
Abstract
Angiogenesis during post-stroke neurovascular recovery is linked to oligodendrocyte precursor cells (OPCs) phenotypic shifts. Oridonin, a natural diterpenoid, has the potential to regulate post-stroke angiogenesis, but its specific mechanism remains unclear. Intraperitoneal injection of oridonin significantly promoted angiogenesis in the ischemic penumbra of stroke model mice. However, endothelial cell proliferation, sprouting, and tube formation assays demonstrated that oridonin exerted no direct regulatory effect on endothelial cell-triggered angiogenesis in vitro. Unexpectedly, oridonin notably promoted perivascular OPCs proliferation in the ischemic penumbra and enhanced the secretion of the pro-angiogenic factor Wnt7a by immunofluorescence staining and enzyme-linked immunosorbent assay (ELISA) analysis. Meanwhile, the conditioned medium from oridonin-treated OPCs enhanced the angiogenic effect of endothelial cells, which was reversed by inhibition of the Wnt7a/β-catenin pathway. Mechanistically, drug affinity responsive target stability (DARTS) and microscale thermophoresis (MST) assays confirmed that oridonin directly bound to α-ketoglutarate-dependent prolyl hydroxylase 2 (PHD2), thereby inhibiting the ubiquitination and degradation of hypoxia-inducible factor 1-alpha (HIF-1α) in OPCs and regulating their proliferation and pro-angiogenic effects. Collectively, oridonin targets PHD2 to inhibit HIF-1α ubiquitination and degradation in OPCs, thereby facilitating OPC proliferation and activating the Wnt7a/β-catenin pathway to promote endothelial angiogenesis after stroke.
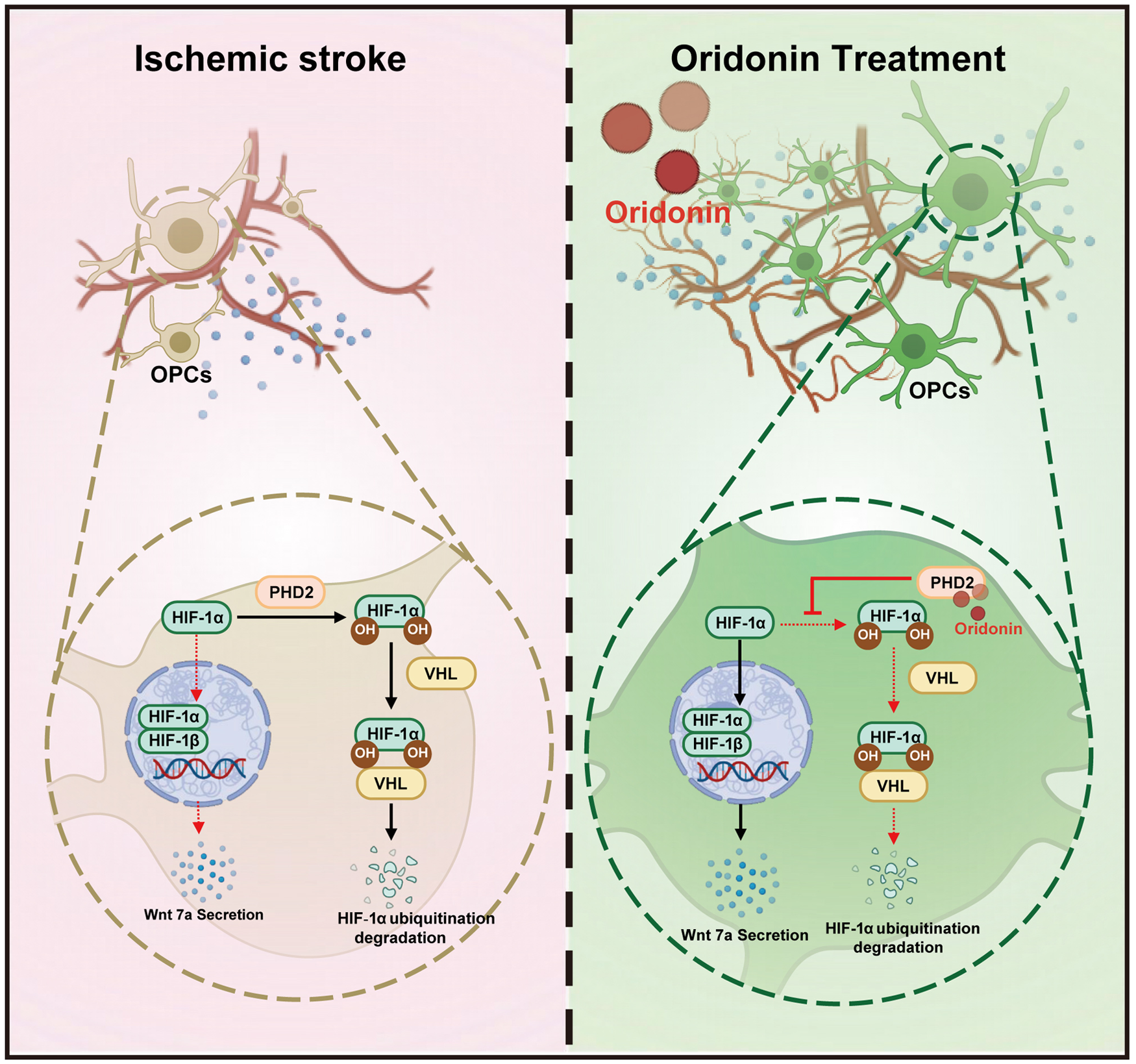
Introduction
Ischemic stroke, accounting for approximately 85% of all stroke cases, results from a reduction or cessation of cerebral blood flow due to arterial occlusion caused by thrombosis or embolism. 1 Although new therapy strategies and drugs for ischemic stroke are constantly developing, it is still the second leading cause of mortality and third leading cause of disability worldwide.2,3 Currently, acute focal stroke is managed using drug thrombolysis and mechanical thrombolysis, but these therapy methods are limited to a narrow treatment time window and secondary injuries of hemorrhage transformation.4,5 Therefore, developing novel regenerative therapies targeting both mortality reduction and neurological rewiring is imperative to address the unmet clinical needs in ischemic stroke recovery.
Emerging research highlights the critical role of neurovascular networks in brain function, underscoring the importance of bidirectional communication between neurons and blood vessels. 6 Certainly, angiogenesis serves as a critical neuroprotective mechanism that facilitates neural regeneration and functional restoration following ischemic stroke. 7 Promoting angiogenesis during the recovery period of ischemic stroke could help to improve collateral reperfusion in the ischemic penumbra, promote nerve regeneration, and improve prognosis and prolong survival time.8,9 As is widely acknowledged, early-phase ischemia triggers localized angiogenesis, which facilitates macrophage recruitment and necrotic tissue clearance via newly formed microvasculature. 10 However, this acute stage angiogenesis is transient and typically diminishes within weeks post-ischemia. 10 At present, there are limited tools available to promote angiogenesis in the recovery phase after ischemic stroke.
In recent years, it has been reported that oligodendrocyte precursor cells (OPCs) are involved in the formation of white matter blood vessels during development, and physical interactions between OPCs and endothelial cells during angiogenesis have been found.11–13 Moreover, transplantation of OPC into the ischemic side of the brain can promote angiogenesis. 14 Therefore, therapeutic modulation of angiogenesis via targeted OPC regulation emerges as a promising novel strategy for ischemic stroke intervention. Oridonin, which belongs to the kauridine-type tetracyclic diterpenoid natural organic compounds, has antioxidant, anti-inflammatory and anti-tumor effects in a variety of diseases.15–18 Our previous studies have found that oridonin inhibited oxidative stress and reduced endothelial cell apoptosis in the acute phase of ischemic stroke. 17 Moreover, we found that oridonin could increase vascular continuity, microvessel density and branch node number in the ischemic penumbra during ischemic stroke recovery, suggesting that oridonin could promote angiogenesis. However, our preliminary research results show that oridonin failed to affect the proliferative capacity of brain microvascular endothelial cells, the main effector cells of angiogenesis.
Here, we hypothesize that oridonin might act on other types of cells rather than microvascular endothelial cells to promote angiogenesis in ischemic stroke. Whether OPCs serve as the primary cellular target mediating the pro-angiogenic effects of oridonin remains to be elucidated. Therefore, this study aimed to explore the impact of oridonin on OPCs and to uncover the molecular mechanisms underlying its pro-angiogenic effects in ischemic stroke.
Materials and methods
Experimental animals and administration
Male C57BL/6J mice, 8 weeks old and weighing 20 ± 2 g, were obtained from the Animal Resource Center of the Faculty of Medicine, Nanjing Medical University for the present study. All animal procedures were conducted in strict accordance with the Guide for the Care and Use of Laboratory Animals and the regulations of the Animal Protection and Use Committee of Jiangsu Experimental Animal Association. All experimental protocols were approved by the Institutional Animal Care and Ethics Committee (IACUC) of Nanjing Medical University (approval number: IACUC-2203042). This study complied fully with the ARRIVE guidelines 2.0 for reporting animal experiments. All animals were randomly divided into the following three groups by a randomized block design: Sham + Vehicle group, tMCAO + Vehicle group, and tMCAO + oridonin group. For the Sham group, mice were subjected to the same procedures as the other groups except for inserting a filament. In the tMCAO + oridonin group, the mice were intraperitoneally administered with oridonin once per day at a dose of 20 mg per kg after reperfusion.
Reagents and drugs
Oridonin (#T2790; TargetMol, Shanghai, China) was dissolved in dimethyl sulfoxide (DMSO) for storage (500 mM) and then further diluted to working concentrations in vitro. XAV-939 (#T1878; TargetMol, Shanghai, China) was dissolved in DMSO for storage (10 mM) and then diluted to a working concentration of 10 µM to treat bEnd3 cells.
Preparation for the transient focal cerebral ischemia and reperfusion model
The protocol of transient middle cerebral artery occlusion (tMCAO) was conducted by using the intraluminal filament technique as previously described. 19 Briefly, the common carotid artery (CCA), external carotid artery (ECA) and internal carotid artery (ICA) were carefully exposed via a midline neck incision. The CCA was temporarily occluded, and a nylon monofilament (0.18 ± 0.01 mm, L1800; Guangzhou Jialing Biotechnology) was inserted into the ICA via the ECA to the middle cerebral artery and left for 45 min. After 45 min of occlusion, the monofilament was withdrawn for reperfusion. Local cerebral blood flow was measured using a MoorFLPI Full-field Laser Perfusion Imager (MoorFLPI-2; Gene&I, Beijing, China) to confirm successful establishment of the transient focal cerebral ischemia–reperfusion model. In this study, the infarction core and penumbra were divided as follows: the infarction hemisphere was 2 mm apart from the sagittal plane and 30° oblique to the sagittal plane, with the lateral cortex as the ischemic core and the medial cortex as the ischemic penumbra. 20
Neurological function assessment
Neurological functional deficits in mice were blindly evaluated via four validated behavioral tests: neurological scoring, corner test, adhesive removal test, and fear conditioning test (FCT), which reliably quantify sensorimotor, postural, and cognitive memory impairments. Neurological function was scored using the modified Garcia score covering body proprioception, spontaneous activity, limb symmetry, lateral turning, forelimb locomotion, and climbing behavior. 21 In the corner test, mice were placed in a 30° corner; full turns with rearing were recorded, and right turns were counted over 10 trials. For the adhesive removal test, 2 × 3 mm adhesive strips were attached to bilateral forepaws, and the latency to contact and remove the strips was measured. Cognitive and memory functions were assessed by FCT with a high-sensitivity weight transducer system, where mice were habituated for 10 min, trained with a 30-s tone paired with a 1-s foot shock, and tested for 5 min without shock, and the percentage of freezing behavior was analyzed.
Density of vessels measurement
Sterile saline with 10 mg/mL FITC–dextran 2000 kDa (#FD2000S; Sigma–Aldrich) was prepared and stored protected from light. Mice were anesthetized and injected intravenously with 0.1 mL of the solution. After 30 min of circulation, brains were harvested, fixed in 4% PFA overnight, and sectioned (180 μm) using a vibratome. Sections were mounted, coverslipped, and imaged via Z-stack fluorescence microscopy (light-protected throughout). 3D reconstruction was performed with Imaris 9.0.1, and vascular density/junctions were quantified using AngioTool. 22
Extraction and culturing of primary oligodendrocyte precursor cells
Primary OPCs were obtained from cerebral cortices of 1-day-old postnatal C57BL/6J mice as previously described. 23 Briefly, primary OPCs were isolated from cerebral cortices of postnatal 1-day-old C57BL/6J mice. Cortices were dissected, digested with 0.25% trypsin/EDTA and 0.025% DNase I at 37 °C for 15 min, centrifuged, resuspended in DMEM, and filtered through an 80-μm strainer. Cells were seeded into polylysine-coated flasks and cultured for 8–10 days. Microglia were removed by shaking (180–220 rpm, 30–60 min), followed by overnight shaking (200–220 rpm) after PBS washes. The supernatant was collected, incubated in uncoated vials for 1 h, and centrifuged (1000 rpm, 5 min, 4 °C). Purified OPCs were plated in polylysine-coated flasks/plates (9–15 × 103 cells/cm2) with OPC proliferation medium (DMEM-F12, 1% N2, 2% B27, 1% P/S, 40 ng/mL PDGF-AA). Medium was replaced fully on day 1 and every 1.5 days thereafter.
Oxygen–glucose deprivation and reoxygenation (OGD/R)
The ODG/R was performed as described previously. 23 Before the OGD experiment, OPC and bEnd.3 cells were cultured with DMEM complete medium in an incubator (Thermo Fisher Scientific, Waltham, MA, USA) with 95% air and 5% CO2 at 37 °C. During the OGD procedure, cells were treated with deoxygenated conditions (95% N2 and 5% CO2) and glucose-free DMEM (#11966-025; Life Technologies, MD, USA) for 5 or 6 h. After OGD, the cells were transferred to a normoxic incubator and replaced with normal DMEM medium.
Cell culture and drug treatment
Mouse primary OPCs and the mouse brain microvascular endothelial cell line bEnd.3 were used in the in vitro assay. The cells were cultured with DMEM medium, added with 10% FBS (#10099141; Gibco, NY, USA) and 1% penicillin/streptomycin in an incubator (95% air and 5% CO2) at 37 °C. The cells were seeded into 24-well plates before exposure to OGD treatment for 5 or 6 h. When reperfusion was performed, oridonin was added to the normal DMEM medium simultaneously. After OGD/R and drug treatment, the biochemical indicators were measured and subsequent processes were carried out.
Establishment of non-contact co-culture system
The bEnd.3 cell line was provided by Procell Life Science & Technology Co., Ltd. (Wuhan, China). OPCs were seeded at 1.2 × 105 cells/well in 6-well plates (1.5 mL medium/well). After OGD/R and oridonin treatment, supernatants were collected at 24 h, centrifuged (1000g, 5 min), and stored as OPC-conditioned media (24 h-Veh-OPC-CM and 24 h-Ori-OPC-CM) at −80 °C. bEnd.3 cells were inoculated with 4 × 104 cells in 24-well plates. The control group was not treated; the OGD/R group was added with normal medium during reoxygenation, and the OGD/R + OPC conditioned medium group was added with OPC-CM of the above groups during reoxygenation.
Nuclear–cytoplasmic fractionation assay
Nuclear–cytoplasmic fractionation was conducted following the kit manufacturer’s protocol. Briefly, cultured cells (5 × 106 cells/mL) were harvested by centrifugation at 800 rpm for 3 min (4 °C), then washed twice with ice-cold PBS. Cell pellets were resuspended in cytoplasmic lysis buffer, lysed on ice and centrifuged, with the supernatant collected as the cytoplasmic fraction. The remaining pellet was resuspended in nuclear lysis buffer, incubated on ice for 30 min to 1 h, and centrifuged to harvest the nuclear fraction. Protease and phosphatase inhibitors were added to both fractions. Protein concentrations were determined by BCA assay. Samples were denatured by boiling with loading buffer and subjected to Western blotting to examine target protein expression and subcellular distribution.
Co-immunoprecipitation assay
Total protein was isolated from cells using a non-denaturing lysis buffer with protease inhibitors. The protein lysate was incubated with anti-HIF-1α antibody (#Ab51608; Abcam) and rabbit IgG (#Ab125938; Abcam) at 4 °C overnight. Then, Protein A/G agarose beads (#B23202; Selleckchem) were added and incubated at 4 °C for 4 h. The immunoprecipitates were collected by centrifugation and washed with PBS. The mixture was subjected to Western blot analysis.
Cell siRNA and plasmid transfection
Cells at 60% density were transfected with Wnt7a siRNA or PHD2 overexpression plasmid using Lipofectamine 3000 (#L3000008; Thermo Fisher Scientific), following the provided instructions. After 12 h of transfection, the culture medium was replaced with fresh medium, and the cells were further incubated for 40 h before downstream analysis. Western Blotting was used to verify transfection efficiency. The siRNA sequence of Wnt7a is as follows: sense (5′–3′)-AGGAGAACAUGAAGCUGGAGUGUAA; antisense (5′–3′)-UUACACUCCAGCUUCAUGUUCUCCU. The pLenti-CMV-mEgln1-3*Myc-GFP-Puro plasmid (#PPL50728-4a; Public Protein/Plasmid Library) was used for the overexpression assay.
Measurement of cell proliferation, spheroid sprouting, and tube formation
In EdU proliferation assay, bEnd.3 cells were seeded at 4 × 104 cells/well in 24-well plates. According to the EdU-594 kit instructions, 10 μM EdU working solution was added 2 h before cell harvest. After staining, samples were mounted and imaged via fluorescence microscopy. For tube formation assay, bEnd.3 cells (7 × 104 cells/well) were plated onto Corning Matrigel (#354234) in 24-well plates. Tube networks were photographed at 4 h, and tube number/length were quantified using ImageJ. 24 In spheroid sprouting assay, bEnd.3 cells were resuspended in DMEM supplemented with 0.25% methylcellulose and seeded at 1.5 × 105 cells/mL in low-adhesion 24-well spheroid microplates. After 24 h incubation, collected spheroid sediments were resuspended in Matrigel, plated and allowed to polymerize for 30 min, then supplemented with DMEM. 25 Following another 24 h culture, the sprouting area was analyzed by ImageJ.
Pull-down assay
For the pull-down assay, cell lysates were prepared using non-denaturing lysis buffer with protease inhibitors, centrifuged (1200g, 15 min), and supernatants collected. Input samples (50 μL) were reserved. Pre-cleared lysates were incubated with streptavidin beads (4 °C, 2 h) to remove nonspecific binders. Equal protein amounts were then incubated with oridonin/Bio-ori and beads (2 h). Post-incubation, beads were washed (PBS/0.1% Tween-20), boiled in loading buffer, and analyzed by Western blot.
Molecular docking
Molecular docking and virtual screening were conducted using AutoDock Tools software (version 1.5.7). The three-dimensional crystal structure of PHD2 (PDB ID: 6YW1) was acquired from the Research Collaboratory for Structural Bioinformatics Protein Data Bank (RCSB PDB). The canonical molecular structure of oridonin (PubChem CID: 5321010) was retrieved from the PubChem Compound Database. The optimal ligand-receptor complex generated through molecular docking simulations was subsequently subjected to structural visualization and analysis utilizing Discovery Studio 2021 Client and PyMOL (version 2.2.0) for detailed intermolecular interaction assessment.
Drug affinity responsive target stability (DARTS)
Drug affinity responsive target stability (DARTS) was conducted as previously described. 26 Lysates from OPC were incubated with or without oridonin (5 μM) for 1 h at room temperature. Samples were boiled immediately after adding loading buffer to stop the digestion. Each sample was loaded onto SDS-PAGE for western blotting.
PHD2-His recombinant protein preparation
PHD2-His recombinant protein expression for the binding assay was performed as described previously. 27 The recombinant protein preparation process involves gene cloning into expression vectors with optimized codons and affinity tags (His-tag) for host systems like E. coli or mammalian cells, followed by protein expression induced by IPTG. Subsequent purification employs Ni-NTA resin and AKTA, with final validation via SDS–PAGE to achieve >95% purity.
Microscale thermophoresis (MST) assay
The above-mentioned recombinant fusion proteins were fluorescently labeled with the Monolith His-Tag Labeling Kit (NanoTemper Technologies, Munich, Germany) following the manufacturer’s protocol. For the binding studies, recombinant PHD2 was prepared at a concentration of 50 nM, while oridonin was prepared at 500 μM. Quantitative analysis of molecular interaction affinity was performed using MST technology with a Monolith NT.115 instrument (NanoTemper Technologies, Munich, Germany), employing well-established fluorescence-based methodologies for binding characterization.
Western blotting (WB)
Protein lysates were extracted using RIPA buffer (Beyotime Biotechnology, Shanghai, China) and quantified via BCA assay. SDS-PAGE separated proteins, which were transferred to PVDF membranes (Roche, Mannheim, Germany). After blocking with 5% BSA/TBST (2 h, room temperature), membranes were incubated with primary antibodies (12 h, 4 °C), followed by HRP-conjugated secondary antibodies (2 h, room temperature). Protein bands were visualized by ECL using a Tanon 5200 system and quantified with ImageJ. Primary antibodies used in this study for WB were as follows: β-actin (1:1000, #66009-1-1g; Proteintech), β-tubulin (1:1000, #10094-1-AP; Proteintech), HIF-1α (1:1000, #Ab308433; Abcam), Wnt7a (1:1000, #Ab274321; Abcam), β-catenin (1:1000, #51067-2-AP; Proteintech), Histone H3 (1:1000, #17168-1-AP; Proteintech).
Immunofluorescence staining
Mice brains were postfixed overnight (4 °C), dehydrated, and sectioned (20 µm). Sections were permeabilized with 0.2% Triton-X-100 (3 min), blocked with 10% BSA (1 h, 37 °C), and incubated with primary antibodies (4 °C overnight) including CD31 (1:100, #AF3628-SP, R&D Systems), platelet-derived growth factor receptor alpha (PDGFRα) (1:200, #Ab96569; Abcam), PDGFRα (1:500, #Ab203491; Abcam), Olig2 (1:100, #AF2418, R&D Systems), IB4 (1:1000, #13838S; Invitrogen), Ki67 (1:200, #Ab15580; Abcam), and α-SMA (1:200, #80008-1-RR; Proteintech). Slices were washed with PBST and incubated with secondary antibodies (1:1000, Invitrogen) for 1 h at RT. DAPI (#C3619-PI19B, Southern Biotech) was added for 10 min (light-protected). The quantification of images was analyzed with ImageJ software as previously described.
Quantitative real-time PCR
Total RNA was extracted with TRIzol reagent (Invitrogen Life Technologies, CA, USA), quantified by NanoDrop 2000 (NanoDrop Technologies, Thermo Scientific, USA), and reverse-transcribed using SuperScript III (Invitrogen, USA). qPCR was performed on a QuantStudio 5 system with SYBR Green Master Mix and a CFX-96 instrument. Melting curve analysis confirmed specificity, and GAPDH-normalized data were analyzed via the 2−ΔΔCt method. Primer sequences for RT-PCR were as follows:
VEGF-α (F: ACATTGGCTCACTTCCAGAAACAC; R: GGTTGGAACCGGCATCTTTATC)
HIF-1α (F: TGACTGTGCACCTACTATGTCACTT; R: GGTCAGCTGTGGTAATCCACTC)
Ang-1 (F: CAGCATCCTGCAGAAGCAAC; R: TCTCTTTTTCCTCCCTTTAGCA)
Wnt7a (F: GACAAATACAACGAGGCCGT; R: GGCTGTCTTATTGCAGGCTC)
GAPDH (F: TGTGTCCGTCGTGGATCTGA; R: CCTGCTTCACCACCTTCTTGAT)
Statistical analysis
Data were shown as mean ± SD. Two statistical methods were used, including one-way ANOVA and Kruskal–Wallis test. Normality of grouped data was assessed using the Shapiro–Wilk test (p > 0.05 indicated normality). For normally distributed data, one-way ANOVA was applied (p ⩽ 0.05 significant difference). Non-normal data were analyzed via the Kruskal–Wallis test (p ⩽ 0.05 significant difference). GraphPad Prism 8.02 was applied to analyze statistical significance.
Results
Oridonin significantly accelerated angiogenesis in the penumbra area after tMCAO/reperfusion
We initially established a mouse model of cerebral ischemia/reperfusion (I/R) injury via transient middle cerebral artery occlusion (tMCAO). Cerebral blood flow was monitored in real time using a laser Doppler flowmeter to verify successful model establishment (Figure 1(a)). Oridonin (Figure 1(b)) administration was initiated concurrently with reperfusion. We first evaluated the effects of oridonin treatment on neurological and cognitive impairments in mice during the recovery phase after tMCAO surgery, using the Garcia score, corner test, adhesive removal test, and fear conditioning test (FCT). The results showed that oridonin treatment markedly improved the neurological Garcia score in mice at 14 days post-tMCAO (Figure 1(c)). According to the corner test and adhesive removal test, oridonin significantly reduced the percentage of right turns and shortened the latency to remove adhesive tapes in tMCAO mice, indicating that oridonin ameliorates sensorimotor dysfunction after ischemic stroke (Figure 1(d) and (e)). Moreover, oridonin treatment greatly alleviated cognitive functional deficits in tMCAO mice as revealed by the FCT (Figure 1(f) and (g)). Collectively, these findings demonstrate that oridonin treatment facilitates neurological functional repair during the recovery period following ischemic stroke.
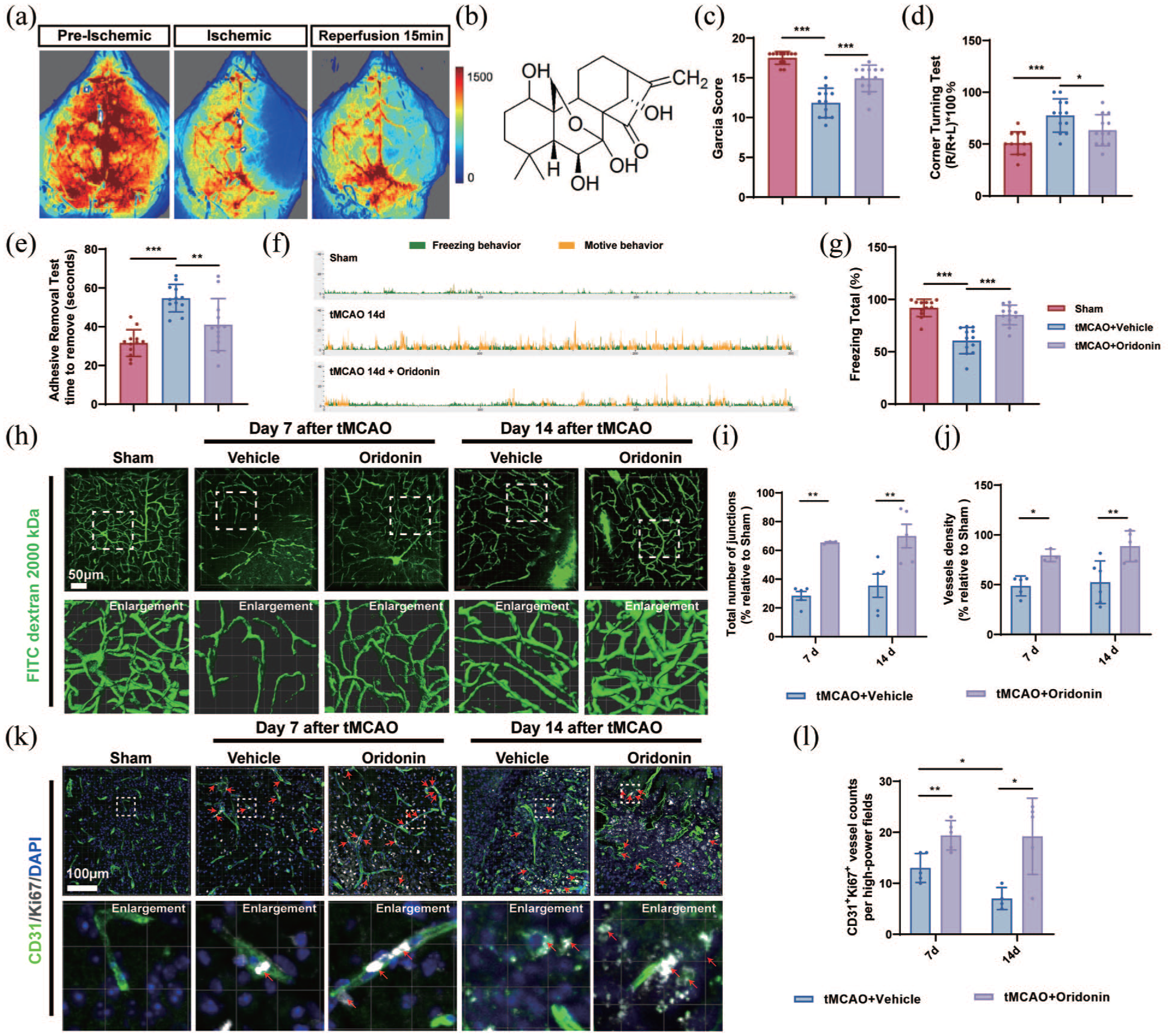
Oridonin significantly enhanced angiogenesis in the penumbra area of ischemic stroke model mice: (a) the pattern diagram of successful model validation was captured from a laser Doppler blood flow meter, (b) the molecular structure of oridonin, (c) neurological deficit was assessed using Garcia score in I/R 14-day + vehicle and I/R 14-day + oridonin groups or in sham groups, (d, e) the corner test and adhesive removal test was performed in I/R 14-day + vehicle and I/R 14-day + oridonin groups or in sham groups, (f, g) representative images of the motive behavior and the freezing behavior in FCT test and the percentage of freezing total was analyzed in I/R 14-day + vehicle and I/R 14-day + oridonin groups or in sham groups, (h) representative confocal microscopy images and 3D reconstruction of FITC–dextran 2000 kDa signal (green) in sham and penumbral regions at 7 and 14 days after tMCAO. Scale bar = 50 μm, (i) quantitative analysis for the number of junctions, (j) vessel density of vascular structure in ischemic penumbra, (k) CD31 (green) and Ki67 (white) co-localization was used to observe the newly formed microvessels in the penumbral area at 7 and 14 days after tMCAO. Scale bar = 100 μm, and (l) quantitative analysis was conducted to determine the co-localization for each group. Four fields of view were captured and averaged per sample for all quantitative analyses. Statistical differences were analyzed by two-way ANOVA and Sidak’s multiple comparisons test. Data are presented as mean ± SD.
To assess angiogenesis and visualize vascular morphology within the ischemic penumbra, mice from each group underwent intravenous injection of 2000 kDa FITC-dextran. Our results showed that vascular continuity and branching architecture remained well-organized in the sham group. By contrast, in the ischemic penumbra of tMCAO mice, vascular continuity was severely disrupted, accompanied by compensatory revascularization at days 7 and 14 after reperfusion compared with the sham group. Meanwhile, quantitative analysis revealed that the total number of vascular junctions and vessel density increased progressively over time following tMCAO surgery (Figure 1(h)–(j)). Unexpectedly, we found that oridonin treatment significantly attenuated the decrease in vascular density and the number of junctions in the ischemic penumbra area, which demonstrated its potential ability to promote angiogenesis after tMCAO/reperfusion (Figure 1(h)–(j)). Since oridonin reduces endothelial cell death during the acute phase after ischemic stroke, 17 the increased vascular density observed in the oridonin-treated group on days 7 and 14 may be attributed to the alleviation of endothelial cell loss in the acute stage rather than angiogenesis. To verify whether oridonin could promote angiogenesis during the recovery phase, we initiated oridonin administration starting on day 3 after tMCAO surgery and examined vascular density at days 7 and 14 (Figure S1(A)). The results revealed that even without intervention in the first 3 days after tMCAO, oridonin still effectively elevated vascular density and the total number of vascular junctions in the ischemic penumbra on days 7 and 14 (Figure S1(B)–(D)). Furthermore, CD31+/Ki67+ immunofluorescence co‑staining was performed to directly detect vascular endothelial cell proliferation, thereby reflecting angiogenic activity. And the results showed that oridonin administration markedly increased the number of CD31+/Ki67+ cells at 7 and 14 days after tMCAO compared with the tMCAO group (Figure 1(k) and (l)). To clarify the long-term functional sustainability of oridonin-induced angiogenesis, α-SMA and CD31 double immunofluorescence staining was used to assess pericyte coverage and vascular maturation at 21 days after tMCAO. Compared with the tMCAO group, oridonin significantly increased perivascular α-SMA coverage (Figure S1(E) and (F)), suggesting that oridonin-induced neovessels can mature into functionally stable vasculature during the late recovery phase. Taken together, these results indicated that oridonin has the potential to promote angiogenesis in the mouse penumbra area after tMCAO.
Oridonin promoted angiogenesis through an endothelial cell-independent pathway in vitro
Current evidence demonstrates that post-ischemic angiogenesis predominantly occurs through sprouting angiogenesis, a highly coordinated multistage process. This complex biological cascade involves sequential events including endothelial cell proliferation, polarized migration, sprout elongation with specialized tip and stalk cell differentiation, and ultimately lumenogenesis, which collectively drive the establishment of a functional and structurally mature vascular network. 28 In order to determine whether oridonin exerted its role in promoting angiogenesis by directly acting on cerebral vascular endothelial cells, we examined the effects of oridonin on proliferation, sprouting and tube-forming behavior of vascular endothelial cells in vitro. The results of the EdU-labeled proliferation experiment showed that oridonin did not attenuate the reduction of endothelial cells caused by OGD/R (Figure 2(a) and (b)), suggesting that oridonin exerted the pro-angiogenesis effects not via promoting endothelial cell proliferation. Meanwhile, the tube formation assay and sprouting assay further demonstrated that oridonin-induced angiogenesis occurred independently of direct endothelial cell activation, which displayed as oridonin treatment did not improve the total branch length and the number of junctions in the tube formation experiment and the ratio of sprouts to apheroid in the sprouting experiment (Figure 2(c)–(g)). These findings suggest that oridonin promotes angiogenesis via indirect mechanisms involving other angiogenesis-associated cell types during the ischemic stroke recovery phase.
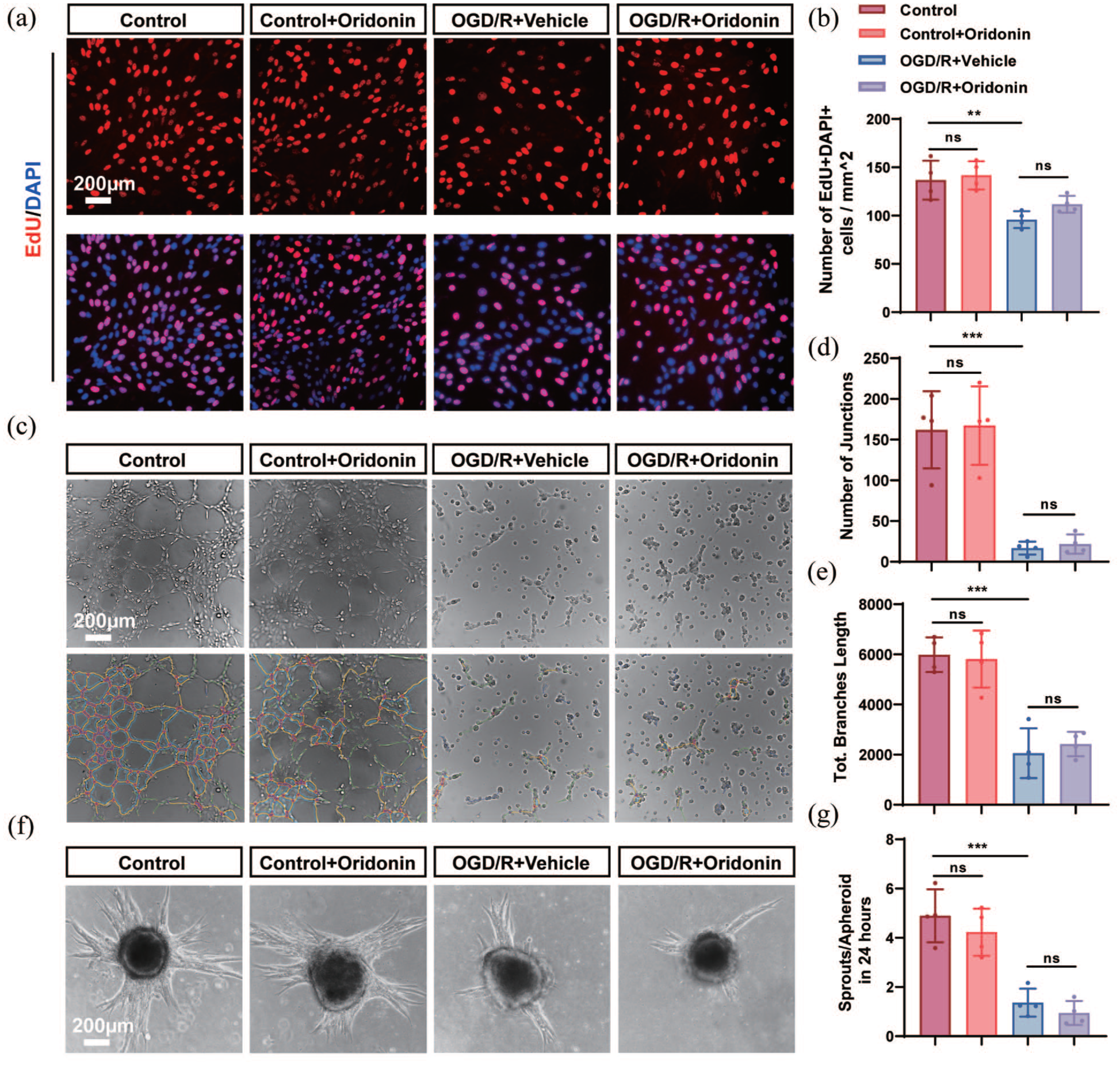
Oridonin promoted angiogenesis through an endothelial cell-independent pathway in vitro: (a) representative confocal microscopy images for EdU-labeled cell proliferation (red) of endothelial cells, (b) quantitative analysis for the number of EdU+DAPI+ cells in each view. Scale bar = 200 μm, (c) representative images showing the tube formation of bEnd.3 cells in each group. Scale bar = 200 μm, (d) total branch length, (e) the number of junctions of the tube-forming assay were quantitatively analyzed in each group, (f) representative images of the sprouting assay in each group. Scale bar = 200 μm, and (g) quantitative analysis was conducted to determine sprouts/apheroid in 24 h for each group. Three fields of view were captured and averaged per sample for all quantitative analyses. Statistical differences were analyzed by one-way ANOVA and Tukey’s multiple comparisons test. Data are presented as mean ± SD.
Oridonin increased the number of perivascular OPCs in the penumbra area after tMCAO/reperfusion
It has been reported that the maintenance of revascularization via vascular remodeling is contingent upon progenitor cell populations and necessitates direct cellular interactions between OPCs and endothelial cells throughout angiogenesis.29,30 Therefore, we speculated whether OPCs serve as the primary cellular target mediating pro-angiogenic effects of oridonin. In order to observe the physical distribution and density of OPCs around microvessels in ischemic penumbra at different time points after tMCAO/reperfusion, we used PDGFRα and Olig2, highly expressed on OPC cells, and vascular endothelial cell markers IB4 and CD31 for immunofluorescence co-localization, respectively. Three-dimensional imaging and reconstruction revealed two distinct subtypes of OPCs: (a) perivascular OPCs, which envelop microvessels with intricate cellular processes, and (b) parenchymal OPCs, which are spatially segregated from vascular structures (Figure 3(a)). Given the distinct spatial distribution of the two OPC subtypes, we quantitatively assessed the temporal dynamics of microvascular density and OPCs distribution within the ischemic penumbra at 1, 3, 7, and 14 days after tMCAO/reperfusion. The results showed that the vascular density in the ischemic penumbra area increased significantly and continued until the 14th day, and there were significant differences in vascular density at the 7th and 14th days compared with the 1st and 3rd days after reperfusion (Figure 3(b) and (c)). Notably, our analysis demonstrated a progressive increase in perivascular OPC density from days 1 to 14 post-tMCAO/reperfusion, with the number of OPCs significantly surging and morphological changes including enlarged cell bodies, enhanced branching, and vascular-like structural remodeling at day 14 after tMCAO/reperfusion (Figure 3(b) and (d)). As expected, we found a significant positive correlation between microvessel density and the number of OPCs after tMCAO/reperfusion (Figure 3(e)). Collectively, these findings demonstrate a significant positive correlation between perivascular OPC abundance and vascular density within the ischemic penumbra, indicating a potential functional role for perivascular OPCs in preserving microvascular integrity during post-stroke recovery.
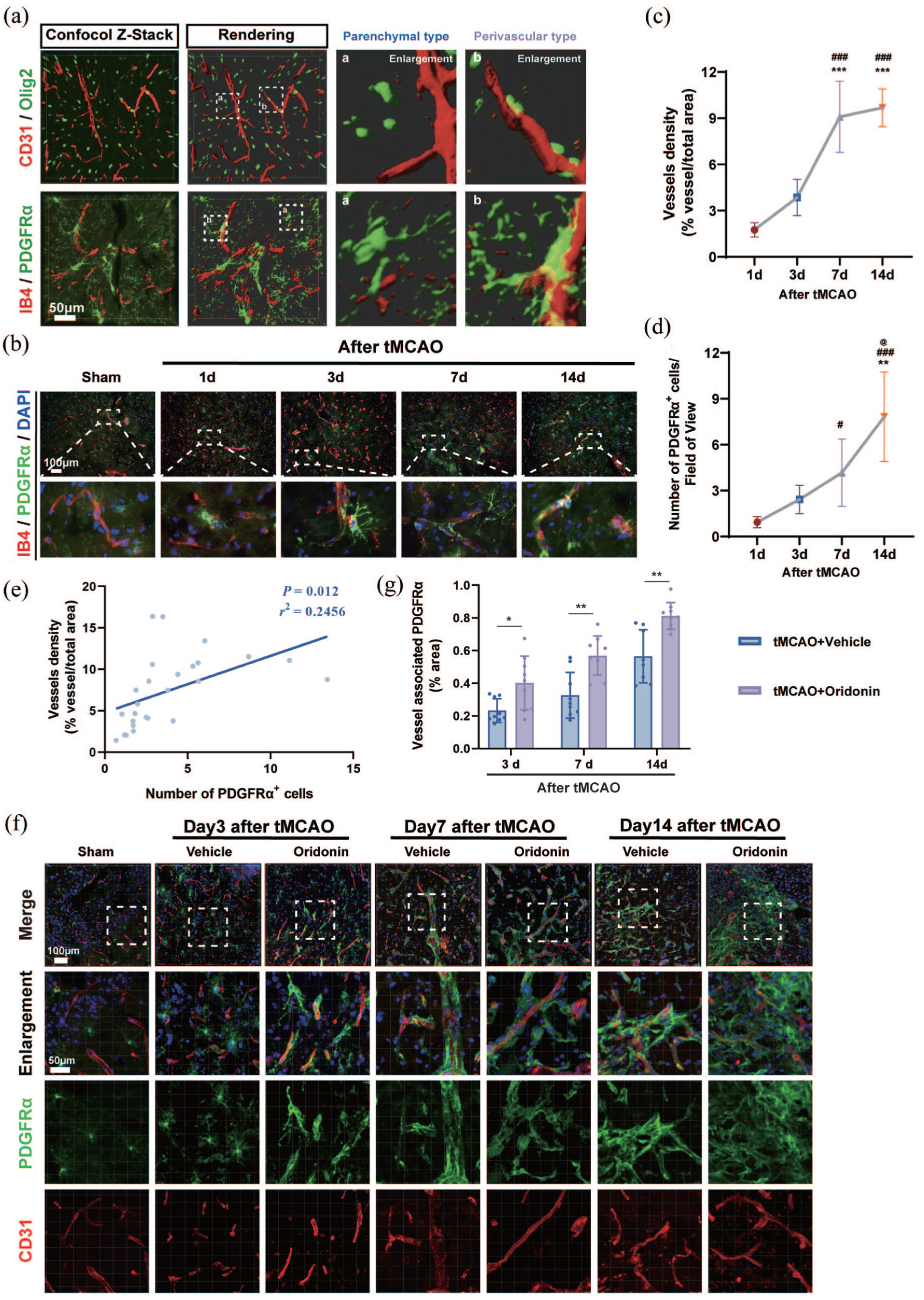
Oridonin increased the number of perivascular OPCs in the penumbra area after ischemic stroke: (a) representative z-stack confocal images and 3D-reconstruction for immunostaining of PDGFRα (green), CD31 (red), Olig2 (green), and IB4 (red) of the penumbra area at day 3 after tMCAO. Scale bar = 50 μm, (b) IB4 (red)/PDGFRα (green)/DAPI (blue) triple labeling of penumbral regions in the sham group and tMCAO group at 1, 3, 7, and 14 days after ischemic stroke. Scale bar = 100 μm, (c, d) quantitative analysis of the microvascular density and the number of OPCs of the penumbra area from days 1 to 14 after tMCAO for each group. Statistical differences were analyzed by one-way ANOVA and Tukey’s multiple comparisons test. Data are presented as mean ± SD. **p < 0.01. ***p < 0.001 versus tMCAO 1-day group. #p < 0.05. ###p < 0.001 versus tMCAO 3-day group. @p < 0.05 versus tMCAO 7-day group, (e) the correlation analysis between the number of PDGFRα+ OPCs and vascular density demonstrates a significant linear correlation (r2 = 0.2456, p = 0.012), (f) representative confocal images of PDGFRα (green) and CD31 (red) immunostaining in ischemic penumbra at 3, 7, and 14 days after tMCAO. Scale bar = 100 μm, and (g) fluorescence intensity analysis of vessel-associated PDGFRα was performed for the percentage of translocation of vessel-associated OPCs. Statistical differences were analyzed by two-way ANOVA and Sidak’s multiple comparisons test. Data are presented as mean ± SD. *p < 0.05. **p < 0.01.
To further explore whether the effect of oridonin on the positive relationship between perivascular OPC and microvessels, as well as the morphological changes, the perivascular PDGFRα+ OPCs at the 3, 7, and 14 days post-tMCAO were quantified in the ischemic penumbra area. Strikingly, while the majority of OPC are of the parenchymal subtype under normal conditions, an increase in the number of perivascular OPC was observed at 7 days after cerebral ischemia. And oridonin administration could promote the formation of vascular OPC in the ischemic penumbra on the third day, and the amount of perivascular OPC was significantly higher than that in the tMCAO group at corresponding time points (Figure 3(f) and (g)). These results indicate that oridonin treatment increase the number of perivascular OPC in the penumbra area of ischemic stroke model mice, which may indirectly promote the angiogenesis process after ischemic stroke.
Oridonin significantly enhanced the proliferation of OPCs after ischemic stroke
Studies have implied that OPCs can proliferate and migrate along blood vessels to the ischemic core to play a protective role after ischemic stroke. 31 To evaluate whether oridonin increased the number of perivascular OPCs in the penumbra area by promoting its own proliferation, the fluorescent colocalization staining of PDGFRα (a marker for OPCs) and Ki67 (a marker for proliferating cells) was used for the observation of OPCs proliferation. Our results showed that compared with the tMCAO group, the number of OPCs expressing PDGFRα+/Ki67+ was significantly increased in the penumbra after oridonin administration at 3 and 7 days post-tMCAO/reperfusion (Figure 4(a) and (b)).
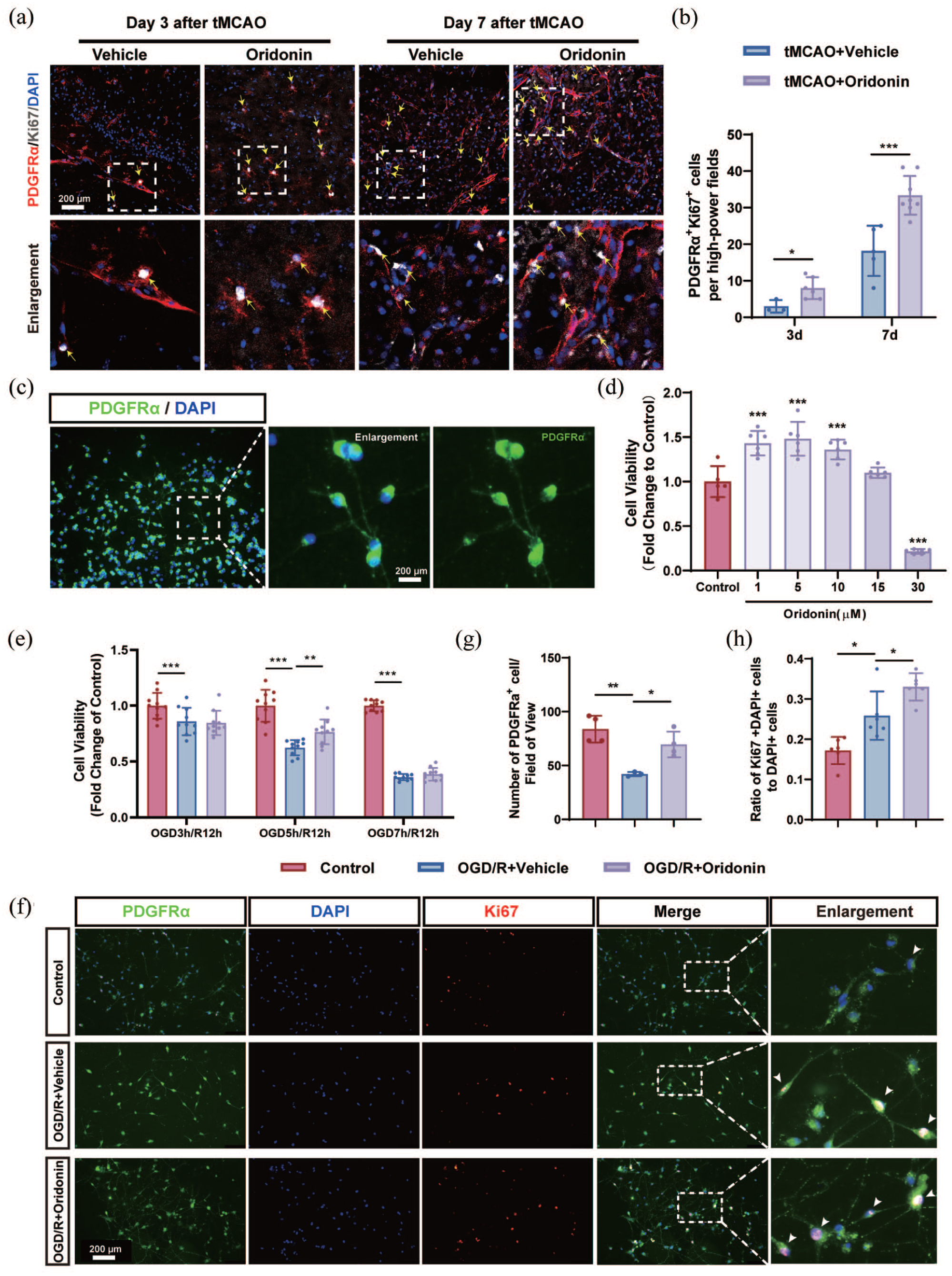
Oridonin enhanced oligodendrocyte precursor cells proliferation after ischemic stroke: (a) representative confocal images of PDGFRα (red), Ki67 (white), and DAPI (blue) immunostaining in ischemic penumbra at 3 and 7 days after ischemic stroke. Yellow arrows were used to indicate PDGFRα+/Ki67+ cells in the field. Scale bar = 200 μm, (b) quantitative analysis of PDGFRα+/Ki67+ OPCs of ischemic penumbra in each group, (c) isolation and purification of OPCs were visualized by immunofluorescence. Scale bar = 200 μm, (d) OPCs viability following various concentrations of oridonin (0, 1, 5, 10, 15, or 30 μM) was analyzed by MTT assay, (e) the effects of oridonin (5 μM) on the viability of OPCs after OGD 5 h/R 12 h was analyzed by MTT assay, (f) representative confocal images of PDGFRα (green), Ki67 (red), and DAPI (blue) immunostaining in primary OPCs after OGD/R. White arrows were used to indicate PDGFRα+/Ki67+ cells in the field. White boxes indicated areas that were enlarged in the fifth column of panel (f). Scale bar = 200 μm, (g, h) quantitative analysis of PDGFRα+ cell number and PDGFRα+/Ki67+ OPCs in each group. Three fields of view were captured and averaged per sample for all quantitative analyses. Statistical differences were analyzed by one-way ANOVA and Tukey’s multiple comparisons test or two-way ANOVA and Sidak’s multiple comparisons test. Data are presented as mean ± SD.
To further validate the in vitro effects of oridonin, primary OPCs were isolated and treated with varying concentrations of oridonin. As shown in Figure 4(c), phase-contrast microscopy revealed that cultured primary OPCs exhibited multipolar morphology and immunofluorescence staining for PDGFRα, a specific OPC marker, demonstrated a culture purity of 93.3%. MTT assay results showed that 5 μM oridonin significantly increased the OPC viability under normal culture conditions and OGD/R treatment (Figure 4(d) and (e)). Meanwhile, our results revealed that oridonin treatment significantly elevated the population of both PDGFRα+ OPCs and PDGFRα+/Ki67+ proliferating OPCs relative to the OGD/R group (Figure 4(f)–(h)).
Oridonin activated the Wnt7a/β-catenin pathway in OPCs to enhance endothelial angiogenic functions
Based on the above results, we found that oridonin directly promoted the proliferation of OPCs and increased the number of perivascular OPCs after ischemic stroke. However, it is still unclear whether and how OPC cells promote vascular endothelial cell angiogenesis under oridonin. It has been reported that hypoxic perivascular OPCs secreted a greater amount of pro-angiogenic factors, including VEGFa, Ang-1, and Wnt7a, to promote angiogenesis. 32 Moreover, numerous studies have shown that Wnt/β-catenin signaling is involved in angiogenesis in different pathophysiological processes. 33 Meanwhile, astrocyte-derived Wnt7a has been reported to promote endothelial cell proliferation and tight junction expression by activating β-catenin signaling after ischemic stroke. 34 To confirm whether oridonin elevated the pro-angiogenic factor levels, we detected the pro-angiogenic factor mRNA transcription changes in OPCs after OGD/R. RT-qPCR analysis revealed that oridonin-treated OPCs exhibited significantly elevated mRNA expression of VEGF-α, Ang-1, and Wnt7a compared to the OGD/R group (Figure 5(a)), with Wnt7a demonstrating the most pronounced change compared with the OGD/R group. Meanwhile, western blot analysis further confirmed that OGD/R treatment significantly reduced Wnt7a protein levels in OPCs, which was robustly reversed by oridonin treatment (Figure 5(b) and (c)). Meanwhile, we detected the concentration of Wnt7a by ELISA assay in the OPC conditioned medium (CM) under different treatments. The results showed that oridonin treatment markedly increased the Wnt7a levels in OPC CM compared with the OGD/R group (Figure 5(d)). Furthermore, bEnd.3 cells were incubated with CM collected from OPCs cultured with or without oridonin intervention (designated as Veh-OPC-CM and Ori-OPC-CM), so as to detect changes in the activity of the Wnt7a/β-catenin signaling pathway. We detected β-catenin nuclear translocation in bEnd.3 endothelial cells to verify Wnt/β-catenin pathway activation. The results showed that compared with the Control group, OGD/R treatment markedly inhibited the nuclear translocation of β-catenin. Treatment with Veh-OPC-CM failed to induce significant changes compared with the OGD/R group, whereas Ori-OPC-CM evidently promoted β-catenin nuclear translocation in bEnd.3 cells (Figure 5(e) and (f)). Therefore, we propose that the Wnt7a/β-catenin pathway plays a central role in mediating oridonin-induced angiogenesis.
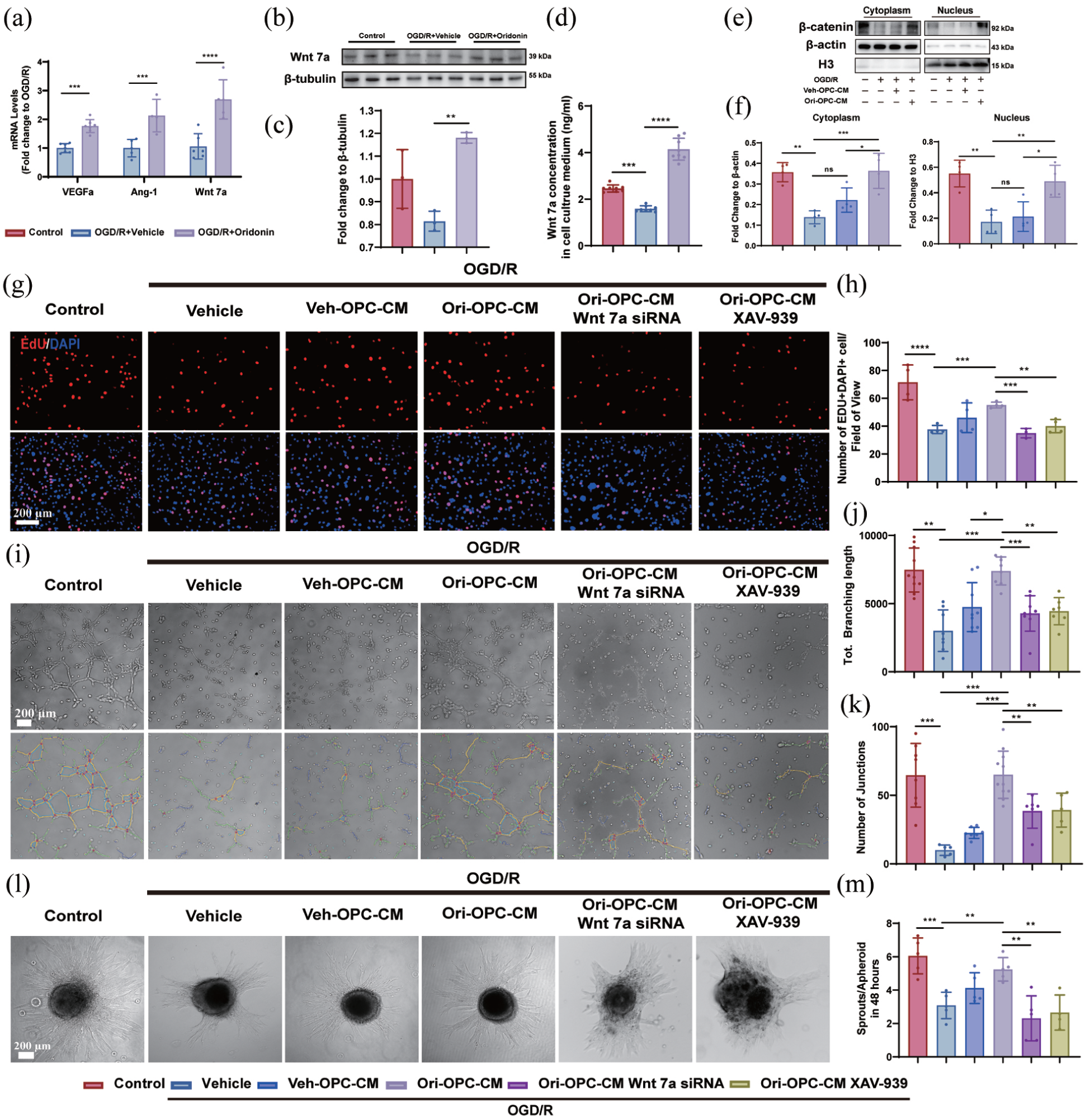
Oridonin activated the Wnt7a/β-catenin pathway in OPCs to enhance endothelial angiogenic functions: (a) the VEGFa, Ang-1, and Wnt7a mRNA levels changes were evaluated by RT-qPCR in each group, (b) the Wnt7a protein expression in OPCs after OGD/R with/without oridonin treatment, (c) the quantitative analysis of gray value was counted by fold change to β-tubulin, (d) the Wnt7a concentration in OPCs culture medium was assessed by ELISA assay, (e, f) nuclear–cytoplasmic fractionation was performed and analyzed to observe the nuclear translocation of β-catenin following different treatments, (g) EdU-labeled detection of bEnd.3 cell proliferation in different treatments was detected after OGD 6 h/R 24 h. Scale bar = 200 μm, (h) the number of EdU+DAPI+ cells per field of view was counted in each group, (i) the tube-forming assay and lumen integrity analysis were conducted to observe the tubular ability of endothelial cells. Scale bar = 200 μm. The total branching length (j) and the number of junctions (k) were quantitatively analyzed in each group, (l) representative images showed the spheroid sprouts of bEnd.3 cells. Scale bar = 200 μm, and (m) quantitative analysis of sprouts area/apheroid area of bEnd.3 cells in each group. Three fields of view were captured and averaged per sample for all quantitative analyses. Statistical differences were analyzed by one-way ANOVA and Tukey’s multiple comparisons test. Data are presented as mean ± SD.
To verify this conjecture, we knocked down Wnt7a in OPCs via siRNA (Figure S2(A) and (B)) or inhibited the Wnt7a/β-catenin pathway with XAV-939. We then treated bEnd.3 cells with Veh-OPC-CM or Ori-OPC-CM and monitored angiogenesis-related markers to evaluate angiogenic responses (Figure S2(C)). The EdU proliferation assay demonstrated that OPC-CM at a 12.5% dilution significantly enhanced the proliferation of bEnd.3 cells compared to the untreated vehicle group (Figure S2(D) and (E)). Our results showed that although Veh-OPC-CM promoted endothelial cell proliferation compared to OGD/R alone, Ori-OPC-CM elicited a markedly stronger proliferative response in bEnd.3 cells than Veh-OPC-CM. However, this endothelial pro-proliferation effect from Ori-OPC-CM was reversed by Wnt7a knockdown or XAV-939 treatment (Figure 5(g) and (h)). Meanwhile, the tube formation assay revealed that compared to the OGD/R group, Veh-OPC-CM failed to enhance the tubular network formation in bEnd.3 cells, whereas Ori-OPC-CM markedly increased both the total length and junction density of the luminal structures. Notably, the pro-angiogenic effect of Ori-OPC-CM was abolished by either Wnt7a knockdown or XAV-939 treatment, as observed in previous proliferation experiments (Figure 5(i)–(k)). The spheroid sprouting assay similarly showed that Ori-OPC-CM could significantly enhance the budding ability of bEnd.3 cells compared with the OGD/R group, which inhibited by Wnt7a or XAV-939 treatment (Figure 5(l) and (m)). These results indicated that OPC-CM treated with oridonin could promote the abilities of proliferation, sprouting, and tube formation in bEnd.3 cells after OGD/R, and this pro-angiogenic effect depended on OPC-derived Wnt7a acting on endothelial β-catenin.
Oridonin targeted PHD2 to attenuate ubiquitin-mediated HIF-1α degradation in OPCs
It has been reported that HIF-1α regulates the transcription of downstream genes, which modulates angiogenesis and OPC proliferation.35–37 Meanwhile, emerging evidence positions the Wnt/β-catenin pathway as a convergent node for hypoxia-induced responses and HIF-1α-mediated transcriptional regulation. 38 Therefore, we hypothesized that HIF-1α might be involved in the proliferation and the regulation of Wnt/β-catenin signaling in OPCs. And we next quantified HIF-1α protein expression in OPCs at 3, 6, 12, and 24 h after OGD/R. The results demonstrated a transient upregulation of HIF-1α, with peak expression occurring at 6 h, followed by a gradual decline to baseline levels by 12–24 h (Figure 6(a) and (b)). Interestingly, compared with the OGD/R group, oridonin treatment significantly increased HIF-1α protein levels from 3 to 24 h after OGD/R in OPCs (Figure 6(c)–(g)). Meanwhile, to further investigate whether oridonin directly regulated HIF-1α levels in endothelial cells, we assessed HIF-1α expression in bEnd.3 cells after OGD/R. Western blot results showed that there was no significant change in HIF-1α protein level after oridonin treatment compared with the OGD/R group (Figure 6(h)). These results reveal that oridonin can specifically promote the expression level of HIF-1α in OPCs.
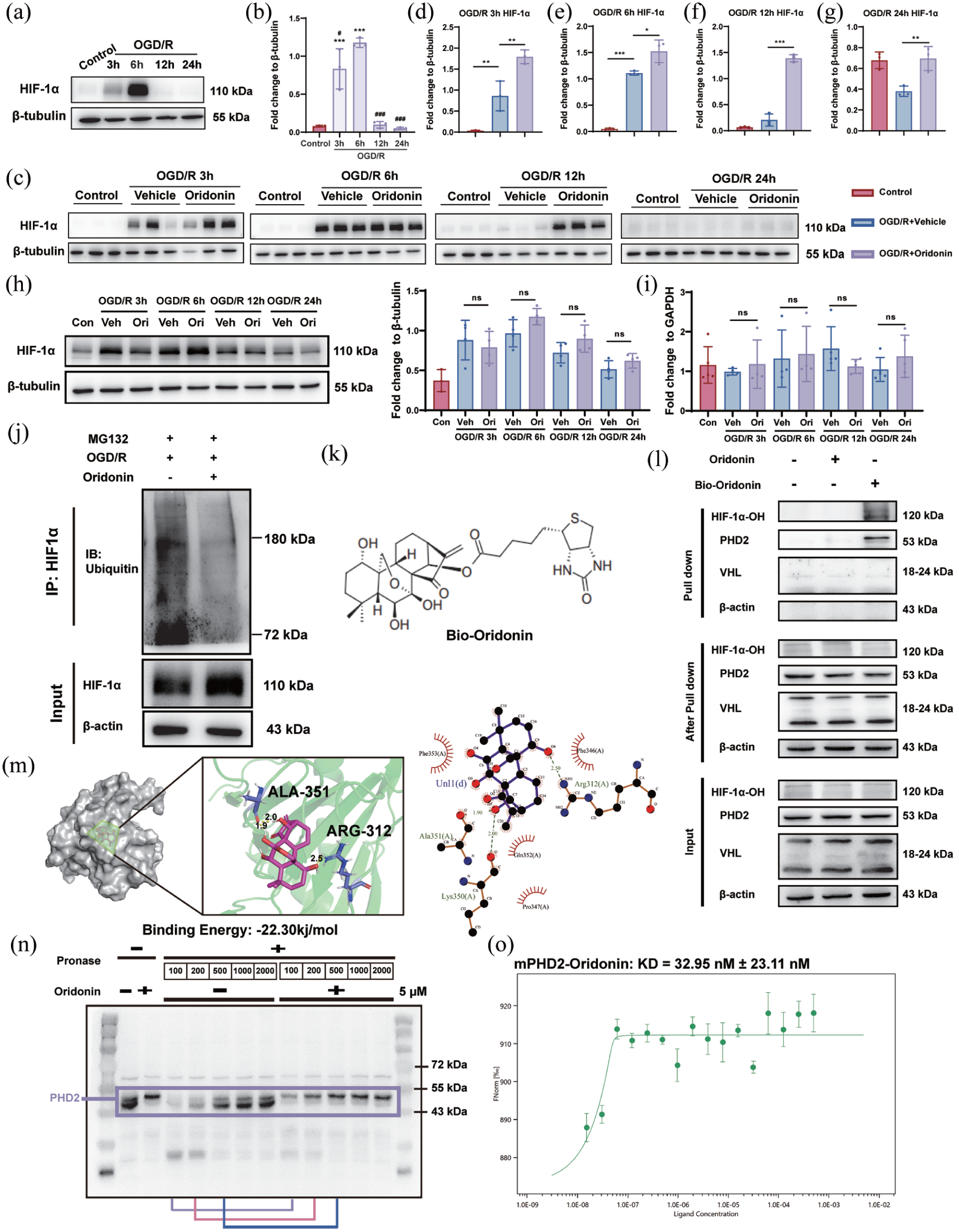
Oridonin targeted PHD2 to attenuate ubiquitin-mediated HIF-1α degradation in OPCs: (a, b) the dynamic changes of HIF-1α expression in OPCs after OGD 5 h and reoxygenation (3, 6, 12, and 24 h) were detected by western blotting, (c–g) HIF-1α expression changes were analyzed by western blotting in OPCs after treatment with 5 μM oridonin or not under different OGD/R time points, (h) Western blotting analysis of HIF-1α expression changes in bEnd.3 cells after normal culture and OGD/R 3–24 h with/without oridonin treatment, respectively, (i) the mRNA level change of HIF-1α in OPC under different treatments was quantified by RT-qPCR, (j) the effects of oridonin on ubiquitination of HIF-1α after OGD/R were analyzed by in vitro ubiquitination assays. HIF-1α was immunoprecipitated with anti-HIF-1α antibody and immunoblotted with anti-Ub antibody, (k) the molecular structure of Bio-oridonin, (l) cell lysates of OPC were incubated with Bio-oridonin (5 μM) for 2 h, which were then pulled down using streptavidin beads. The total proteins (input), bound proteins (pull down), and remaining proteins (after pull down) were immunoblotted as indicated, (m) the molecular docking 3D and 2D results of the optimal conformation of oridonin binding to PHD2 showed that the linked amino acids were ALA 351 and ARG 312, respectively, (n) lysates from OPC were incubated with or without oridonin (5 μM) for 1 h, and different concentrations of pronase were added for 20 min; PHD2 content was detected using western blot analysis, and (o) the binding of fluorescently labeled mPHD2 to oridonin was analyzed with MST. Statistical differences were analyzed by one-way ANOVA and Tukey’s multiple comparisons test. Data are presented as mean ± SD.
To explore the mechanisms involved in the up-regulative effects of oridonin on the HIF-1α protein expression in OPCs, we firstly detected the HIF-1α mRNA expression in OPCs from 3 to 24 h after OGD/R. RT-qPCR analysis showed that HIF-1α mRNA was not significantly changed after oridonin treatment compared with the OGD/R group, indicating that oridonin modulates HIF-1α protein degradation rather than its transcription or translation in OPCs (Figure 6(i)). HIF-1α is known to be extremely unstable and easily degraded by ubiquitination and HIF-1α degradation is mainly mediated by the ubiquitin-proteasome pathway. 39 Therefore, the impact of oridonin on HIF-1α ubiquitination complex protein levels in OPCs following OGD/R was detected. Firstly, we demonstrated that 10 µM MG132, a proteasome inhibitor, could significantly inhibit the immediate degradation of ubiquitinated HIF-1α by the proteasome (Figure S3(A) and (B)). CO-IP experiment results showed that oridonin markedly inhibited the ubiquitination degradation of HIF-1α (Figure 6(j)). These results demonstrate that oridonin specifically upregulates HIF-1α expression in OPCs by inhibiting its ubiquitination degradation, thereby enhancing OPC proliferation and Wnt7a secretion.
It is well established that α-ketoglutarate-dependent prolyl hydroxylase (PHD) catalyzes the hydroxylation of proline residues within the oxygen-dependent degradation domains of HIF-1α under normoxic conditions. This process is primarily mediated by PHD2, whose enzymatic product is recognized by the VHL ubiquitin ligase complex, thereby triggering HIF-1α ubiquitination and subsequent degradation.40,41 Since oridonin significantly inhibited HIF-1α ubiquitination and degradation, we hypothesized a direct interaction between oridonin and PHD2, HIF-1α-OH, or VHL. To identify oridonin’s potential protein targets, we biotinylated oridonin (Bio-oridonin) and performed streptavidin bead-based pull-down assays (Figure 6(k)). The result showed that Bio-oridonin could bind to PHD2 and HIF-1α-OH, but not VHL, compared with unlabeled oridonin (Figure 6(l)). Furthermore, we performed molecular docking experiments to determine whether PHD2 binds directly to oridonin. The results indicated a potential interaction between PHD2 and oridonin, which is characterized by a binding energy of −22.30 kJ/mol (Figure 6(m)). In order to test the direct bond between PHD2 and oridonin, DARTS were conducted to confirm this direct interaction between oridonin and PHD2. Following exposure to 5 μM oridonin, PHD2 levels remained elevated compared to the untreated group across varying pronase concentration gradients, suggesting that oridonin binds to PHD2 and protects it from enzymatic degradation (Figure 6(n)). Meanwhile, we examined the binding capacity of oridonin to the other two members of the PHD family using DARTS assays to clarify whether oridonin interacts with PHD2 in a specific manner. The results revealed that oridonin could not attenuate the gradient protease-mediated degradation of PHD1 and PHD3, indicating that oridonin binds specifically to PHD2 (Figure S4(A) and (B)). Furthermore, we validated the direct binding of oridonin and PHD2 by MST, and the results definitely confirmed that the binding affinity between oridonin and PHD2 was 32.95 ± 23.11 nmol/L (Figure 6(o)). Therefore, these results demonstrate that oridonin directly targets PHD2 to inhibit HIF-1α ubiquitination degradation.
PHD2 overexpression reversed oridonin-induced high Wnt7a expression in OPCs and Wnt/β-catenin activation in vascular endothelial cells
To determine whether PHD2 is required for the pro-angiogenic actions of oridonin in ischemic stroke, we overexpressed PHD2 in OPCs. We first verified the overexpression efficiency of PHD2 (Figure 7(a) and (b)). Under PHD2 overexpression, we examined the Wnt7a expression in OPCs after OGD/R and oridonin treatment, and the results showed that PHD2 overexpression significantly reversed the upregulation of Wnt7a expression induced by oridonin in OPCs (Figure 7(c) and (d)). Furthermore, we collected conditioned medium (CM) from OPCs exposed to OGD/R and oridonin with or without PHD2 overexpression, and treated OGD/R-injured bEnd.3 cells with the collected CM. The results revealed that compared with CM from OPCs subjected to oridonin and OGD/R alone, PHD2 overexpression markedly reversed the stimulatory effect of Ori-OPC-CM on β-catenin nuclear translocation in bEnd3 cells (Figure 7(e)–(g)). Thus, our results indicate that oridonin exerts pro-angiogenic effects by activating Wnt/β-catenin signaling between OPCs and endothelial cells in a PHD2-dependent manner.
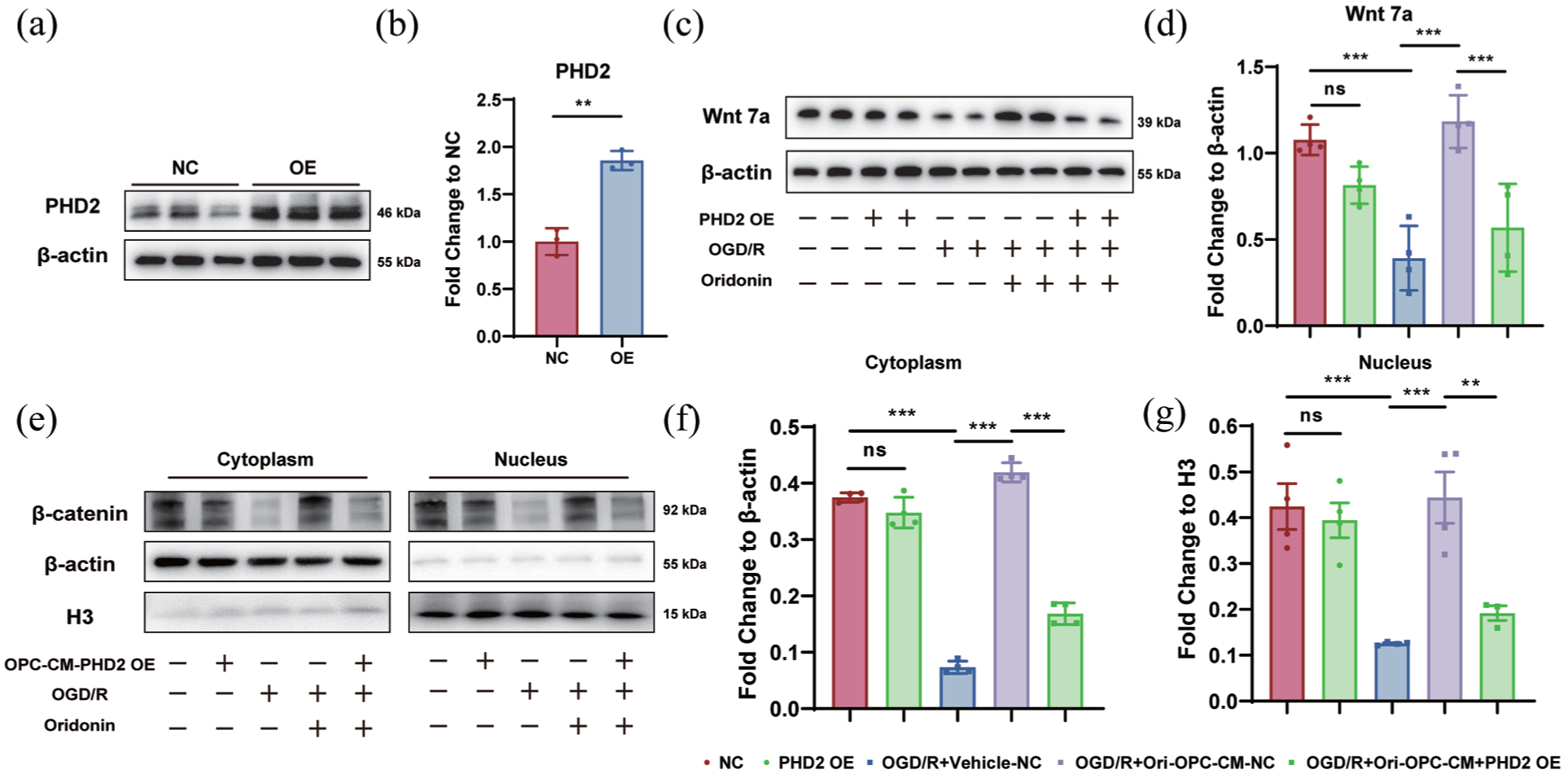
PHD2 overexpression reversed oridonin-induced high Wnt7a expression in OPCs and Wnt/β-catenin activation in vascular endothelial cells: (a, b) the PHD2 overexpression was conducted and verified the overexpression efficiency by WB, (c, d) the Wnt7a protein expression in OPCs after OGD/R and oridonin treatment with or without PHD2 overexpression was detected by WB and the quantitative analysis of gray value was counted by fold change to β-actin, (e) nuclear–cytoplasmic fractionation was performed and examined by WB to observe the nuclear translocation of β-catenin following different treatments, (f, g) quantitative analysis of β-catenin gray value was counted by fold change to β-actin or Histone H3. Statistical differences were analyzed by one-way ANOVA and Tukey’s multiple comparisons test. Data are presented as mean ± SD.
Discussion
Angiogenesis serves as a critical protective mechanism that facilitates neural regeneration and functional recovery in the pathophysiological progression of stroke. 29 Oridonin is a bioactive substance extracted from the traditional Chinese herb Rabdosia rubescens and belongs to the class of tetracyclic diterpenoid, and as a small-molecule compound with a molecular weight of 364 Da, oridonin has been proven to freely cross the blood–brain barrier (BBB) and exert neuroprotective effects in multiple neurological diseases, including traumatic brain injury, Alzheimer’s disease and depression.42–44 This endows oridonin with promising clinical translational potential as a therapeutic agent for ischemic stroke. Our previous study found that oridonin improved BBB integrity and increased the expression of tight junction proteins via inhibiting endothelial oxidative stress injury, consequently attenuating peripheral inflammatory cell infiltration and neuroinflammation during the acute phase. In the present study, we innovatively revealed that oridonin prevents HIF-1α from undergoing ubiquitination and subsequent degradation in OPCs by targeting PHD2, and this stabilization of HIF-1α stimulates OPCs proliferation and upregulates the Wnt7a/β-catenin signaling pathway, ultimately enhancing endothelial cell-induced angiogenesis.
Angiogenesis, a vital mechanism for neuroprotection and tissue repair post-ischemic stroke, operates with spatiotemporal specificity.24,45 Occurring in the penumbra area as early as 12 h after stroke, angiogenesis contributes to the reformation of the neural vascular network and restoration of blood flow and neuronal metabolism in tissue at risk. 46 However, this protective revascularization process is temporary. Our study found that oridonin increased the vessel density and number of junctions at 7 and 14 days after ischemic stroke. Furthermore, we were surprised to find that these beneficial effects of oridonin did not depend on its promotion of endothelial cell proliferation, sprouting, and tube formation, which inspired us to explore whether oridonin might act on other cell types.
Numerous studies have reported that pluripotent glial cells, particularly OPCs, relocate to perivascular zones post-stroke, actively participating in revascularization and tissue remodeling. 47 Our study identified two distinct subpopulations of OPCs: perivascular OPCs, which extensively envelop microvessels via their fine cytoplasmic processes, and parenchymal OPCs, which reside distal to vascular structures, aligning with previous reports.32,48 Meanwhile, our data demonstrated a time-dependent increase in OPC density during the 14-day post-tMCAO period, paralleled by a progressive augmentation of vascular network density. The correlation analysis showed that the number of perivascular OPCs was positively correlated with vessel density. Importantly, we found that oridonin could significantly increase the number of perivascular OPCs in the ischemic penumbra area at 3, 7, and 14 days after tMCAO. Furthermore, we unexpectedly identified a novel pharmacological property of oridonin, demonstrating its significant capacity to enhance OPCs proliferation in vivo and in vitro. These findings suggest that oridonin’s pro-angiogenic effects may be mediated through its stimulation of OPC proliferation. This finding is consistent with the model of periodontitis in which oridonin promotes LPS-induced proliferation of human periodontal ligament stem cells. 49 These findings both demonstrate the proliferative activity of oridonin on pluripotent stem cells. Research evidence indicates that oligodendrocyte precursor cell (OPC) migration from blood vessels and subsequent detachment are essential for proper myelination.12,50 Our experimental observations demonstrate that Oridonin enhances the proliferation of perivascular OPCs. However, its potential role in promoting remyelination in white matter areas requires further investigation.
Recent studies have demonstrated that OPCs residing in perivascular hypoxic niches exhibit enhanced secretion of pro-angiogenic factors, thereby actively contributing to angiogenesis. 32 Our study found that oridonin significantly increased the mRNA and protein expression levels of pro-angiogenic factor Wnt7a in OPCs after OGD/R treatment, accompanied by a similar increase in Wnt7a secretion in OPCs medium. The increase in Wnt7a secretion from OPCs was maybe responsible for the pro-angiogenic effect of oridonin. Therefore, we further found that OPC-CM treated with oridonin could significantly improve proliferation, sprouting, and tube formation of bEnd.3 after OGD/R, but these effects were reversed by Wnt7a knockdown in OPCs or XAV-939 treatment. These data suggested that oridonin mediates crosstalk between OPCs and endothelial cells through enhancing Wnt7a/β-catenin signaling, thus promoting angiogenesis. Consistent with previous reports, studies have shown that interactions between OPCs and endothelial cells orchestrate vascular development in neonatal white matter via Wnt signaling, with this axis further implicated in mitigating hypoxic-ischemic injury. 11
OPC-encoded HIF protein function is an essential regulator of angiogenesis, and Wnt7a is the direct HIF-inducible target gene.37,51,52 Meanwhile, established knowledge has confirmed that HIF-1α is closely involved in cell proliferation through its transcriptional activity.53,54 Here, we hypothesized that essential functions of HIF-1α were involved in oridonin’s role in OPCs proliferation and Wnt7a secretion. Our data demonstrate a biphasic response of HIF-1α protein levels during reoxygenation following OGD, with peak expression observed at 6 h post-reoxygenation. Meanwhile, oridonin significantly increased the protein level of HIF-1α in OPCs at 3, 6, and 12 h after OGD/R. However, quantitative RT-PCR analysis revealed that oridonin had no significant effect on HIF-1α mRNA transcription levels, suggesting its potential role in inhibiting HIF-1α protein degradation. PHDs are responsible for regulating HIF-1α activity in an oxygen-dependent manner.55–57 It is well known that HIF-1α is easily recognized by VHL for ubiquitination and degradation after hydroxylation by PHD2.58,59 Therefore, we further investigated and demonstrated that oridonin inhibited the OGD/R-induced ubiquitin-proteasome degradation of HIF-1α protein, and interacted with the hydroxylation of HIF-1α and PHD2 by a biotinylation assay. Furthermore, the DARTS and MST assay confirmed that oridonin inhibits the ubiquitination degradation of HIF-1α targeting PHD2, and thereby promotes OPC proliferation to exert the pro-angiogenesis effects after ischemic stroke. Although there have been studies reported that oridonin exerted its anti-angiogenesis activity through significantly decreasing HIF-1α expression in breast cancer and colon cancer,60,61 this may be due to the different regulatory effects of oridonin on pluripotent stem cells such as OPCs and cancer cells, which are completely different cell types. Meanwhile, accumulating evidence demonstrates that pharmacological suppression of HIF1α triggers OPC differentiation and HIF1α is indispensable for preserving the pluripotent stem cell properties of oligodendrocyte precursor cells (OPCs), a process associated with enhanced Wnt7a secretion,62,63 which is consistent with our study that oridonin stabilizes HIF1α by inhibiting its ubiquitination in OPCs, thereby augmenting Wnt7a production and subsequent endothelial cell crosstalk to facilitate angiogenesis. Our study highlights the key role of oridonin in inhibiting HIF-1α protein ubiquitination degradation by targeting PHD2.
In sum, our results provide strong evidence that oridonin promotes post-stroke angiogenesis by targeting PHD2 to stabilize HIF-1α, driving OPC proliferation and Wnt7a/β-catenin pathway activation, which orchestrates endothelial-mediated angiogenesis. Our data may provide a natural candidate compound for accelerated angiogenesis and a promising treatment agent for ischemic stroke. Our study has demonstrated that oridonin increased OPCs proliferation and angiogenesis, and this reparative response is significantly impaired in aged individuals—the primary demographic affected by stroke. Given the well-documented age-related decline in OPC regenerative capacity and its detrimental effects on white matter repair, this rationale underscores the therapeutic potential of oridonin in targeting the OPC proliferation pathway, particularly in clinically relevant aging populations where endogenous repair fails.
Supplemental Material
sj-docx-1-jcb-10.1177_0271678X261462715 – Supplemental material for Oridonin inhibits HIF-1α ubiquitination by targeting PHD2 in oligodendrocyte precursor cells to promote angiogenesis after ischemic stroke
Supplemental material, sj-docx-1-jcb-10.1177_0271678X261462715 for Oridonin inhibits HIF-1α ubiquitination by targeting PHD2 in oligodendrocyte precursor cells to promote angiogenesis after ischemic stroke by Wei Guo, Shu-Qi Cheng, Yan-Yan Liu, Han-Lian Xiao, Xue-Wei Ren, Lei Li, Juan Ji, Xi-Yue Zhang and Xiu-Lan Sun in Journal of Cerebral Blood Flow & Metabolism
Supplemental Material
sj-docx-2-jcb-10.1177_0271678X261462715 – Supplemental material for Oridonin inhibits HIF-1α ubiquitination by targeting PHD2 in oligodendrocyte precursor cells to promote angiogenesis after ischemic stroke
Supplemental material, sj-docx-2-jcb-10.1177_0271678X261462715 for Oridonin inhibits HIF-1α ubiquitination by targeting PHD2 in oligodendrocyte precursor cells to promote angiogenesis after ischemic stroke by Wei Guo, Shu-Qi Cheng, Yan-Yan Liu, Han-Lian Xiao, Xue-Wei Ren, Lei Li, Juan Ji, Xi-Yue Zhang and Xiu-Lan Sun in Journal of Cerebral Blood Flow & Metabolism
Footnotes
Author contributions
Xiu-Lan Sun conceived the project and wrote the manuscript. Wei Guo and Shu-Qi Cheng assisted with study design, performed in vivo and in vitro experiments, and contributed to data analysis. Yan-Yan Liu and Han-Lian Xiao conducted supplementary animal and molecular biology experiments during the manuscript revision. Xue-Wei Ren and Lei Li synthesized and purified the PHD2 protein and performed binding assays. Juan Ji and Xi-Yue Zhang critically revised the manuscript. All authors have approved the final manuscript and have agreed to be accountable for all aspects of the work.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National Natural Science Foundation of China (nos. 82373848, 81973301, and 82304479) and the National Key Research and Development Program of China (no. 2021ZD0202901).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.