
Abstract
Several studies have shown the ability of transcription factor 12 (TCF12) to promote tumor malignant progression, but its function in glioma cells has not been fully elucidated. In this study, we analyzed the data from TCGA by bioinformatics and found that in glioma tissue, TCF12 was conspicuously highly expressed while miR-218-5p was significantly low-expressed. The downregulation of miR-218-5p was correlated with adverse prognosis in patients with glioma. miR-218-5p was found to be negatively associated with TCF12 by Pearson correlation analysis, and dual luciferase assay was employed to verify that miR-218-5p and TCF12 had a targeting relationship. qRT-PCR and Western blot assays were used to verify that the expression of TCF12 was regulated by its upstream regulator miR-218-5p. Moreover, cell experiments validated that overexpressed TCF12 could promote the proliferation, migration, and invasion of glioma cells and inhibit their apoptosis, whereas overexpressing miR-218-5p at the same time could reverse this phenomenon. Our study demonstrates the regulatory mechanism of the miR-218-5p/TCF12 axis in gliomas, which lays a foundation for searching for new therapeutic approaches for glioma.
Introduction
Glioma is the most common primary intracranial malignancy in adults, 1 and can be classified as astrocytomas, oligodendrogliomas, and ependymomas, based primarily on microscopic observation of tumor tissue sections. 2 Glioma is characterized by high malignancy, rapid proliferation, easy recurrence, and poor prognosis, and is considered one of the most intractable tumors. 3 The main treatment for glioma is surgery combined with chemotherapy or radiotherapy, 4 but patient prognosis remains poor. 5 Therefore, understanding the mechanism that triggers the regulation of glioma cells is more conducive to finding new therapeutic strategies.
Transcription factor 12 (TCF12) protein is one of the basic helix-loop-helix (bHLH) E proteins that plays an important role in the development and differentiation of neurons and can be functioned as a mediator for the osteogenic differentiation of bone marrow mesenchymal stem cells both in vivo and in vitro.6,7 Studies have shown that TCF12 also plays a key regulatory role in some cancers. For example, TCF12 is highly expressed in ovarian cancer tissue, while downregulating the expression of TCF12 can inhibit the proliferation and metastasis of ovarian cancer cells and promote their apoptosis. 8 Overexpression of TCF12 in liver cancer cells can enhance cell proliferation and metastases, while deleting TCF12 can cause the opposite effect. 9 Overexpression of TCF12 inhibits the proliferation, migration, and invasion of prostate cancer cells in vitro as well as tumor growth in vivo. 10 However, there is little research into the role of TCF12 in gliomas.
MicroRNAs (miRNAs) typically control genes involved in cellular processes such as inflammation, cell cycle regulation, stress response, differentiation, apoptosis, and migration. 11 Since the discovery of miRNAs, they have become an important research topic, and with a deeper understanding of their role in cancer, miRNAs have become popular tools and targets for novel therapies. 12 A study has shown that miRNAs can be tumor suppressors or oncogenes whose dysregulated expression is associated with various cancers. 13 As one of the members of the miR-218 family, miR-218-5p is authenticated to be dysregulated in varying cancers and affects the malignant progression of cancers. In gastric cancer, miR-218 hinders cell proliferation, induces cell cycle blockage at G1 phase, and hampers tumor progression and metastasis. 14 In non-small cell lung cancer, miR-218-5p represses cell proliferation and migration via epidermal growth factor receptor (EGFR). 15 miR-218-5p restrains cell growth and metastasis of cervical cancer through the LYN/NF-кB signaling pathway. 16 However, its effect on glioma cells needs further study.
This study demonstrates that TCF12 was significantly highly expressed and miR-218-5p was significantly low-expressed in glioma tissue by bioinformatics analysis, and there was a significant negative correlation between them. Experiments were performed to verify the effect of TCF12 on the biological function of glioma cells and to discover the regulatory mechanism of the miR-218-5p/TCF12 axis in glioma cells. The results further elucidate the relationship between the miR-218-5p/TCF12 axis and glioma, providing new approaches for the diagnosis and treatment of glioma.
Methods
Bioinformatics analysis
The mature miRNA (normal: n = 5, tumor: n = 530) and mRNA (normal: n = 5, tumor: n = 698) expression data of gliomas were downloaded from the TCGA database (https://portal.gdc.cancer.gov/) on January 1, 2020. Based on the downloaded mRNA expression data, t test was adopted to detect the differential expression of TCF12 in normal tissue and glioma tissue with normal tissue as control. The R package edgeR was used to perform differential analysis of miRNAs (|logFC|>2, padj < 0.01), with the normal tissue (n = 5) in the miRNA gene expression matrix as a control, thereby obtaining differentially expressed miRNAs (DEmiRNAs), the miRNA of interest among which was then analyzed via survival analysis. The miRDB (http://mirdb.org), starBase (http://starbase.sysu.edu.cn/), and mirDIP (http://ophid.utoronto.ca/mirDIP/index.jsp) databases were employed to detect the upstream miRNAs regulating TCF12, which were intersected with the downregulated DEmiRNAs. Next, Pearson correlation analysis was conducted on the obtained miRNAs and TCF12, and the miRNA that was the most corelative with TCF12 was selected for the study.
Cell culture
The normal human glial cell line HEB (MZ-0831) and glioma cell lines U251 (MZ-0186), U87 (MZ-2007), T98-G (ATCC CRL-1690), and A172 (MZ-0012) were purchased from Ningbo Mingzhou Biotechnology. Cells were cultivated in Dulbecco modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin at 37°C with 5% CO2.
Cell transfection
The transfected cells were inoculated into 6-well plates (5×105 cells/well). Cell transfection started when cell confluence reached 60%–70%. Briefly, 50 nM miR-218-5p mimic, 50 nM negative control mimic (NC-mimic), 1 µg pcDNA3.1-TCF12 plasmid (oe-TCF12), or 1 µg empty pcDNA3.1 plasmid vector (oe-NC) was transfected into U251 cell line with Lipofectamine 2000 (Invitrogen, Carlsbad, CA) following the manufacturer’s instructions. All gene fragments and plasmids were accessed from RiboBio. All transfected cells were prepared at 37°C with 5% CO2 for 2 days.
Quantitative real-time polymerase chain reaction assay
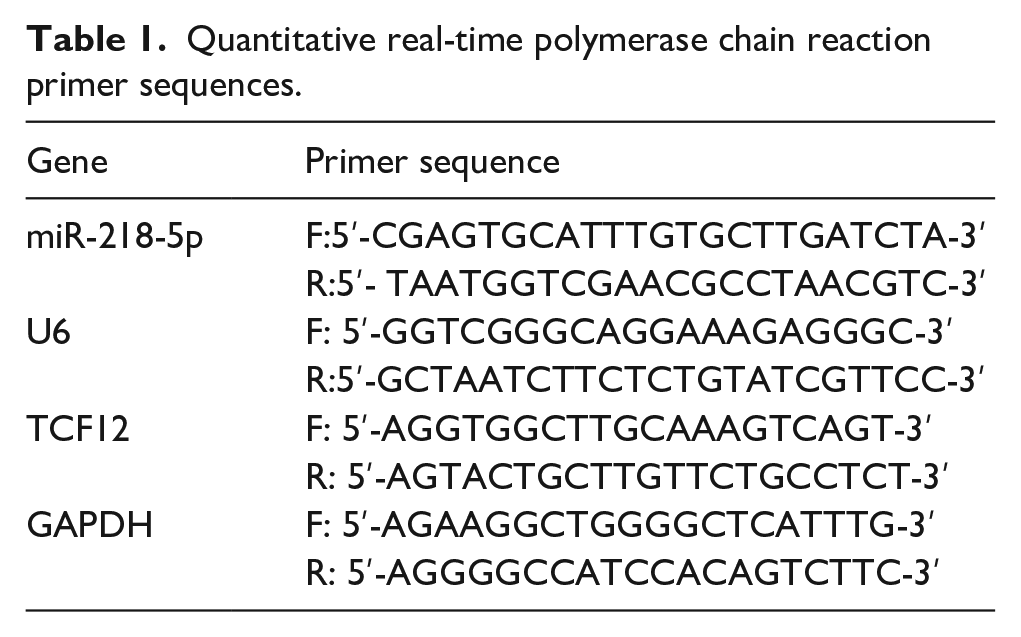
Total RNA was extracted from transfected cells by TRIzol reagent (Life Technologies). miRNAs were reversely transcribed to cDNAs with miScript IIRT Kit (Qiagen) according to the instructions. The mRNAs were transcribed to cDNAs with PrimeScript RT Master Mix (Takara [Dalian]). The expression of miRNA was detected by miScript SYBR Green PCR Kit (Qiagen) and the expression of mRNA was detected by SYBR® Premix Ex Taq TM II (Takara Bio Inc.). Quantitative real-time polymerase chain reaction (qRT-PCR) was run on Applied Biosystems® 7500 real-time PCR systems (Thermo Fisher Scientific) to detect expression levels of miR-218-5p and TCF12 mRNA. TCF12 was internally regulated by GAPDH and miR-218-5p by U6. The 2–ΔΔCt method was used to analyze the relative expression. The experiment was repeated 3 times. The primer sequences are listed in Table 1.
Quantitative real-time polymerase chain reaction primer sequences.
Western blot
The transfected cells were rinsed with phosphate-buffered saline and lysed with RIPA lysis buffer (Beyotime). The concentration of the total proteins was measured with the BCA protein assay kit (Pierce). Cell lysates were electrophoresed with sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes, which were incubated overnight at 4°C with the following primary antibodies: rabbit anti-TCF12 (Abcam) and rabbit anti-GAPDH (Abcam). Then the membranes were incubated for 2 hours at room temperature with secondary antibody goat anti-rabbit immunoglobulin G H&L (HRP) (Abcam). The protein expression was detected with GAPDH as an internal control by an enhanced chemiluminescence kit (GE Healthcare).
CCK-8
To assess cell activity, transfected cells were inoculated into 96-well plates at a density of 5×103 cells per well. Cell proliferation was measured 1, 2, and 3 days after incubation by CCK-8 assay (Dojindo), and the absorbance was measured at 450 nm. The experiment was repeated three times.
Cell migration and invasion assay
For the cell migration assay, transfected cells were inoculated into 6-well plates and cultured to 90% confluence. An artificial wound was created with a 2-mm cell scraper. Cells were cultured in serum-free medium and cell migration was observed with a light microscope at 0 and 48 hours, respectively. The experiment was repeated three times. The wound healing rate (%) = (wound width at 0 hours – wound width at 48 hours)/wound width at 0 hours × 100%.
For the cell invasion assay, a layer of Matrix matrigel (BD Biosciences) was added onto Transwell chambers. Cell suspension containing 5×105 cells was added to the upper chamber and DMEM containing 10% FBS was added to the lower chamber. After incubation at 5% CO2, 37 °C for 24 hours, cells that did not pass through the membrane were removed with cotton swabs. The remaining cells were stained with gentian violet (Sigma-Aldrich; Merck-KGaA) for 10 minutes at room temperature, rinsed with water, and finally counted under an optical microscope. The experiment was repeated three times.
Apoptosis assay
The transfected cells were incubated for 48 hours and digested by trypsin. The cells were then resuspended in binding buffer. Annexin V-FITC and propidium iodide were then added. Apoptosis was detected with FACS Calibur flow cytometry (Becton Dickinson). The results were analyzed later. Each experiment was repeated three times.
Dual luciferase assay
Luciferase vectors psiCHECK-TCF12 (Sangon Co.) fused with wild-type (TCF12-WT) and mutant TCF12 3′UTR (TCF12-MUT) were constructed. miR-218-5p-mimic/NC-mimic and psiCHECK-TCF12 WT/MUT were co-transfected into U251 cells according to the operation instructions of Lipofectamine 2000 reagent (Invitrogen). Luciferase activity was measured by the Dual-Luciferase Reporter Assay System (Promega). The experiment was repeated three times.
Statistical analysis
All the data were analyzed with GraphPad Prism 6.0 and each experiment was repeated 3 times. The results were presented as mean ± standard deviation. The comparison between two groups was analyzed by Student t test, while the comparison among multiple groups was analyzed by one-way analysis of variance. P<0.05 indicated that the difference was statistically significant.
Results
TCF12 is significantly highly expressed in glioma cells
According to several studies, TCF12 plays a vital role in cancer progression. This study investigated TCF12 and clarified its biological role in the progression of glioma. Glioma data in the TCGA database showed that the expression of TCF12 was significantly upregulated in glioma tissue (Figure 1 [A]). qRT-PCR was used to detect the expression of TCF12 in glioma cell lines and a significant increase was found in the expression of TCF12 mRNA in cancer cells relative to that in normal cells (Figure 1 [B]). The above results showed that TCF12 was prominently overexpressed in glioma cells. In order to better explore the mechanism of TCF12, the U251 cell line with the highest expression in TCF12 was chosen for follow-up experiments.
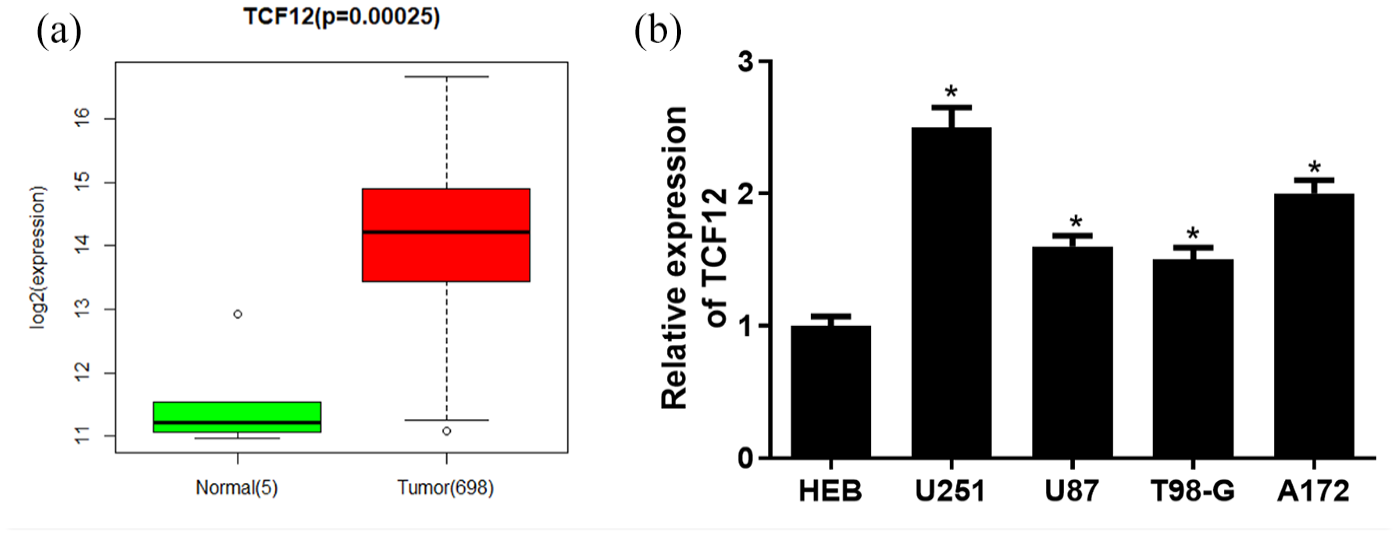
Significant high expression of TCF12 in glioma cells. (
Overexpression of TCF12 promotes the proliferation, migration, and invasion of glioma cells and inhibits their apoptosis
To study the effect of TCF12 on the biological function of glioma cells, we upregulated the expression of TCF12 in the U251 cell line. The transfection efficacy of TCF12 in glioma cell lines was assessed by qRT-PCR and Western blot, the results of which displayed that TCF12 mRNA and protein expression levels increased notably in the oe-TCF12 group (Figure 2 [A]), and the cells successfully overexpressing TCF12 were used in subsequent experiments. The results of the CCK-8 showed that overexpression of TCF12 promoted the proliferation of glioma cells (Figure 2 [B]). Transwell and wound healing assays showed that overexpression of TCF12 promoted the ability of glioma cells to invasion and migration (Figure 2 [C] and [D]). Flow cytometry showed that the apoptosis capacity of glioma cells decreased after overexpressing TCF12 (Figure 2 [E]). The above results suggested that overexpression of TCF12 hastened the proliferation, migration, and invasion of glioma cells and hindered their apoptosis.
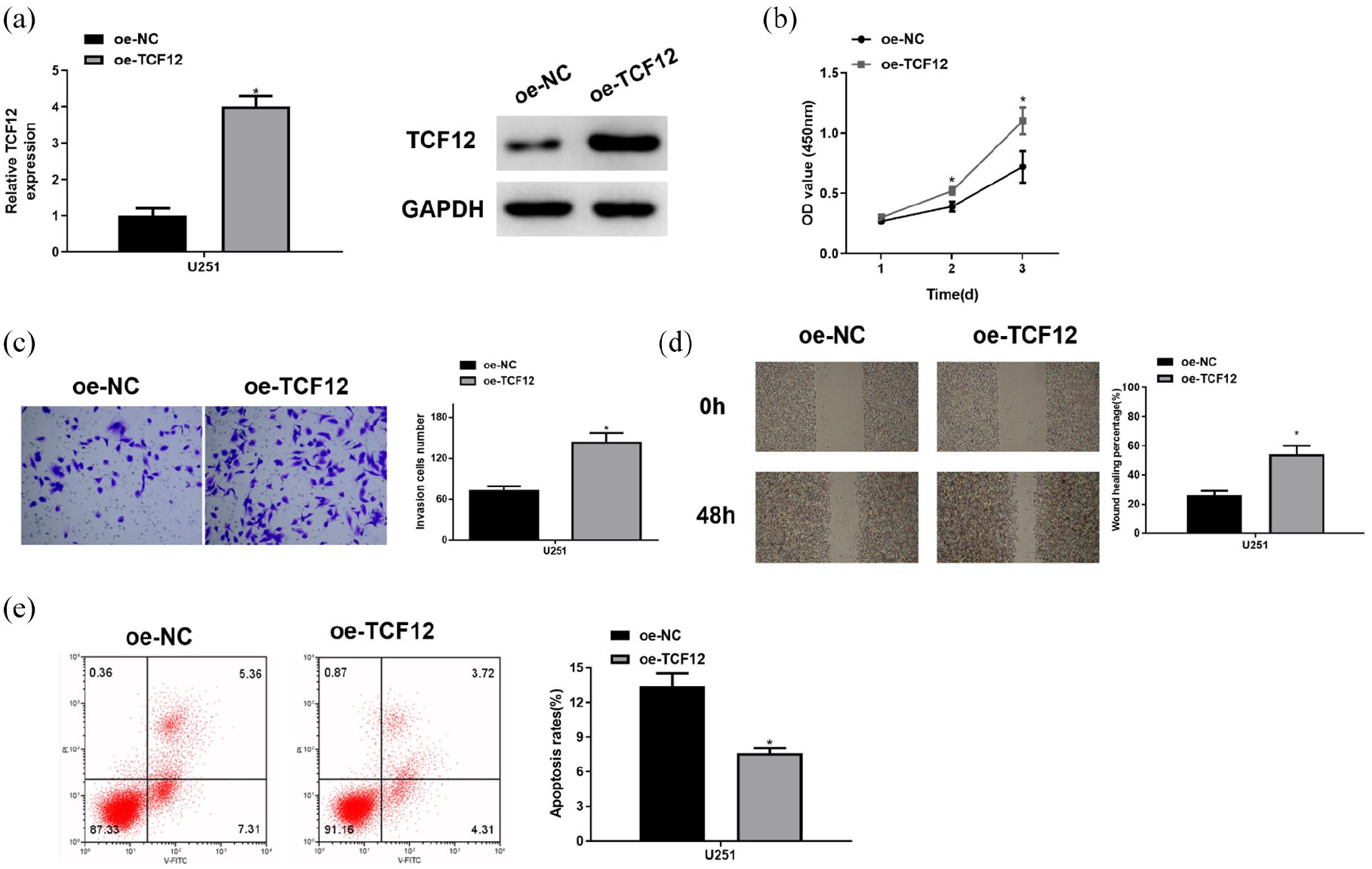
Overexpression of TCF12 promotes the proliferation, migration, and invasion of glioma cells and inhibits their apoptosis. (
miR-218-5p is low-expressed in glioma cells and is significantly negatively associated with TCF12
The above results elucidated that TCF12 was significantly highly expressed in glioma. In order to investigate the upstream regulatory mechanism of TCF12 in glioma, R package “edgeR” was used to analyze the DEmiRNAs from TCGA database by bioinformatics analysis, and 78 DEmiRNAs were obtained. Among them, 43 were upregulated and 35 were downregulated (Figure 3 [A]). Upstream miRNAs of TCF12 were predicted with starBase, mirDIP, and miRDB bioinformatics databases, and the results were intersected with 35 downregulated DEmiRNAs, obtaining 3 miRNAs (miR-139-5p, miR-7-5p, and miR-218-5p), which bound to TCF12 (Figure 3 [B]). Pearson correlation analysis revealed that TCF12 had the highest correlation coefficient with miR-218-5p (–0.32), indicating a remarkable inverse correlation (Figure 3 [C]). miR-218-5p was significantly downregulated in glioma tissue in TCGA (Figure 3 [D]), and survival analysis showed that patients with glioma with high expression of miR-218-5p could live longer than those with low expression (Figure 3 [E]). Then, the expression of miR-218-5p in glioma cell lines and a normal cell line was detected with qRT-PCR, the result of which showed that the expression in glioma cell lines was significantly downregulated (Figure 3 [F]). The above results elucidated that TCF12 was significantly negatively correlated with miR-218-5p and miR-218-5p was significantly decreased in glioma cells.
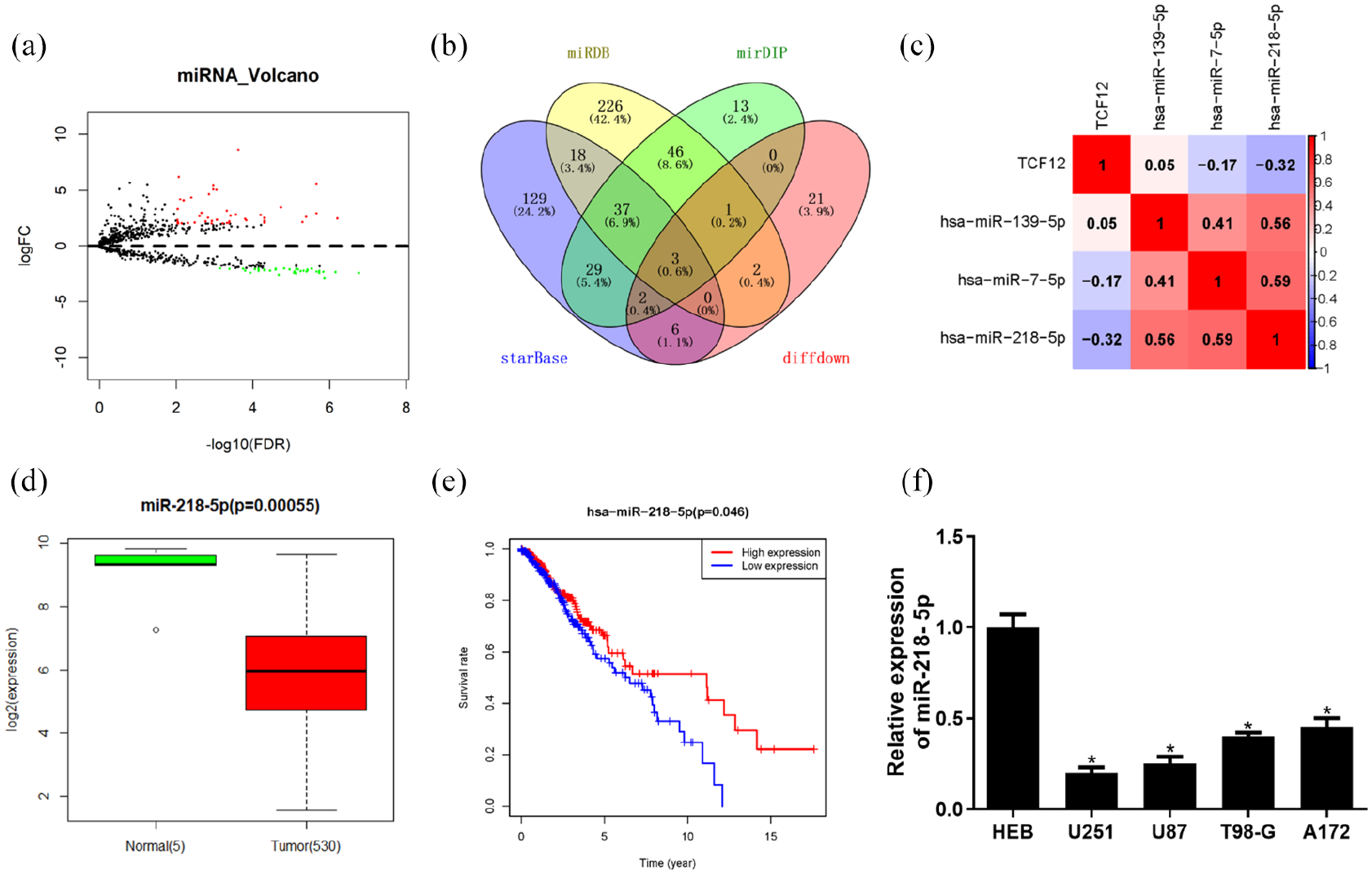
miR-218-5p is downregulated in glioma cells and significantly negatively correlated with TCF12. (
miR-218-5p targets TCF12 and inhibits its expression in glioma cells
The previous results showed that miR-218-5p was significantly negatively correlated with TCF12 in glioma cells, and then whether miR-218-5p had a targeting relationship with TCF12 was verified. In the starBase database, the targeted binding sequence of miR-218-5p on TCF12 3′UTR was predicted (Figure 4 [A]). The dual luciferase assay was employed for further verification. The result revealed that overexpression of miR-218-5p inhibited the luciferase activity of TCF12-WT but had no effect on that of TCF12-MUT, demonstrating that miR-218-5p could bind to TCF12 (Figure 4 [B]). Moreover, the overexpression of miR-218-5p inhibited the mRNA and protein expression of TCF12 in glioma cells (Figure 4 [C] and [D]). These results demonstrated that miR-218-5p could target TCF12 and inhibit its expression in glioma cells.
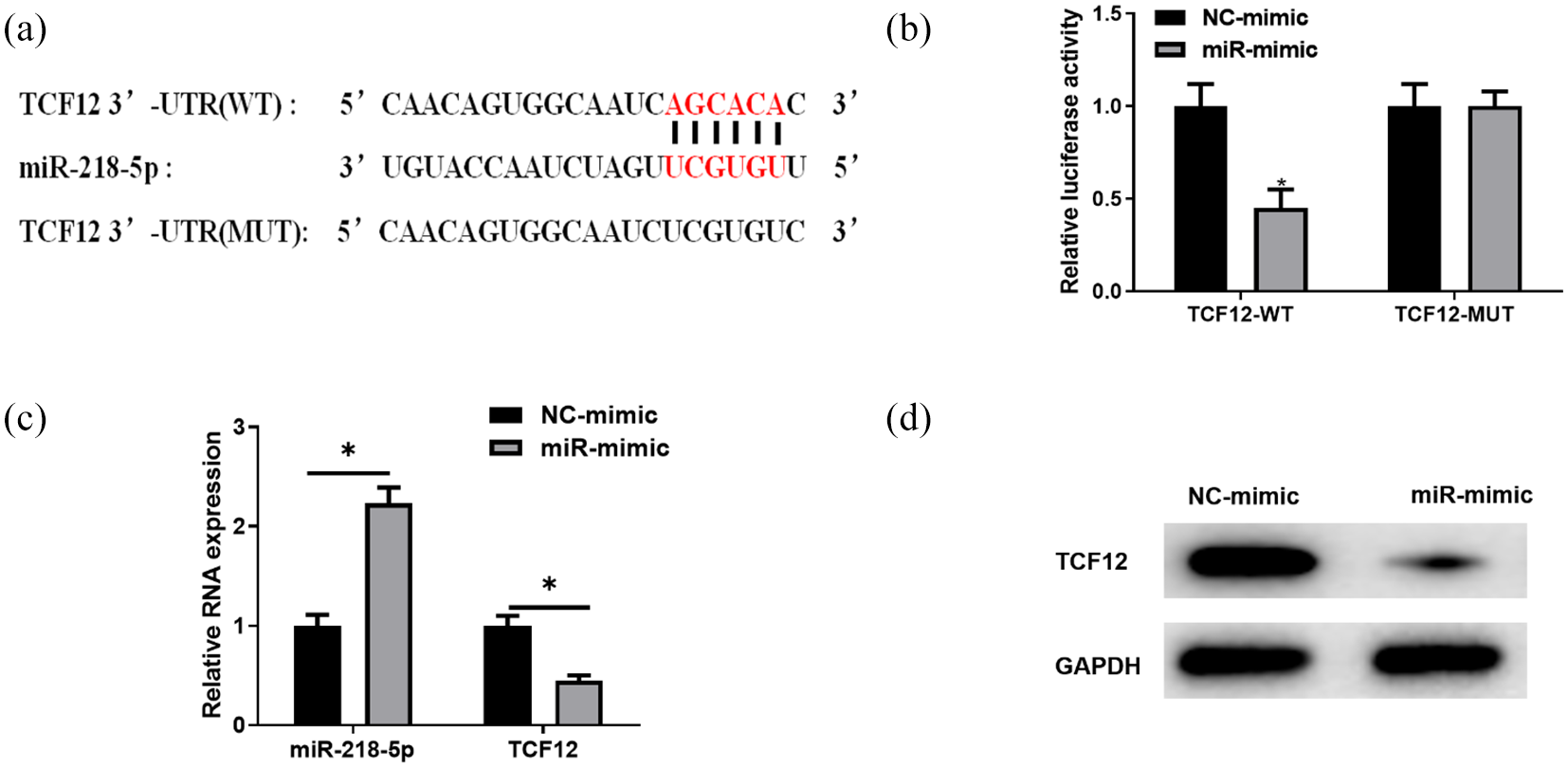
miR-218-5p targets TCF12 and inhibits its expression in glioma cells. (
miR-218-5p downregulates TCF12 to inhibit the proliferation, migration, and invasion of glioma cells and promote apoptosis
To verify the mechanism of the miR-218-5p/TCF12 axis in glioma cells, we constructed cell strains with overexpressed miR-218-5p (miR-mimic + oe-NC) and simultaneously overexpressed TCF12 and miR-218-5p (miR-mimic + oe-TCF12). The expression of TCF12 in cell lines was detected by qRT-PCR and Western blot. The results showed that the expression of TCF12 was significantly downregulated in the cells overexpressing miR-218-5p, while there was a marked increase of TCF12 expression in miR-mimic + oe-TCF12 compared with that in the miR-mimic + oe-NC group (Figure 5 [A]). CCK-8 assay showed that overexpressed miR-218-5p inhibited the proliferative capacity of glioma cells, whereas simultaneously overexpressing TCF12 and miR-218-5p would attenuate the inhibitory effect of overexpressed miR-218-5p on the proliferative capacity of glioma cells (Figure 5 [B]). Transwell and wound healing assays showed that overexpressed miR-218-5p downregulated the invasion and migration capacities of glioma cells, while the inhibitory effect of overexpressed miR-218-5p on cell invasion and migration was attenuated after simultaneously overexpressing miR-218-5p and TCF12 (Figure 5 [C] and [D]). Cell apoptosis was examined by flow cytometry and it showed that overexpression of miR-218-5p significantly promoted the apoptosis of glioma cells, whereas simultaneously overexpression of miR-218-5p and TCF12 blocked the apoptosis-promoting effect of miR-218-5p on glioma cells (Figure 5 [E]). The above results demonstrated that miR-218-5p could inhibit the proliferation, migration, and invasion of glioma cells and promote their apoptosis by inhibiting TCF12 expression.
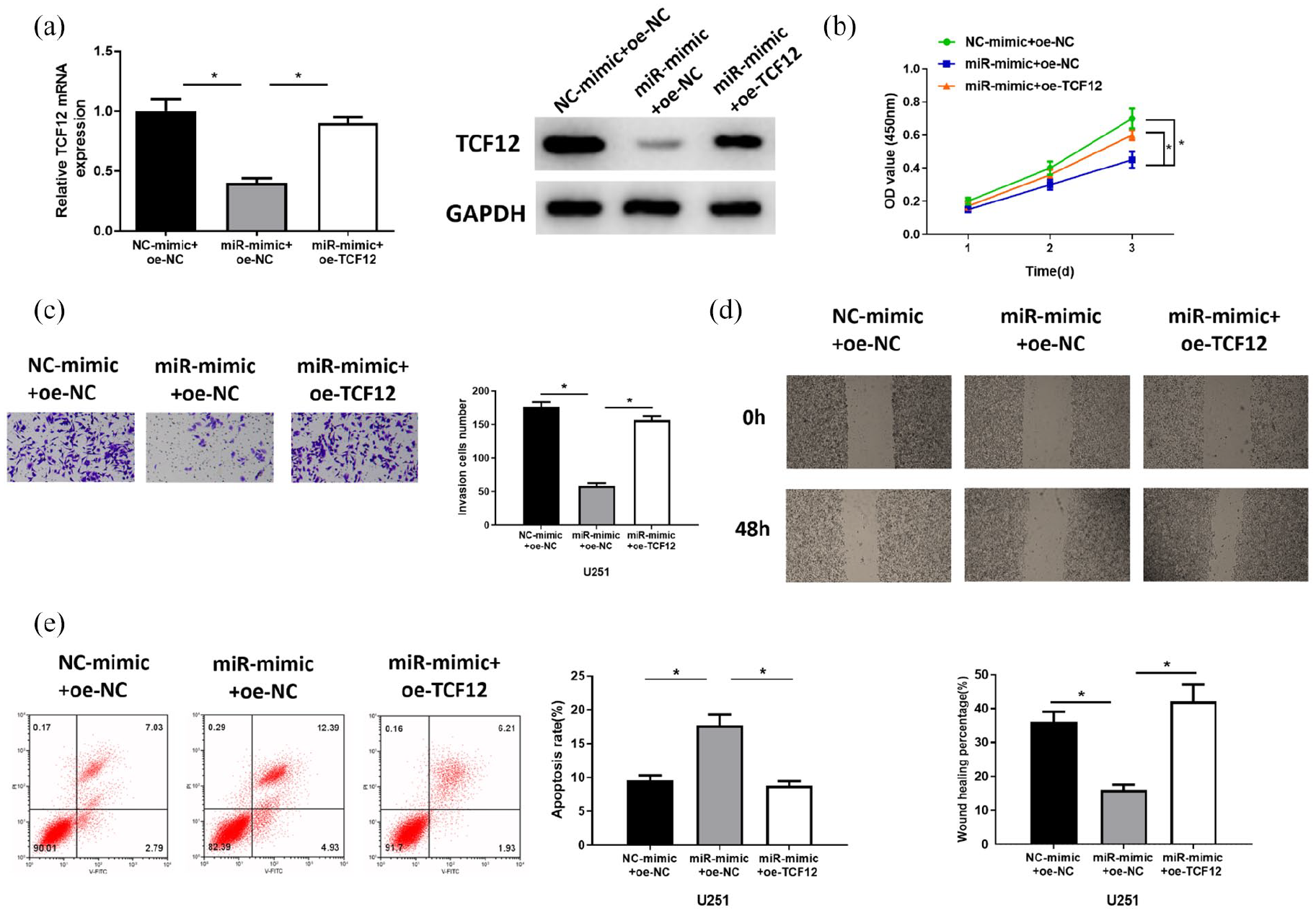
miR-218-5p inhibits the proliferation, migration, and invasion of glioma cells and promotes their apoptosis by targeting TCF12. (
Discussion
The dysregulation of mRNA translation is common in tumor cells, and many oncogenes and anticancer genes affect the translation mechanism, making aberrant translation a widespread feature of tumor cells. 17 Several studies have shown that dysregulation of mRNA expression can regulate the biological function of tumor cells and affect patient prognosis. For example, decreased expression of CDC25A can inhibit the proliferation and invasion of breast cancer cells and promote apoptosis. 18 The survival rate of patients with colorectal cancer with high TGFBI expression is significantly lower than that of patients with low TGFBI expression, and high TGFBI expression indicates a poor prognosis of patients with colorectal cancer. 19 It has been revealed that TCF12 protein is a transcriptional repressor of E-cadherin and its overexpression can promote the metastasis of colorectal cancer. 20 TCF12 is highly expressed in gastric cancer tissue and downregulation of TCF12 can inhibit tumor malignant process. 21 Overexpression of TCF12 can attenuate the inhibitory effect of miR-204 on cell migratory capacity of cervical cancer cells. 22 So we chose TCF12 as our study objective. We found that TCF12 expression was significantly upregulated in glioma tissue by bioinformatics analysis, and overexpression of TCF12 promoted the proliferation, migration, and invasion of glioma cells and inhibited their apoptosis, suggesting that TCF12 is an oncogenic factor in glioma cells.
A significant negative correlation between miR-218-5p and TCF12 was revealed by predicting upstream miRNAs of TCF12 using bioinformatics databases and Pearson correlation analysis in this study. To verify the existence of the targeting relationship between miR-218-5p and TCF12, we constructed TCF12 plasmids of wild-type and mutant type and verified their targeting relationship by dual luciferase assays. qRT-PCR and Western blot verified that miR-218-5p would inhibit the expression of TCF12. Here, we first put forward that miR-218-5p targets TCF12 and inhibits its expression in glioma cells. As one of the miRNA family members, miR-218-5p has been proved to play a critical role in kinds of cancers. For instance, miR-218-5p, downregulated in oral squamous cell carcinoma, can negatively regulate cell invasion by targeting CD44-ROCK, and it can be used as an effective therapeutic target for oral squamous cell carcinoma. 23 miR-218-5p inhibits cancer cell proliferation and migration via EGFR in non-small cell lung cancer. 15 In a separate study, miR-218-5p inhibited cell growth and metastasis in cervical cancer via the LYN/NF-κB signaling pathway. 16 The aforementioned studies demonstrate that miR-218-5p as a cancer suppressor in a variety of cancers is expected to be a potential biomarker for cancer diagnosis and treatment. In the present study, a series of cell experiments were performed to investigate the specific mechanism of the miR-218-5p/TCF12 axis in glioma cells. The results showed that overexpression of miR-218-5p inhibited the proliferation, migration, and invasion and promoted apoptosis of glioma cells, whereas overexpression of TCF12 attenuated the inhibitory effect of overexpression of miR-218-5p on glioma cells. It is shown that miR-218-5p downregulates the proliferation, migration, and invasion of glioma cells and promotes their apoptosis by targeting TCF12. In the study by Li et al., 24 upregulation of miR-218-5p decreased the proliferative and invasive capacities of glioma cells. Moreover, Gao et al. 25 believed that miR-218 could repress cell proliferation of glioma through targeting YY1. These results are similar to our results, demonstrating that miR-218-5p suppresses the malignant progression of glioma by acting as an oncogenic factor.
We found that in glioma tissue, TCF12 expression was upregulated and miR-218-5p expression was downregulated, and they were significantly negatively correlated. miR-218-5p inhibited the proliferation, migration, and invasion of glioma cells and promoted their apoptosis by targeting TCF12. Our study first sheds light on the regulatory role of the miR-218-5p/TCF12 axis in glioma cells, deepening our understanding of the molecular mechanism of miR-218-5p in gliomas. This finding provides new therapeutic strategies for patients with glioma, and enables miR-218-5p to become a potential biomarker for the diagnosis and treatment of glioma. Nevertheless, this study has certain limitations. Further work needs to be done in vivo to unravel the impact of miR-218-5p/TCF12 axis on the growth and metastasis of glioma. We will also dig deeper into the downstream signaling pathways of TCF12, thereby improving the molecular mechanism of miR-218-5p on the malignant progression of glioma.
Footnotes
Availability of data and materials
The data used to support the findings of this study are included within the article. The data and materials in the study are available from the corresponding author on reasonable request.
Authors’ contributions
All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.