
Abstract
Background:
Amylovis-201 (CNEURO-201) shows strong neuroprotective activity in Alzheimer’s disease (AD) models due to its anti-Aβ aggregating properties and sigma-1 receptors (S1R) agonist activity. The drug is neuroprotective in transgenic rodent models of AD that could translate into disease-modifying effects by both protecting neuronal and glial viability and actively decreasing amyloid aggregation. The individual vulnerability to the neurodegenerative process in AD is highly dependent on the “cognitive reserve,” understood as the efficacy of brain plasticity throughout lifetime events.
Aims:
We aimed to determine whether Amylovis-201 efficacy in AD could be potentiated through stimulation of cognitive reserve. We combined Amylovis-201 administration with an experimental model of environmental enrichment (EE), using training on the Hamlet device. We analyzed the potentiating effects of the combination on brain plasticity and the development of resilience against memory deficits in a pharmacological AD model.
Methods:
We implemented a combined protocol with repeated per os (PO) administration of Amylovis-201 (0.1 and 1 mg/kg) and EE over 2 weeks (5 days/week), consisting of a 4-hour exploration of the Hamlet, a complex environment mimicking a small village with five functionalized houses.
Results:
The combination of EE and low dose (0.1 mg/kg PO) of Amylovis-201 had an additive effect on hippocampal neurogenesis. It also protected mice against scopolamine (0.5 mg/kg IP)-induced spatial working memory deficits in the Y-maze and, at 0.1 or 1 mg/kg PO, against Aβ25–35-induced memory deficits.
Conclusion:
As EE is known to increase S1R expression, the combination of EE and Amylovis-201 had major effects on brain plasticity and neuroprotection.
Keywords
Introduction
Alzheimer’s disease (AD) is a devastating neurodegenerative disease characterized by a generalized brain toxicity due to (i) pathological Amyloid-β (Aβ) peptide oligomerization and extracellular deposition and (2) pathological tau protein hyperphosphorylation and intracellular aggregation (Hung et al., 2016; Selkoe, 2013). Pathological hallmarks, therefore, include fibrillar amyloid aggregates evolving in senile plaques, intracellular neurofibrillary tangles destroying cellular cytoskeleton integrity, and a generalized neuroinflammation (Hung et al., 2016; Selkoe, 2013). Current therapeutic research aims at identifying small molecules or biological agents that, ideally, reverse the pathological process and therefore cure the disease. At least, an effective intervention must be able to promote effective neuroprotection by limiting the generation of pathological agents and preserving the patient’s brain physiology and cognitive status. Among the most promising pharmacological tracks, drugs acting as sigma-1 receptor (S1R) agonists showed potent neuroprotective activities in preclinical models in vitro and in vivo (Fisher et al., 2016; Marrazzo et al., 2005; Meunier et al., 2006). A compound showed positive results in a clinical phase III trial, Blarcamesine (Hampel et al., 2020; Macfarlane et al., 2025), and other drugs have a very promising profile (Fisher et al., 2016). S1R is a highly conserved transmembrane protein expressed in the endoplasmic reticulum (ER) membrane of numerous different cell types, including neurons and glial cells in the brain (Alonso et al., 2000). The protein acts mainly as a chaperone protein, by interacting with different client proteins and modulating numerous intracellular signaling pathways, including calcium exchange between the ER and mitochondria or cytosol (Hayashi et al., 2000; Hayashi & Su, 2007). S1R shows druggable features, and several small compounds have been characterized as agonists, positive modulators, or antagonists (Maurice, 2021; Vavers et al., 2019).
We have identified a new family of naphthalene derivatives forming stable complexes with Aβ peptides, using in silico evaluations, and inhibiting their aggregation in vitro. The lead compound, Amylovis-201 (CNEURO-201), was shown to decrease Aβ burden in the subiculum of the hippocampal formation of 3 × Tg AD mice and to improve their cognitive performance after chronic daily treatment for 8 weeks (Rivera-Marrero et al., 2020). Chronic oral treatment with 0.1 mg/kg of Amylovis-201 alleviated learning and memory deficits, reduced cholinergic dysfunction, glial activation, and oxidative stress, and decreased the Aβ burden in 12–14 months old 3 × Tg-AD mice (Martínez-Orozco et al., 2025). Amylovis-201, therefore, appears as a very active neuroprotectant compound in AD models. Due to structural similarities between the chemical scaffold of Amylovis-201 and the pharmacophore requirements for an effective binding to S1R, we previously examined the drug interaction with S1R and, based on in silico, in vitro, and in vivo experiments, showed that Amylovis-201 is a potent S1R agonist (García-Pupo et al., 2024). Amylovis-201 acts, therefore, as a potent anti-amnesic and neuroprotective drug in animal models, with a unique dual activity as Aβ anti-aggregating agent and neuroprotective S1R agonist. The drug is a promising neuroprotectant for AD by promoting Aβ clearance and cellular protection.
Environmental enrichment (EE) is known to increase the degree of brain plasticity, notably by stimulating dentate granule cells neurogenesis and glial proliferation, reducing spontaneous apoptotic cells death in key brain structures like the hippocampus, inducing expression of glial-derived neurotrophic factor and brain-derived neurotrophic factor (BDNF), and increasing phosphorylation of key transcription factors like cyclic-adenosine monophosphate (AMP) response element-binding protein (CREB; Young et al., 1999). As a consequence, in neurodegenerative diseases like AD, EE improved cognitive performance and reduced amyloid load and deposition in rodent models, such as PPswe mice (Arendash et al., 2004). EE also induced selective upregulation in levels of specific gene transcripts associated with learning and memory, vasculogenesis, neurogenesis, and cell survival pathways in the brain of transgenic mice (Lazarov et al., 2005). Moreover, EE has been reported to increase S1R expression, for instance, in rats housed in a stimulated environment for 2 weeks after permanent middle cerebral artery occlusion (Ruscher et al., 2011) or in mice trained in the Hamlet test (Crouzier et al., 2020). Repeated treatment with the S1R antagonist NE-100 blocked the behavioral or biochemical consequences of EE in trained mice, such as augmented histone acetylation in the hippocampus and cortex or increased cell proliferation and maturation in the hippocampus (Crouzier et al., 2020). Whether EE and S1R activity could share a common mechanistic mode of action and synergistically affect cellular physiology and pharmacological pathways remains to be explored.
In the present study, we addressed the potentialities of a combination between EE and a treatment with Amylovis-201 on brain plasticity, that is, hippocampal neurogenesis or vulnerability to scopolamine-induced amnesia, and as a protective strategy in the AD context. EE was induced using a 2-week training period in the Hamlet, as previously described (Crouzier et al., 2018, 2020), and combined with a repeated per os (PO) treatment with Amylovis-201, at a dose previously used in 3 × Tg AD mice (Martínez-Orozco et al., 2025). The combination effect was analyzed, first, on hippocampal neurogenesis and behavioral stimulation (scopolamine-induced memory deficits) and, second, on Aβ25–35-induced memory deficits. Results show significant additive effects of EE and Amylovis-201 combination, presumably mediated by upregulation and enhanced activation of the S1R.
Experimental procedures
Animals
C57BL/6J mice (256 males, 54 females; Janvier Labs, Saint-Berthevin, France), aged 7–9 weeks, were used. All Hamlet-based training protocols were conducted exclusively with male subjects. Female mice were housed within the interacting house module of the Hamlet apparatus to provide social stimulation. Mice were housed in plastic cages in groups of eight individuals. They had free access to food and water except when specified, and they were kept in a regulated environment (22 ± 1°C, 40%–60% humidity) under a 12-hour light/dark cycle (light on at 07:00 am). Experiments were carried out between 09:00 am and 06:00 pm, in an experimental room within the animal facility. All animal procedures were conducted in strict adherence to the European Union directive 2010/63 and the ARRIVE guidelines (Kilkenny et al., 2010) and were authorized by the French national ethics committee (APAFIS#2564-201511 0315437060).
Drugs and administration procedures
Methyl (2-{[4-(1-naphthylamino)-4-oxobutanoyl]amino}ethyl)dithiocarbamate (Amylovis-201; CAS 1624322-36-6) was synthesized as previously described (Rivera-Marrero et al., 2020). (−)-Scopolamine hydrobromide, 5-bromo-2′-desoxyuridine (BrdU), and 4-methoxy-3-(2-phenylethoxy)-N,N-dipropylbenzeneethanamine hydrochloride (NE-100) were from Merk (Saint-Quentin-Fallavier, France). All drugs were solubilized in physiological saline (vehicle solution) and administered intraperitoneally (IP) or PO by gavage, in a volume of 5 µL/g body weight, at specified times before the behavioral tests.
Amyloid-β[25–35] fragment (Aβ25–35) and the scrambled control peptide (Sc.Aβ) were from Eurogentec (Angers, France). They were dissolved in sterile distilled water at 3 mg/mL and stored at −20°C until use. They were incubated at 37°C for 4 days allowing Aβ25–35, but not Sc.Aβ (Maurice et al., 1996; Zussy et al., 2013), to oligomerize as small prefibrillar aggregates (Zussy et al., 2013), and injected intracerebroventricularly (ICV) at the dose of 9 nmol/3 µL using the Haley and McCormick (1957) method with the approximate stereotaxic coordinates AP: Bregma −0.5, L: ±1.0, H: −2.5 mm according to Paxinos and Franklin (2004), under isoflurane anesthesia as previously described as previously described (Maurice et al., 1996).
The Hamlet test
The Hamlet apparatus, designed by us but manufactured by Viewpoint (Lissieu, France), is shown in Figure 1(a); (Crouzier et al., 2018, 2020). The device is 1.6 m in diameter and placed in a room with standard laboratory furniture and posters on the walls. The room was uniformly illuminated (200 Lux). The Hamlet has an agora at its center and streets expanding in a star shape toward five functionalized roofed compartments, called houses (Figure 1(b)). Walls and walkways were made of white or black infrared (IR) light-transparent Plexiglas. The agora served as a gathering area and a starting point for training and test trials. The functionalized houses contained a food dispenser (physiological function encoded: Eat), two water dispensers (Drink), a novomaze® shelter (Viewpoint) (Hide), a running wheel (Run), or a compartment for a stranger mouse (Interact). A wire mesh screen separated the stranger mouse, whereas the other houses could be entered freely. The stranger mouse was a female C57BL/6J mouse. Movements were recorded by an IR light camera set above the Hamlet and analyzed using a Videotrack® software (Viewpoint, Lyon, France). Training was performed according to the published protocol (Crouzier and Maurice, 2018). In brief, animals from the same housing cage, that is, groups of 8 individuals knowing each other, were placed in the Hamlet for 4 hours per day during a 2-week training period (Figure 1(c)). During training, the number of entries and duration of presence in each house were videotracked and analyzed. Global activity was determined by dividing the time spent in each of the five houses by the number of mice in the Hamlet for each hour during the 4 hours of a training session. Topographical memory was evaluated by testing twice animals, 3 and 4 days after the last training session, individually, after water-deprivation (WD) or not (NWD; Crouzier and Maurice, 2018). In the WD condition, the drinking bottle was removed from the housing cage 15 hours before testing. Each mouse, in the WD or NWD condition, was placed in the agora and free to explore the Hamlet during a 10-min session. The exploratory behavior was videotracked and analyzed in terms of latency to reach the goal and other houses and the number of errors (entries into a street not directed to the goal house). Trained (T) mice were compared between NWD and WD conditions, and with non-trained (NT) animals, naive to the Hamlet, and also tested in WD and NWD conditions.
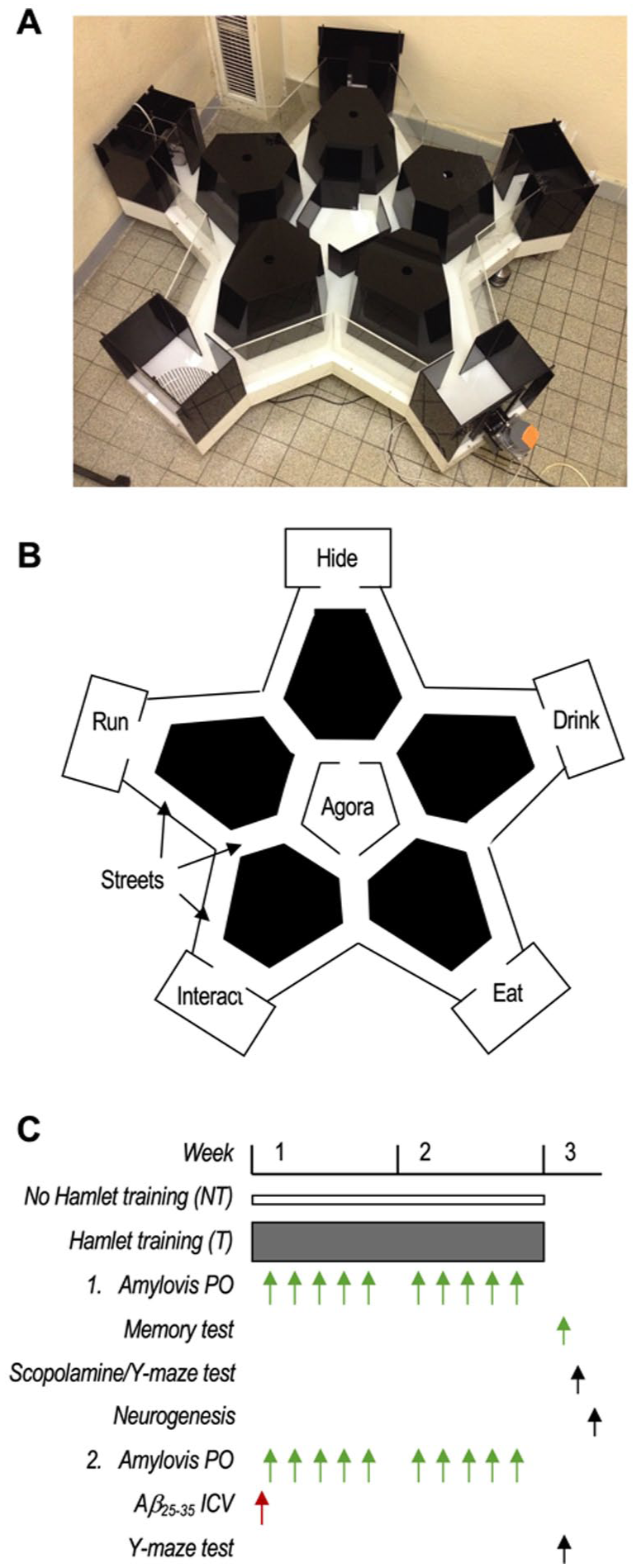
The Hamlet test and protocol were used. (a) The device in the experimental room. (b) The map of Hamlet shows the five functionalized houses, the central agora, and streets extending from it. (c) Protocol 1: animals were trained in the Hamlet (T) or not (NT) and received Amylovis-201 daily (0.1 or 1 mg/kg PO). The memory test in water-deprived (WD) or non-water-deprived (NWD) conditions was performed 72 hours after the last training session. Animals were then tested for scopolamine (0.5 mg/kg IP)-induced alternation impairment in the Y-maze, and hippocampal neurogenesis was analyzed. Protocol 2: Mice received Aβ25–35 (9 nmol ICV) and were then trained or not in the Hamlet and/or received Amylovis-201 daily (0.1 or 1 mg/kg PO). They were then tested for spontaneous alternation in the Y-maze.
Experimental groups
Two independent experiments were performed (Figure 1(c)). First, animals (3 batches of eight mice, n = 24 max per condition) were treated with Amylovis-201 (0.1 mg/kg PO) or vehicle solution (Veh, V) 5 days a week for 2 weeks and trained (T) or not (NT) in the Hamlet. This protocol was designed according to the observation that the S1R antagonist NE-100 blocked Hamlet training effects on brain plasticity using a similar administration protocol (Crouzier et al., 2020). Weight gain, exploration of the Hamlet, and topographic memory were analyzed before testing scopolamine-induced amnesia (impairment of spontaneous alternation in the Y-maze measured 20 minutes after scopolamine injection) or hippocampal neurogenesis in the mice. In the second experiment, animals (3 batches of eight mice, n = 8 or 16 max per condition) received an ICV administration of Aβ25–35 or Sc.Aβ peptide (9 nmol) and were then treated with Amylovis-201 (0.1 or 1 mg/kg PO) or vehicle solution 5 days a week for 2 weeks and trained (T) or not (NT) in the Hamlet. Weight gain and exploration of the Hamlet were analyzed, and the spontaneous alternation ability was measured and analyzed as an impact of EE + Amylovis-201 treatment on Aβ25–35-induced impairment of alternation. Animals were sacrificed after the behavioral observations, and for some of them, their brains were dissected out and post-fixed for immunofluorescence analyses (see section “Tissue preparation and brain slicing”).
Spontaneous alternation in the Y-maze
Spontaneous alternation performance was measured in the Y-maze as an index of spatial working memory (García-Pupo et al., 2024; Lahmy et al., 2013; Maurice et al., 1996; Villard et al., 2011). Animals were allowed to explore the maze for 8 minutes and the percentage of alternation was calculated as (actual alternations/number of arm entries − 2) × 100. Scopolamine, 0.5 mg/kg, was administered IP, 20 minutes before the session.
Tissue preparation and brain slicing
BrdU was administered at 150 mg/kg IP twice at 4-hour intervals, and mice were sacrificed after 24 hours. Mice were anesthetized with ketamine (75 mg/kg IP) + xylazine (25 mg/kg IP), transcardially perfused with 40 mL phosphate buffer saline followed by 40 mL AntigenFix (F/P001X, Microm Microtech, Brignais, France). The brain dissected out and stored in Antigenfix at 4°C was finally transferred to AntigenFix/sucrose 10%, 48 hours after sacrifice, and stored at 4°C until use. They were cut in coronal sections (25 µm thickness) using a freezing microtome (Microm HM 450, Thermo Fisher, Waltham, MA, USA), collected in a 24-well plate, and stored in cryoprotectant at −20°C. Serial sections were selected to include the hippocampal formation and processed onto slides, with each slide containing five to eight sections from one mouse.
Neurogenesis analysis
The brain sections were blocked with a solution containing 0.01 M PBS, 0.1% Triton X-100, and 10% normal goat serum solution for 30 minutes. They were subsequently incubated at 4°C overnight with primary antibodies diluted in PBS: rat anti-BrdU (dilution 1:250, ref. ab6326, Abcam, Boston, MA, USA) or rabbit anti-doublecortin (DCX, dilution 1:500, ref. 4604S, Cell Signaling Technology). After several washes, sections were incubated with biotinylated goat anti-rat (dilution 1:500, ref. ab207997, Abcam) or anti-rabbit (dilution 1:200, ref. BA-1000, Vector Laboratories) secondary antibody and finally in avidin–biotin–peroxidase complex (PK-4000) and detected with diaminobenzidine. After dehydration through successive baths of alcohol, all sections were counterstained with DAPI and cover-slipped with mounting medium (Mountex). The BrdU immunoreactive cells in the granule cell layer and sub-granular zone of the dorsal dentate gyrus (DG; AP: Bregma −1.8 to −2.5 mm) were manually counted using an optical microscope (DM 2500, Leica) under white light at a magnification of ×40. Each DCX neuron having a single dendrite was measured within 40 μm of the soma. An image of hippocampus cells was projected on a computer, with a ×40 objective, and a line segment was drawn along the middle of the cells. DCX dendrites crossing the segment were counted. The number of BrdU or DCX-positive cells was determined by combining cell counts per section for the whole DG and multiplying by the number of brain sections, including the hippocampus. Bilateral countings were averaged, and 5–8 slides were counted per animal (n = 5–8, N = 5). All data were expressed as a percentage of the Veh-treated NT group.
Statistical analyses
Data were expressed as mean ± SEM. They were analyzed using the Prism v10.6 statistical software (GraphPad, CA, USA), by one-way, two-way, or three-way analyses of variance (ANOVA) for multiple comparisons (F values), with treatment or training days and ICV or PO treatments as independent factors. Repeated measure was considered for treatment or training days, as specified. Post hoc comparisons were done using Dunnett’s test. A one-column t-test was also used when comparing to a reference level. The level of statistical significance considered was p < 0.05. Statistical analyses were performed using the Prism v10.0 (GraphPad software, Boston, MA, USA) software All statistical values are detailed in Supplemental Table 1.
Results
Combination of EE and Amylovis-201 treatment
We first examined the effect of combining EE, achieved through group training of mice in the Hamlet device, with low-dose Amylovis-201 treatment, known for its cognition-enhancing properties (García-Pupo et al., 2024; Mercerón-Martínez et al., 2024; Martínez-Orozco et al., 2025; Rivera-Marrero et al., 2020). Mice were trained in the Hamlet 4 hours daily for 2 weeks, or not, and received daily oral treatment with Amylovis-201 (0.1 mg/kg; Martínez-Orozco et al., 2025) or vehicle solution (Figure 1(c)). Mouse body weight was monitored regularly and, compared with NT groups, Hamlet exploration resulted in significantly lower weight gain, unaffected by pharmacological treatment (Figure 2(a) and (b)). Hamlet exploration was first analyzed as a global activity, that is, the total number of entries into the houses during the 4-hour period, at the beginning (day 1), mid-term (day 6), and end (day 10) of training (Figure 2(c)). Amylovis-201 treatment significantly decreased global activity on days 1 and 10 compared with vehicle-treated animals. However, topographic analysis of Hamlet exploration showed that both vehicle- and Amylovis-treated groups exhibited a house preference order of Run > Hide = Drink = Eat > Interact when analyzed by number of entries (Figure 2(d)), and Run = Interact = Hide = Eat > Drink when analyzed by duration of presence (Figure 2(e)). Furthermore, vehicle-treated animals showed sustained exploration through day 1, whereas Amylovis-201-treated animals displayed a peak in exploration for both parameters on day 6 (Figure 2(d) and (e)). These observations indicated that Amylovis-201 treatment mildly affected Hamlet exploration during the training phase.
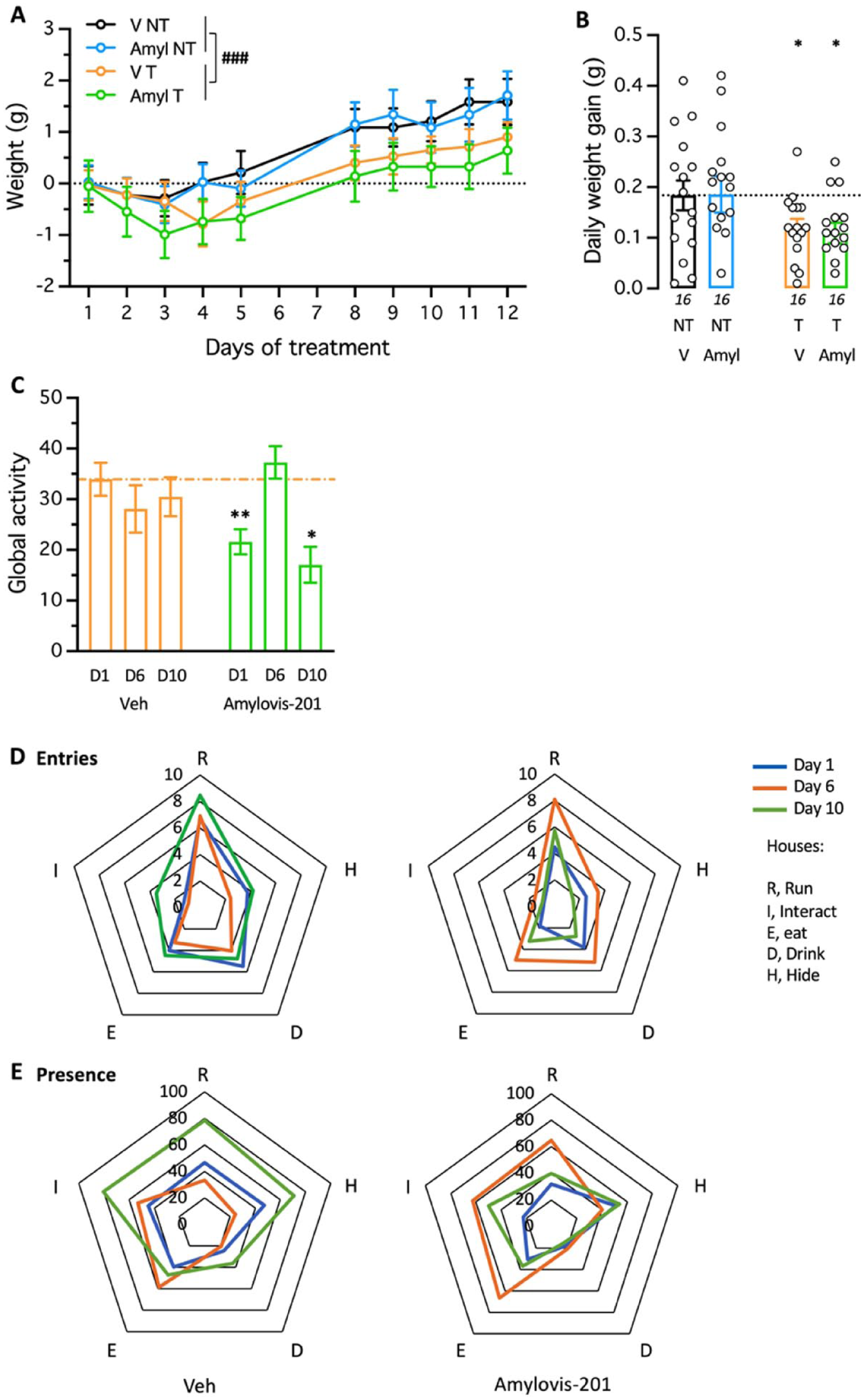
Effect of Amylovis-201 during training in the Hamlet. (a) Daily body weight gain of the mice during the 2-week training in the Hamlet and/or PO treatment with Amylovis-201 (0.1 mg/kg). Note that Hamlet training significantly limited weight gain over the training period, similarly in V- or Amylovis-201-treated groups; ###p < 0.001. (b) Averaged daily weight gains over the 2 weeks. *p < 0.05 versus V-treated NT group, Dunnett’s test. (c) Global activity measured as presence in all houses during the 4-hour training duration on days 1, 6, and 10 of training. *p < 0.05, **p < 0.01 versus the same-day Veh-treated group, Dunnett’s test. (d) Entries and (e) durations of presence in each house, expressed per hour per mouse, shown as radar graphs for days 1, 6, and 10 of training for the Veh- and Amylovis-201-treated groups.
The topographic memory was analyzed by testing each mouse individually for its ability to locate the Drink house, 72 hours after the last training session. The animals were tested in water-deprived (WD) conditions and then in non-water-deprived (NWD) conditions. NT animals, either Veh- or Amylovis-201-treated, showed a mean latency to find the Drink house of 179 ± 20 seconds (median 127 (46–247) seconds) not significantly different between the NWD and WD conditions (Figure 3(a)), thus with a NWD/WD ratio close to 1.0 (Figure 2(b)). In terms of errors, NT animals showed a mean number of errors to find the Drink house of 31 ± 3 (median 23 (13–41)), not significantly different between the NWD and WD conditions (Figure 3(c)), thus with a NWD/WD ratio close to 1.0 (Figure 2(d)). Trained Veh-treated animals showed a significant decrease in latency (Figure 3(a)) and a close-to-significance decrease in errors (Figure 3(c)) to find the Drink house in WD as compared to the NWD condition. This resulted in NWD/WD ratios for both parameters significantly higher than 1.0 (Figure 3(b) and (d)). The Amylovis-201 treatment did not affect the ability of the mice to locate the Drink house, as both latencies and errors were significantly lower in WD as compared to NWD condition (Figure 3(a) and (c)). Accordingly, NWD/WD ratios were also significantly higher than 1.0 (Figure 3(b) and (d)). These data suggested that the Amylovis-201 treatment did not markedly impact topographic memory measured following Hamlet training. However, it is noteworthy that errors in the WD group treated with Amylovis were lower than in the vehicle-treated group.
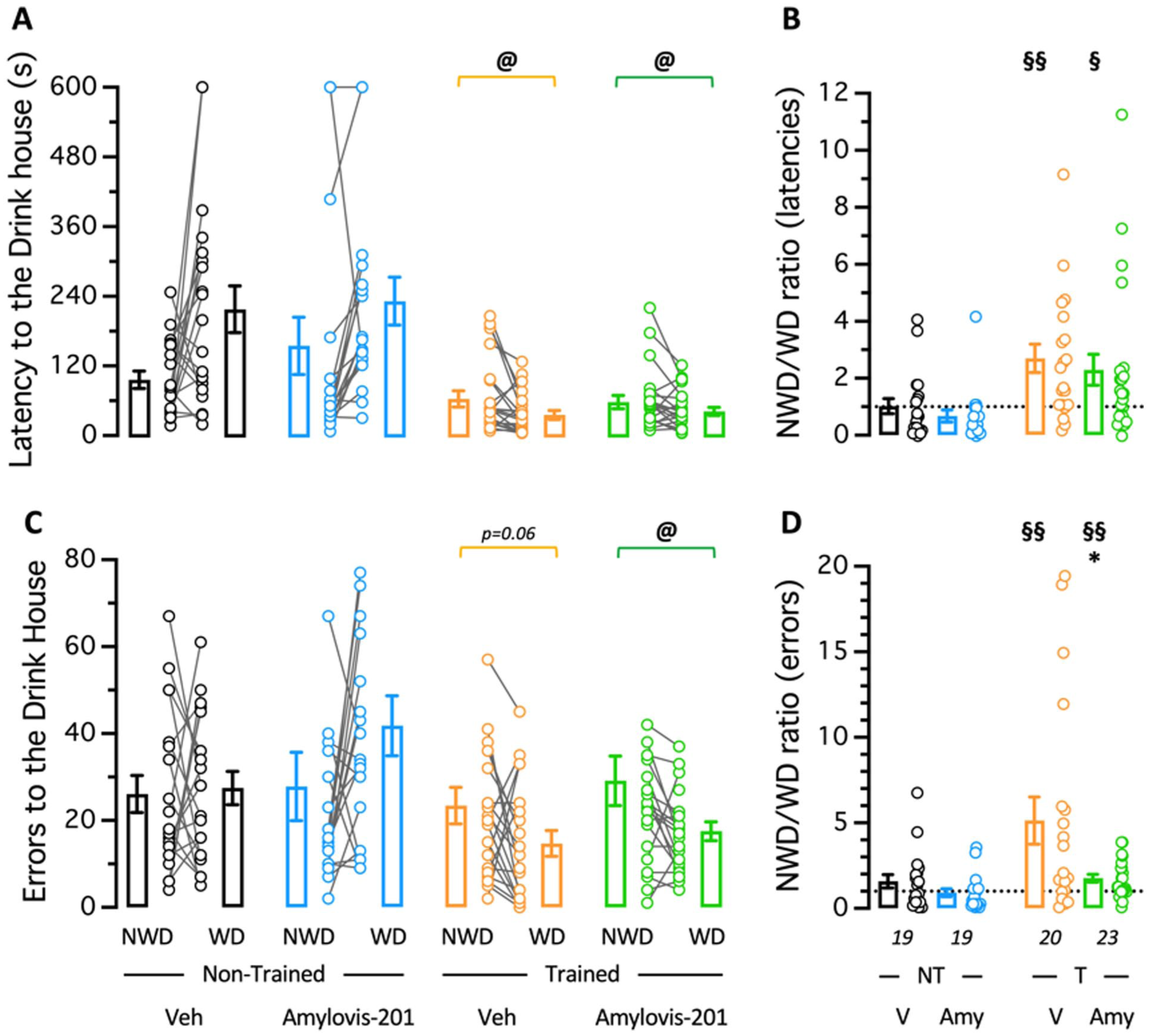
Topographic memory in Veh- or Amylovis-201-treated mice trained in the Hamlet. Three days after the last training session, trained (T) or non-trained (NT) mice were tested during 10 minutes for their ability to reach the Drink house. Animals were tested in water-deprived (WD) or, after 24 hours, in non-water-deprived (NWD) conditions. Data show the latency to reach the Drink house (a) and the calculated ratio between the NWD and WD conditions (b), and the number of errors (b) and the calculated ratio between the NWD and WD conditions (d). In (a and c), data show the mean and SEM as a bar graph and the individual data as NWD-WD pairs. In (b and d), value “1.0” corresponding to the same value in NWD and WD conditions is shown by a dotted line. @p < 0.05, paired t-test in (a and c); *p < 0.05 versus V-treated group; unpaired t-test; §p < 0.05, §§p < 0.01 versus value “1.0,” one-sample t-test in (b and d).
To determine whether the combination of EE and Amylovis-201 could have an additive effect on brain plasticity, we first assessed the scopolamine-induced amnesia. Crouzier et al. (2018) reported that EE confers resistance to the amnesic effect of scopolamine. Indeed, the muscarinic acetylcholine receptor antagonist significantly affected spontaneous alternation in the Y-maze or the passive avoidance response at the high dose of 5 mg/kg IP in trained animals, versus 0.5 mg/kg IP in NT mice (Crouzier et al., 2018). We therefore tested the alternation performances of mice treated, 20 minutes before the session, with scopolamine 0.5 mg/kg IP. Veh-treated NT mice treated with scopolamine showed an alternation percentage close to 50% (Figure 4(a)), indicating a major alternation deficit as compared to the 65%–70% performance observed in control animals, and a number of entries necessary to perform the first alternation (NEFA) of 4.8 ± 0.4 (Figure 4(b)), significantly higher than only three arm entries in control animals. The Amylovis-201-treatment in NT animals attenuated, but non-significantly, the deficit in spontaneous alternation (Figure 4(a)) and significantly decreased the NEFA (Figure 4(b)). Training in the Hamlet also increased, but significantly, the alternation performance (Figure 4(a)) but more moderately improved the NEFA (Figure 4(b)). However, the EE + Amylovis-201 combination resulted in a higher, very significant increase in alternation that reached control values (64.5%; Figure 4(a)) and decreased NEFA to control value (3.4 + 0.2; Figure 4(b)). The pre-administration of the reference S1R antagonist NE-100, 3 mg/kg IP 20 minutes before the session, completely blocked the EE or EE+Amylovis-201 effects (Figure 4(a) and (b)). Analysis of the locomotion during the Y-maze exploration showed that training in Hamlet resulted in an increase in mobility of the animals which was not affected by the Amylovis-201 treatment (Figure 4(c)). Interestingly, the NE-100 treatment also completely blocked this increase in mobility, suggesting an S1R-mediated effect (Figure 4(c)).
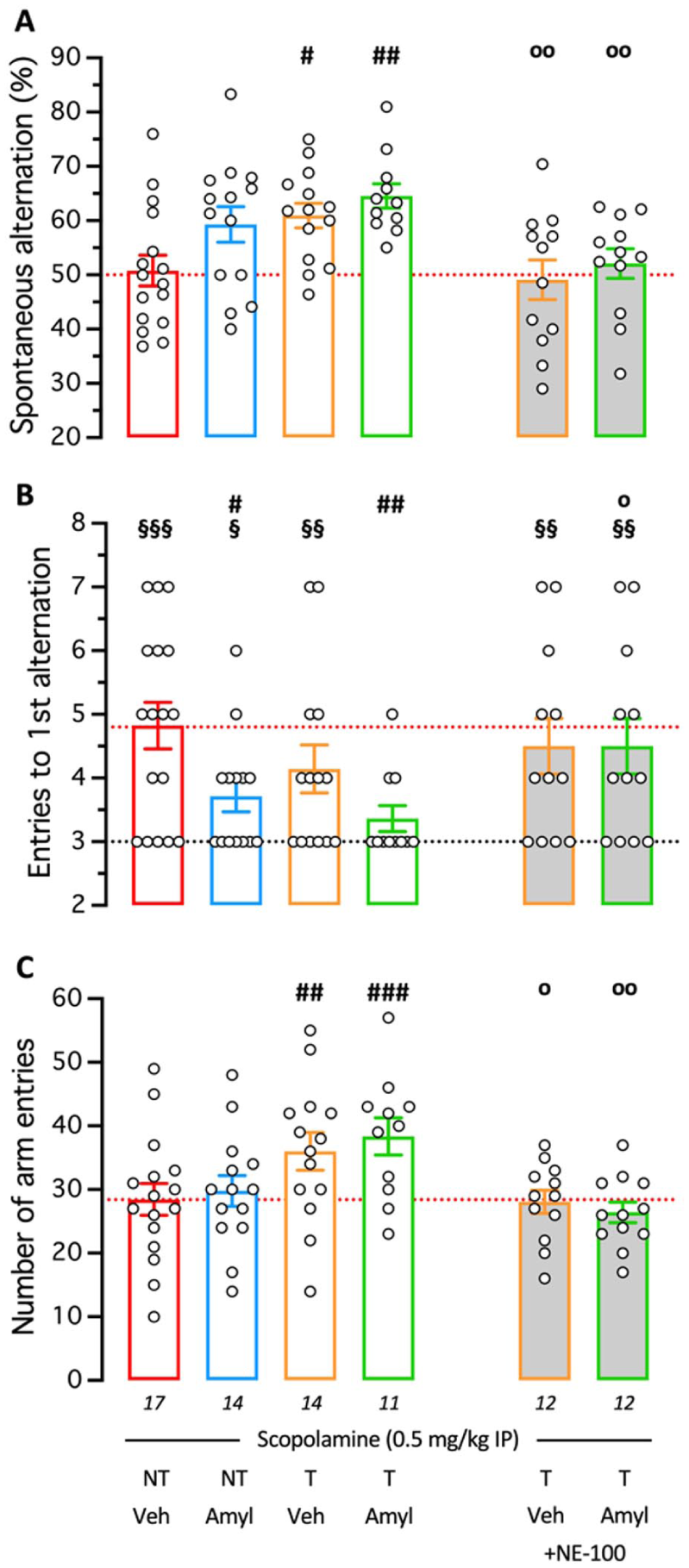
Combined effect of Hamlet training and PO treatment with Amylovis-201 on prevention of scopolamine-induced spontaneous alternation deficits. Mice, trained (T) or not (NT) in the Hamlet and administered with Veh or Amylovis-201 (0.1 mg/kg PO), were treated with scopolamine, 0.5 mg/kg IP 20 min before being tested in the Y-maze: (a) spontaneous alternation, (b) number of entries to the first alternation (NEFA), and (c) total number of arm entries. Some animals received the S1R antagonist NE-100 (3 mg/kg IP) 30 minutes before the Y-maze session. In (a), the red dotted line, 50% alternation level, corresponds to a total impairment in working memory. In (b), the black dotted line, level “3,” corresponds to the minimal arm entries necessary to perform a correct alternation, and the red dotted line is the level in Scopolamine-treated animals. In (c), the red dotted line corresponds to the scopolamine-treated group data. #p < 0.05, ##p < 0.01, ###p < 0.001 versus V-treated NT group treated with scopolamine alone; op < 0.05, oop < 0.01 versus the same treatment group without NE-100; Dunnett’s test. §p < 0.05, §§p < 0.01, §§§p < 0.001 versus value “3.0”; one-sample t-test.
As a major consequence of EE- and S1R-mediated brain plasticity (Crouzier et al., 2020; Kempermann et al., 1997; Li et al., 2010), we analyzed hippocampal neurogenesis across experimental groups. Neither Amylovis-201 treatment alone nor Hamlet training (EE) alone affected the number of newborn cells, whereas the EE + Amylovis-201 combination significantly increased BrdU-positive cell counts (Figure 5(a) and (b)). Cell maturation was assessed by DCX immunofluorescence. Although Amylovis-201 alone did not alter DCX-positive cell numbers, both EE alone and the EE + Amylovis-201 combination significantly enhanced cell maturation (Figure 5(c) and (d)).
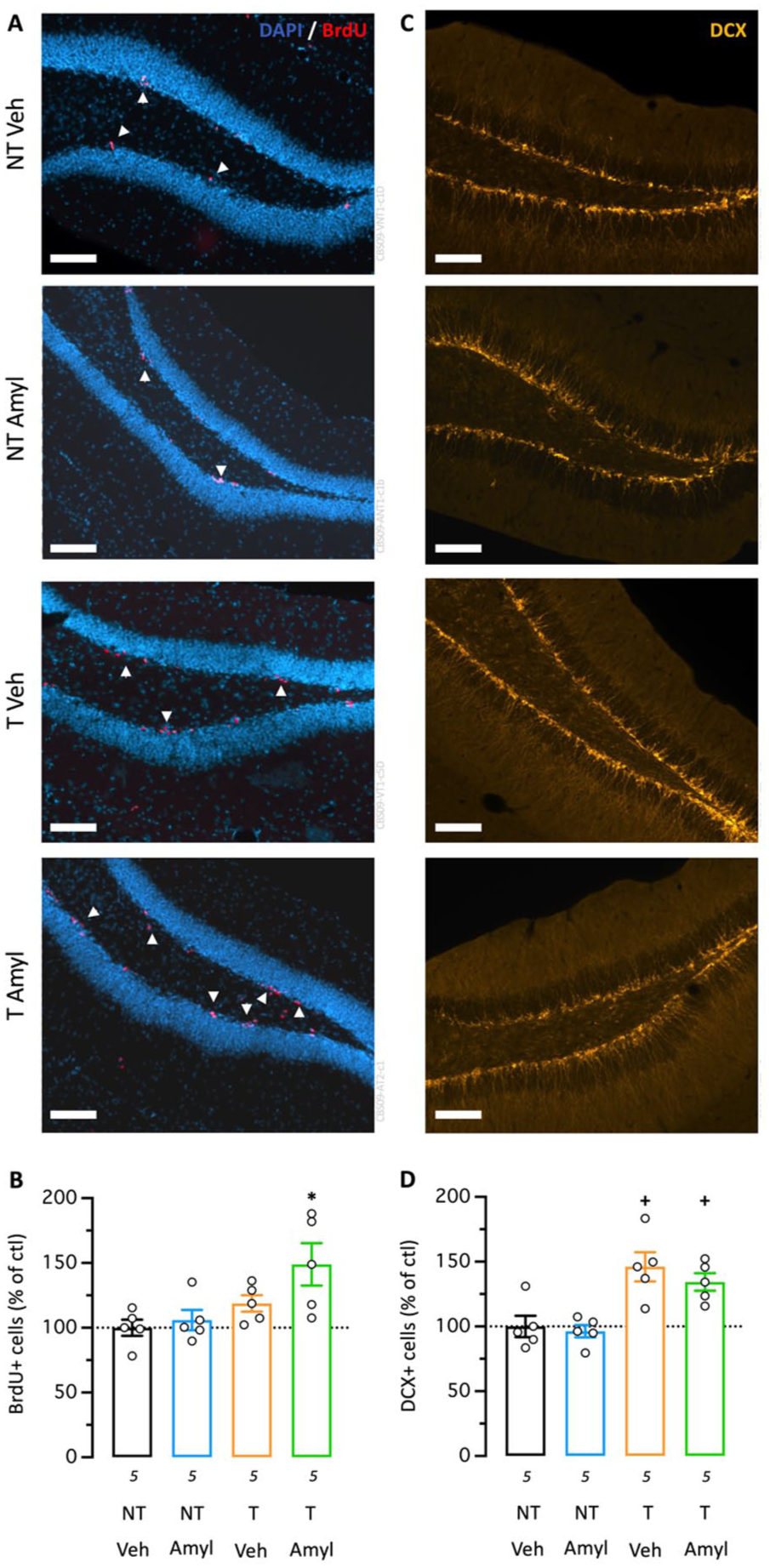
Combined effect of Hamlet training and PO treatment with Amylovis-201 on hippocampal neurogenesis. (a) Typical micrographs and (b) quantification of BrdU immunofluorescence in the mouse hippocampal dentate gyrus, labeling cell proliferation. Arrowheads point to BrdU-positive cells. (c) Typical micrographs and (d) quantification of doublecortin (DCX), labeling cell maturation. Scale bars = 50 µm. *p < 0.05 versus Veh-treated group; +p < 0.05 versus NT group; Dunnett’s test.
These different observations suggested that EE and Amylovis-201 treatment exert additive effects on brain plasticity.
Effect of EE and Amylovis-201 combination on Aβ25–35-induced learning impairments
In this set of experiments, we analyzed the impact of EE and Amylovis-201 treatment, administered at two doses: low (0.1 mg/kg) and high (1 mg/kg) via PO for 2 weeks on Aβ25–35-induced spontaneous alternation deficits. Mice were injected with Aβ25–35 or Sc.Aβ on day 1 and subsequently, the groups treated with vehicle or Amylovis-201 at two independent doses initiated training in the Hamlet apparatus, as presented in Figure 1(c). Body weight was monitored regularly (Figure 6(a)) and a few significant differences as compared to the NT Sc.Aβ/Veh-treated group were noted: the Amylovis-201 high-dose treatment slightly but significantly lowered weight gain in NT animals, and the Aβ25–35 treatment slightly but also significantly increased weight gain in NT animals (Figure 6(b)). These differences disappeared in T animals (Figure 6(b)). Analysis of the global activity confirmed the lowering effect of Amylovis-201 at the low dose (0.1 mg/kg), both in Sc.Aβ- and Aβ25–35-treated animals, with significant effect on days 6 and 10 for the latter (Figure 6(c)). It must be noted that the Amylovis-201 high-dose (1 mg/kg) treatment tended to increase global activity in both the Sc.Aβ- and Aβ25–35-treated groups. Analysis of the house exploration, in terms of entries and presence, confirmed that, without changing the pattern of exploration preference among the houses, the Amylovis-201 treatment at low dose diminished the number of entries in all houses except the Run house for both Sc.Aβ- and Aβ25–35-treated animals and that the Aβ25–35 and Amylovis-201 high-dose treatments tended to increase entries in the Hide, Drink, and Eat houses (Figure 6(d)). The patterns of presence slightly changed with the Aβ25–35 treatment (less presence in the Hide house and more in the Interact house) but failed to show a clear impact of the Amylovis-201 treatments (Figure 6(e)).
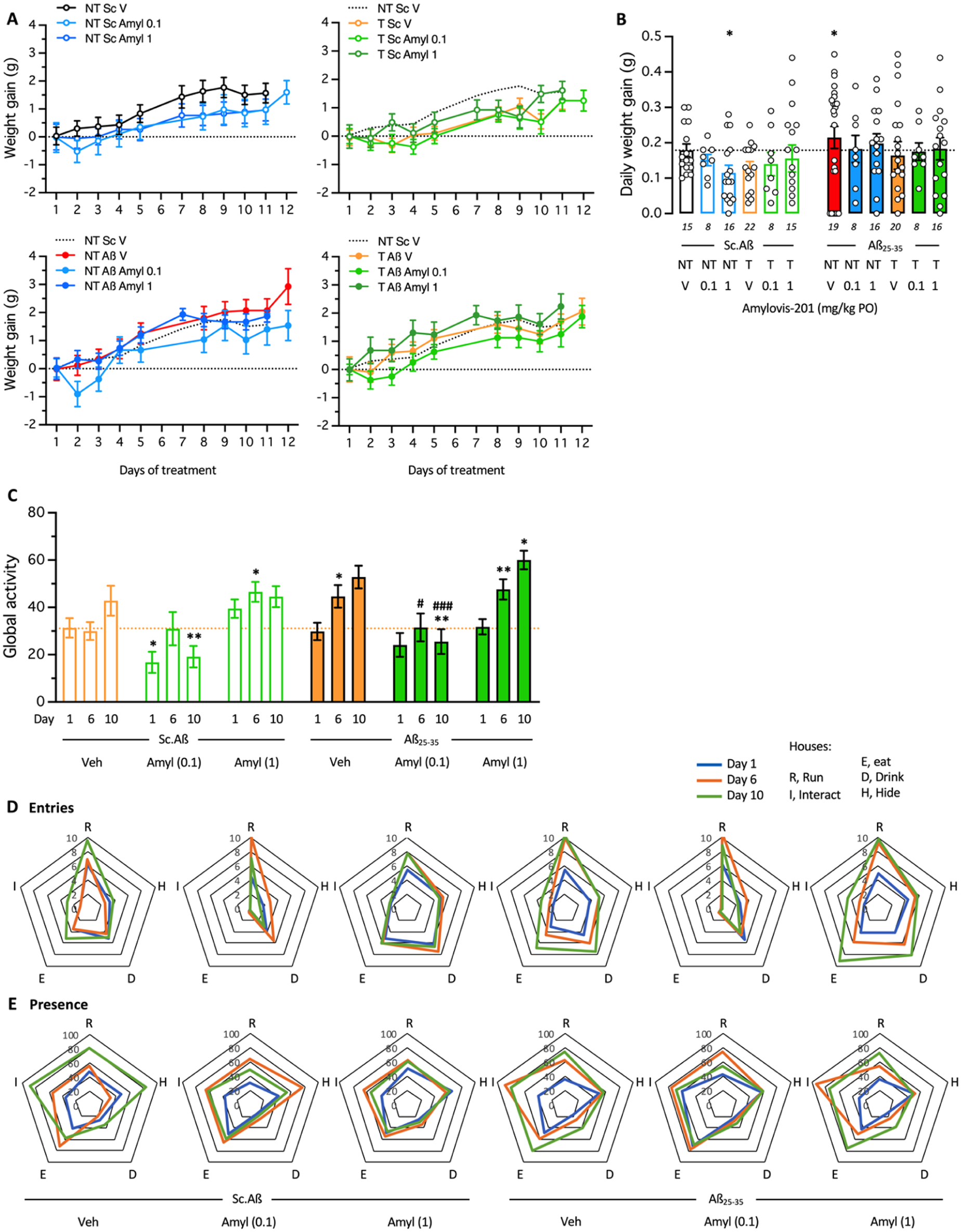
Effect of Aβ25–35 and Amylovis-201 treatments during training in the Hamlet. (a) Daily body weight gain of the mice during the 2-week training in the Hamlet and/or PO treatment with Amylovis-201 (0.1 mg/kg) and /or ICV treatment with Aβ25–35 or Sc.Aβ (9 nmol). (b) Averaged daily weight gains over the 2 weeks. *p < 0.05 versus Sc.Aβ/V-treated NT group, Dunnett’s test. (c) Global activity measured as presence in all houses during the 4-hour training duration on days 1, 6, and 10 of training. *p < 0.05, **p < 0.01 versus the same-day Sc.Aβ/Veh-treated group, #p < 0.05, ###p < 0.001 versus the same-day Aβ25–35/Veh-treated group; Dunnett’s test. (d) Entries and (e) durations of presence in each house, expressed per hour per mouse, shown as radar graphs for days 6 and 10 of training for each group.
After training, the mice were tested in the Y-maze (Figure 7). The training and/or Amylovis-201 treatments slightly but non-significantly affected the spontaneous alternation performances of the mice (Figure 7(a)). Training significantly increased exploration measured using the number of arm entries (Figure 7(b)), as previously observed. The Aβ25–35-treatment resulted in a significant alteration of the alternation performance that was attenuated but non-significantly by the Amylovis-201 treatments. The low-dose treatment appeared more effective (Figure 7(a)). Training failed to affect the Aβ25–35-induced deficit, but the combinations between EE and Amylovis-201 resulted in an extremely significant blockade of Aβ25-35-induced deficit at the low drug dose and in a significant attenuation at the high dose (Figure 7(a)). Interestingly, a pre-treatment with NE-100 before the Y-maze test session completely prevented the attenuating effect of the EE+Amylovis-201 combinations. In terms of mobility, the hyperlocomotion effect due to EE was noted in Aβ25–35-treated groups, this effect was also prevented by the NE-100 treatment (Figure 7(b)).
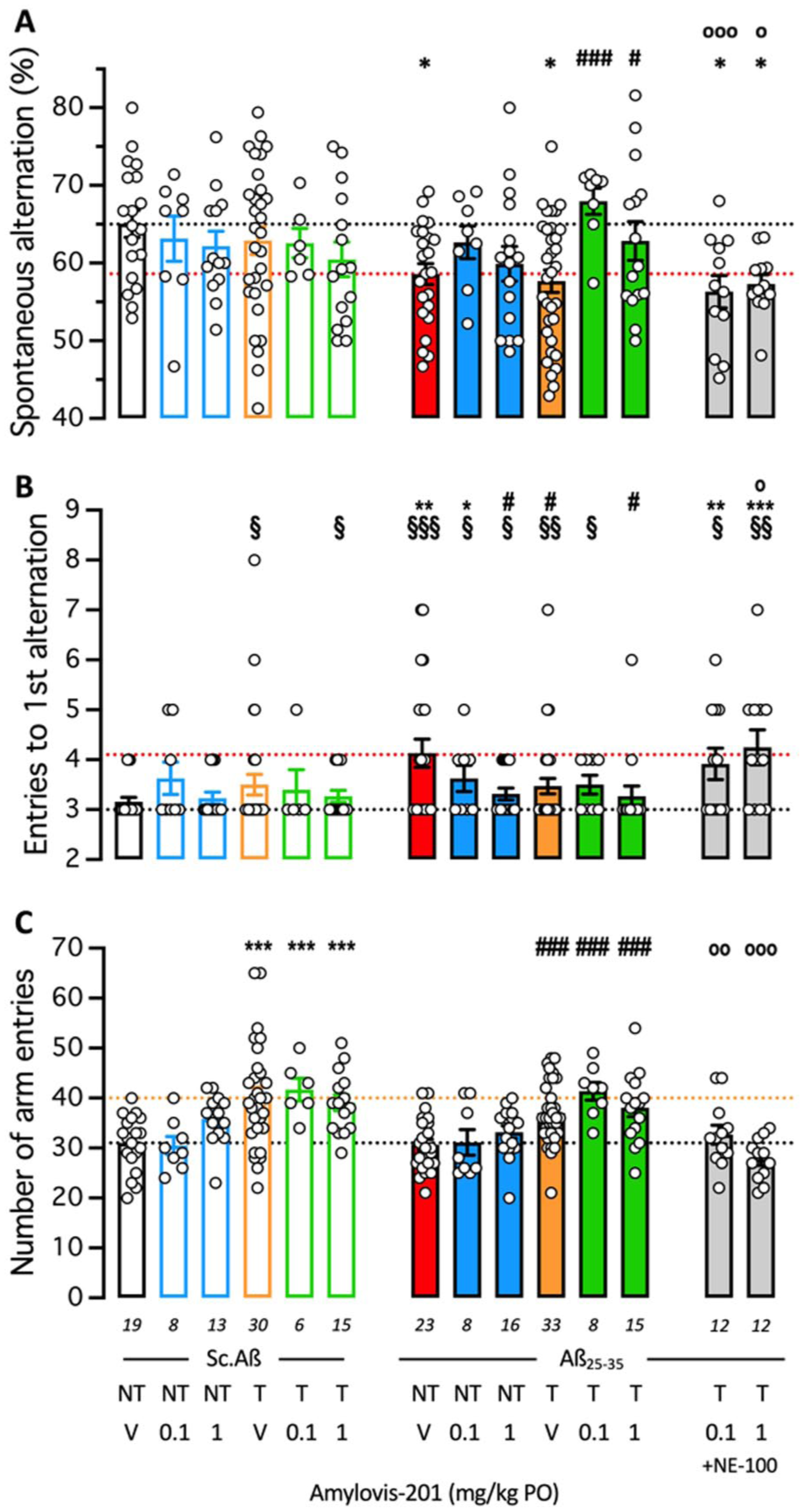
Combined effect of Hamlet training and PO treatment with Amylovis-201 on Aβ25–35-induced memory deficit: (a) spontaneous alternation, (b) number of entries to the first alternation (NEFA), and (c) total number of arm entries in the Y-maze test. Some animals received the S1R antagonist NE-100 (3 mg/kg IP) 30 minutes before the Y-maze session. *p < 0.05, **p < 0.01, ***p < 0.001 versus Sc.Aβ/V-treated NT group; #p < 0.05, ###p < 0.001 versus Aβ25–35/V-treated NT group; °p < 0.05, °°p < 0.01, °°°p < 0.001 versus same treatment group without NE-100; Dunnett’s test. §p < 0.05, §§p < 0.01, §§§p < 0.001 versus value “3.0”; one-sample t-test.
These observations showed that EE and Amylovis-201 induced an additive protective effect against Aβ25–35-induced impairment of spontaneous alternation, a measure of spatial working memory, and that the low-dose treatment with Amylovis-201 was more potent than the high-dose treatment tested here.
Discussion
In the present study, we addressed the potentialities of combining the brain plasticity gain induced by EE and a chronic treatment with Amylovis-201. This drug is a multi-target ligand that particularly alleviates amyloid load in murine models of AD (Martínez-Orozco et al., 2025; Rivera-Marrero et al., 2020) and activated S1R to induce a potent neuroprotection in rodent models (García-Pupo et al., 2024). EE was achieved by a 2-week exposure of groups of mice living together in the same cage in the Hamlet device (Crouzier et al., 2018). The first result was to observe that the Amylovis-201-treatment had a marginal effect on topographic memory mobilized during Hamlet training but significantly improved the resistance to scopolamine-induced amnesia. We previously described that Hamlet training decreased the sensitivity of mice to the scopolamine-induced amnesic effect (Crouzier et al., 2018). Scopolamine blocked spontaneous alternation ability at the dose of 0.5 mg/kg in NT animals, but not in T animals, for which a dose of 5 mg/kg IP was required to measure a significant alteration (Crouzier et al., 2020). Pharmacological blockade of S1R by repeated administration of NE-100 prevented the effects of EE, confirming that this receptor constitutes the primary mediator of EE training action on the brain (Crouzier et al., 2020; Ruscher et al., 2011). We confirmed here that Hamlet training significantly attenuated the scopolamine-induced alternation deficit. We also analyzed the number of entries necessary to do the first alternation (NEFA), which can be considered as an index of the exploration ability of the animals based on spatial memory. It was impaired in scopolamine-treated NT animals (about 5 entries vs 3 for cognitively unpaired animals; Figure 4(b)). EE only mildly reduced NEFA. The Amylovis-201 treatment non-significantly attenuated the scopolamine-induced alternation deficit but significantly attenuated scopolamine-induced increase in NEFA. A combination of Hamlet training and drug treatment resulted in a complete blockade of scopolamine-induced alternation deficit and an increase in NEFA, showing a clear additivity of the effects. Moreover, a NE-100 treatment just before the Y-maze test session completely reversed the beneficial effects of EE and EE + Amylovis-201, suggesting that S1R is primarily involved in this protection. EE has been shown to increase brain plasticity (Van Praag et al., 2000), particularly in the Hamlet test (Crouzier et al., 2018). EE induces neural plasticity by stimulating central nervous system-wide increases in the expression levels for the immediate early genes (IEGs), by increasing expression of neurotrophins such as nerve growth factor, BDNF, and vascular endothelial growth factor (Branchi et al., 2004; During and Cao, 2006), and by stabilizing epigenetic changes in chromatin, such as DNA methylation and histone modification (Arai and Feig, 2011), with direct consequences on hippocampal neurogenesis, particularly (During and Cao, 2006; Olson et al., 2006). We selected the Hamlet training to mimic EE as it can be considered as placing animals in a more ethological environment than the lab breeding cage and allows different pathways to discover food, water, congeners, or burrow, and creates a real enrichment closer to the ethological conditions. When EE is induced by training in the Hamlet, brain plasticity is induced as shown by activation of IEGs (FosB/∆FosB, Arc) in the para-hippocampal gyrus, an increase in BDNF expression, and an increase in histone H2B and H4 acetylation observed within days or weeks of training in the hippocampus and cortex (Crouzier et al., 2018, 2020). Consequently, newborn cell proliferation and cell maturation into new neurons were increased in the hippocampal DG (Crouzier et al., 2018, 2020). We confirmed here the significant increase in cell maturation, although we could not observe a significant increase in newborn cell proliferation in Hamlet training. The major involvement of S1R activity in this pro-neurogenic effect of EE was shown by the observation that the EE effect is blocked with repeated treatment with the S1R antagonist NE-100 or in S1R KO mice (Crouzier et al., 2020). S1R agonist treatment was also described to boost neurogenesis. First, continuous administration of Cutamesine (SA4503) in rat dose-dependently enhanced cell proliferation in the sub-granular zone of the hippocampus (Lucas et al., 2008). Second, dehydroepiandrosterone (DHEA) was shown to prevent the Aβ25–35-impaired survival and dendritic growth of newborn neurons through a S1R-mediated modulation of the PI3K-Akt-mTOR-p70S6k signaling pathway (Li et al., 2010). Third, we described that a 2-week treatment with PRE-084 increased cell proliferation but not cell maturation (Crouzier et al., 2020).
In the present study, Amylovis-201 treatment alone failed to affect neurogenesis. However, only a single dose of 0.1 mg/kg PO was tested, so this result must be further explored using different dosages and administration protocols. Furthermore, the combination of EE and Amylovis-201 significantly increased cell proliferation, suggesting that the dose of the drug alone (0.1 mg/kg) was sub-effective. The EE + Amylovis-201 combination also significantly increased cell maturation. These observations confirmed that S1R sustained stimulation by a combination of S1R agonist repeated treatment and EE markedly boosts hippocampal neurogenesis. Therefore, EE induced by Hamlet training increased S1R expression in brain structures essential for cognition, such as the hippocampus and cortex, rapidly after the initiation of EE (Crouzier et al., 2020), and repeated treatment with Amylovis-201 activated S1R (García-Pupo et al., 2024), boosting numerous cellular pathways in neurons and glial cells (Maurice and Goguadze et al., 2017; Su et al., 2016; Tsai et al., 2017). The observation that an acute NE-100 treatment blocked both EE and/or Amylovis-201-induced behavioral improvements further confirmed that S1R plays a major role in brain plasticity under physiological conditions.
It must be noted that the observation that EE failed to significantly increase cell proliferation in this study was unexpected. We, using Hamlet training, and other authors, using home cage enrichment, previously observed significant increases in cell proliferation (Crouzier et al., 2020; Gage, 2025). Cell proliferation can be assessed using different markers, notably BrdU, incorporated in place of thymidine in newly synthesized DNA of dividing cells, or Ki-67, a protein selectively expressed during active phases of the cell cycle. In our previous experiments, Ki-67 systematically showed a higher increase after Hamlet training, in the +40% (Crouzier et al., 2020) to +60% (Crouzier et al., 2018) range, as compared to BrdU, from +19% (this study) to +40% (Crouzier et al., 2018). Hamlet training, therefore, only provoked a non-significant tendency to increase cell proliferation, so this observation was not in total contradiction with previous observations but rather suggested that BrdU was not the optimal marker for cell proliferation.
The EE + Amylovis-201 combination was then tested in a pathological condition, using the AD murine model induced by ICV injection of Aβ25–35 peptide. This study must be understood as an addition to a corpus of studies that: (1) demonstrated the neuroprotective activity of S1R agonists (Villard et al., 2011), (2) demonstrated that Amylovis-201 is an anti-aggregating agent and a S1R agonist (García-Pupo et al., 2024; Rivera-Marrero et al., 2020), (3) Amylovis-201 and other anti-aggregating agents or S1R agonists are protective in transgenic models of AD (Estévez-Silva et al., 2022; Martínez-Orozco et al., 2025; Rivera-Marrero et al., 2020), and (4) EE has a beneficial effect on AD pathology and on S1R activity (Arendash et al., 2004; Lazarov et al., 2005). We selected the Aβ25–35 model as it allows us to work on young animals and to rapidly and in a standardized manner to induce an AD-like pathology (Canet et al., 2023) and with high predictive value to identify symptomatic or neuroprotective drugs (Carles et al., 2025; Chumakov et al., 2015; Maurice et al., 2013; Rodríguez Cruz et al., 2017; Villard et al., 2011). Mice received the amyloid peptide just before being trained in the Hamlet and/or being treated with Amylovis-201 at 2 doses. A thorough analysis of the mice’s status and exploratory behavior in the Hamlet showed no adverse effect of the treatments, except some limited body weight gain in trained animals (they were more active) and a limited hyperactivity in Aβ25–35-treated mice (prevented by the low dose of Amylovis-201). However, when mice were tested for spontaneous alternation, an additive if not synergic effect of the combination was observed. Both readouts (spontaneous alternation and NEFA) showed some effect of the Amylovis-201 treatment, with the low dose attenuating the alternation deficit and the high dose being more effective on NEFA. So, although the primary readout (the percentage of alternation) would suggest that Amylovis-201 presented, as expected for an S1R agonist (Maurice, 2021), a bell-shaped effect, both doses were effective depending on the parameters analyzed. The EE failed to affect the Aβ25–35-induced alternation deficit but attenuated NEFA, and the combination with each dose of Amylovis-201 significantly blocked the Aβ25–35-induced alternation deficits and increase in NEFA. The combination effects were completely prevented by NE-100, again confirming that the EE-induced beneficial effect is S1R-dependent. S1R activity has been repeatedly shown to be neuroprotective in rodent models of AD. For instance, Blarcamesine prevented Aβ25–35-induced memory deficits and toxicity in mice (Villard et al., 2011); AF710B attenuated cognitive impairments, decreased BACE1 and kinases activity, neuroinflammation, soluble and insoluble Aβ1–40/42 contents and tau pathology in 3 × Tg-AD mice (Fisher et al., 2016) and in McGill-R-Thy1-APP transgenic (Tg) rats (Hall et al., 2017); and Amylovis-201 attenuated memory deficits, toxicity and Aβ1–40/42 load in Aβ25–35-treated, streptozotocin-treated, or 3xTg-AD mice (García-Pupo et al., 2024; Mercerón-Martínez et al., 2024; Martínez-Orozco et al., 2025; Rivera-Marrero et al., 2020). Moreover, the impact of EE on recovery from degenerative pathologies, notably due to cholinergic alterations, has been first shown in pharmacological models, such as kainate-induced seizures and excitotoxic injury (Young et al., 1999), then on cognition improvement in transgenic mouse models of AD (Arendash et al., 2004). Interestingly, this latter report and others (Wolf et al., 2006) showed that EE marginally affects amyloid load despite a significant effect on numerous markers of the physiopathology, like kinases, trophic factors, neuroinflammatory cytokines, have been reported. Combination with Amylovis-201 will therefore allow not only an optimal stimulation of S1R but also to ensure a significant impact on Aβ load, as the drug was first identified as an anti-aggregating agent (Rivera-Marrero et al., 2020). Indeed, and contrarily to reference S1R ligands such as the agonist PRE-084 or even the antagonist NE-100, the drug bears a naphthalene moiety interfering directly with Aβ proteins and therefore prevents Aβ aggregation and disaggregates formed oligomers (Rivera-Marrero et al., 2020; García-Pupo et al., 2024). The EE+Amylovis-201 combination is therefore expected to have a potent impact on AD physiopathology with putative disease-modifying effects. These results will have to be confirmed in more complex memory tests, including repetitive training sessions and measuring reference long-term or recognition memory, which are known to be highly affected in AD, as the present study was limited to a simple spatial working memory test. Moreover, a long-term chronic experiment in transgenic mice, such as APP/PS1, is in preparation that will allow us to compare the effects of numerous physio-pathological markers of AD of the Amylovis-201 treatment alone or in combination with EE, and in comparison with other S1R agonists like PRE-084. Such a study will allow us to confirm that the additivity of the EE + Amylovis-201 neuroprotection is observable (i) in a chronic transgenic model of AD, (ii) not gender-related, (iii) objectifiable under chronic treatment protocols, and (iv) in other behavioral tests addressing different memory processes (long-term, non-spatial, recognition memory) that were not addressed in the present study.
In conclusion, we used training in the Hamlet as a prototypic EE procedure to analyze the impact of its combination with repeated treatment with the S1R agonist Amylovis-201, and we observed an amplified effect on brain plasticity under physiological and pathological (AD) conditions. The therapeutic value of such a combination is expected to lead to a highly effective disease-modifying effect.
Supplemental Material
sj-docx-1-jop-10.1177_02698811261458343 – Supplemental material for Enhanced cognitive protection through the combination of Amylovis-201, an anti-amyloidogenic agent with sigma-1 receptor agonist activity, and environmental enrichment in mice
Supplemental material, sj-docx-1-jop-10.1177_02698811261458343 for Enhanced cognitive protection through the combination of Amylovis-201, an anti-amyloidogenic agent with sigma-1 receptor agonist activity, and environmental enrichment in mice by Roberto Menéndez Soto del Valle, Charlyne Barry-Simonnet, Lison Verjus, Lucie Crouzier, Sairys Peña Alcolea, Marquiza Sablón Carrazana, Tangui Maurice and Chryslaine Rodríguez-Tanty in Journal of Psychopharmacology
Footnotes
Author contributions
Roberto Menéndez Soto del Valle: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. Charlyne Barry-Simonnet: Investigation, Methodology. Lison Verjus: Investigation. Lucie Crouzier: Formal analysis, Investigation, Methodology, Writing – review & editing. Sairys Peña Alcolea: Investigation. Marquiza Sablón Carrazana: Writing – review & editing. Tangui Maurice: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing. Chryslaine Rodríguez-Tanty: Conceptualization, Funding acquisition, Project administration, Validation, Writing – review & editing.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by PHC Carlos J. Finlay programs from Campus France (projects 47069SA to TM and CRT and 49699YF to TM and RMSDV) and Fondo de Cooperación Triangular Unión Europea-Cuba-América Latina y el Caribe (Nº 3058392, ADELANTE, AECID, Ministerio de Asuntos Exteriores y de Cooperación de España. The authors thank the CECEMA animal facility of the University of Montpellier. The work has been supported through the European Cooperation in Science and Technology (COST) Association, SIGMA1_EUROPE action under No. CA23156.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.