κB ligand; OPG, osteoprotegerin; NSO, normal patients. *P < .03. **P = .01.
No significant difference in proliferation was found between control and alendronate-treated HOB ( Figure 7B ). However, addition of 10–9M-10–8M alendronate to OSO increased proliferation significantly from control OSO (P = .01), and OSO no longer showed significant differences from control HOB (P = .92 and P = .96, respectively).
The effect of alendronate on bone formation in OSO and HOB were measured by assaying calcium content. Figure 7C demonstrates that without alendronate treatment, OSO were significantly more mineralized than HOB (P = .01) at 14 days, similar to Figure 3 . A dose-dependent decrease in calcium content was found in alendronate-treated OSO cultures with a significant decrease (P = .02) with 10–10M-10–8M alendronate. There was no significant effect of alendronate on HOB mineralization, except at 10–10M alendronate.
The effect of 10–9 M alendronate was examined on gene expression of OSO. Figure 7D shows that OSO had a small but insignificant increase in RUNX2, type I collagen, osteocalcin, and OPG. However, osteopontin, which was decreased in OSO compared to NSO and HOB ( Figure 4 ), was significantly increased 2.4-fold by alendronate. Alendronate dramatically decreased RANKL but had no significant effect on OPG. To determine protein expression, Western blots were performed on OSO at 14 days ( Figure 7E ). Alendronate increased OPG and slightly decreased RANKL. The RANKL/OPG ratio, an index of bone resorption, was decreased from 1.18 to 0.69 with alendronate, which would reduce bone resorption. Alendronate-induced changes in RANKL/OPG and osteopontin appear to normalize OSO osteoblast function.
Discussion
Osteoblasts cultured from stapes of patients with otosclerosis have characteristics that distinguish them from osteoblasts from normal stapes and peripheral bone. OSO displayed higher cell adherence, proliferated less, and were significantly more mineralized than HOB and NSO. Gene expression analysis demonstrated that OSO were deficient in osteopontin. These properties are consistent with mature otosclerotic lesions, which have increased mineralization and reduced cellularity.9,29
In the normal otic capsule, little bone remodeling occurs after development, but the stapes has the capacity to activate osteoblasts to proliferate and form new bone as seen in culture (NSO). NSO had a similar proliferative capacity as HOB. However, OSO had a significant reduction in proliferation, signifying that the otosclerotic osteoblasts are not easily renewed. Similarly, adhesion studies showed a significant phenotypic change. Osteoblasts adhere to the extracellular matrix via integrins.30-33 Significantly higher OSO adhesion suggests that expression of integrins differ from that of normal NSO and HOB and reflect differences in cell function since integrins mediate growth factor signaling, cell survival, and function. Adhesion and proliferation assays showed that OSO are abnormal in their responses while NSO behaved similar to HOB. Thus, human osteoblast cultures, matched by sex and age, appear to be a promising experimental model for studying the pathophysiology of otosclerosis.
Our results also suggest that osteopontin plays a role in the pathophysiology of otosclerosis, since it is significantly and consistently decreased in OSO compared to NSO and HOB, as determined by qRT-PCR and Western blots. Osteopontin is a 44 kD phosphorylated sialoprotein and member of the Small Integrin-Binding Ligand, N-linked Glycoprotein (SIBLING) family. 34 Osteopontin occurs in many different tissues and biological fluids, abundant in bone, produced by both osteoblasts and osteoclasts,35-37 and found in inner ear fluids. 38 The sites of osteopontin deposition are the cell-mineralization surfaces, osteoid, and cement lines. 39 Osteopontin is thought to play an important role in bone remodeling and in affecting mineral crystal size and bone mineral density with its deficiency causing increased mineral content. 40 It is an inhibitor of ectopic calcification.41-43 Our finding of decreased levels of osteopontin in OSO but otherwise similar levels of other genes compared to NSO and HOB suggests that osteopontin plays a role in hypermineralization that occurs in otosclerosis.
Since otosclerosis involves changes in bone remodeling, 44 the RANKL-OPG ratio was assessed. OPG levels were lower in OSO compared to NSO, which would promote bone resorption. Hypermineralization in enamel and dentin of OPG-knockout mouse teeth has also been reported. 45 OPG levels in the serum and by RT-PCR of bone have been found to be significantly lower in ankylotic stapes of patients with active disease as determined by histological examination, 46 which mirrors our results.
The anomalies in OPG and osteopontin levels in otosclerosis would contribute to hypermineralization and also point to a genetic component of the disease. There is strong evidence for a hereditary component with approximately 50% to 60% of affected persons reporting a positive family history 2 ; however, the penetrance and degree of expression is highly variable, making it difficult to detect inheritance patterns. This may be due to both genes needing to be decreased to produce otosclerosis. Gene expression analysis of 9 human otosclerotic stapes compared to 7 stapes from patients undergoing translabyrinthine surgery from vestibular schwannomas revealed differences in a number of genes, 47 but ostopontin levels were not analyzed. In particular, 2 families of genes involved in interleukin signaling and in inflammation were implicated, but no definitive reason for otosclerosis was determined.
A comparison of OSO and HOB has therapeutic relevance and is important in studies of drugs that alter bone turnover. Systemic administration of such drugs to patients with otosclerosis could alter properties of normal osteoblasts elsewhere in the skeleton. Bisphosphonates are analogs of the naturally occurring substance, pyrophosphate. Like pyrophosphate, bisphosphonates have a high affinity for calcium crystals and inhibit their growth and dissolution. Bisphosphonates decrease bone resorption and are resistant to biological degradation. Nitrogen-containing bisphosphonates, such as alendronate, inhibit the action of osteoclasts through inhibition of an enzyme, farnesyl pyrophosphate synthase, that is involved in the function and survival of osteoclasts. 48 In addition, bisphosphonates have been shown to affect osteoblasts by inhibiting apoptosis 49 and initiating differentiation. 50 Bisphosphonates also affect adhesion molecules, 51 a finding consistent with our finding that OSO adhesion was decreased with alendronate. In a model of vascular calcification in rats, alendronate was shown to increase osteopontin and OPG levels in a dose-dependent manner and to decrease calcification, 52 which is similar to our findings. Alendronate also reduced the RANKL/OPG levels in OSO, which would decrease bone resorption. In women taking alendronate, osteoblast cultures derived from their femoral bone marrow had decreased RANKL/OPG compared to women who had no history of taking bisphosphonates. 53 Bisphosphonates may specifically target otosclerotic osteoblasts and mitigate the deleterious aspects of their abnormal function.
Conclusions
Stapes osteoblasts cultured from patients with otosclerosis are significantly different from osteoblasts of normal patients’ stapes and peripheral bone in their ability to adhere, proliferate, and mineralize. In a dose-dependent manner, alendronate changed OSO so that they were not significantly different from NSO and HOB. In OSO, alendronate up-regulated osteopontin levels that were deficient and decreased high levels of RANKL/OPG. The present in vitro findings help to explain the possible beneficial role for bisphosphonates in the treatment of otosclerosis.
Author Contributions
Disclosures
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.