
Abstract
Aims:
Although high expression of ZBTB7A is positively relative to metastasis in nasopharyngeal carcinoma (NPC) patients, the association between its low expression and metastasis of NPC remains unclear. The present study aimed to definitely identify the association.
Methods:
The level of ZBTB7A was effectively knocked down by stable transfection of short hair RNA plasmid in NPC cell lines CNE2 and 5-8F (shRNA-CNE2 and shRNA-5-8F), compared with the cells that stably transfected empty plasmid (NC-CNE2 and NC-5-8F). The levels of ZBTB7A were assessed by real-time polymerase chain reaction and Western blot in the cell lines. MTT assay, colorimetric focus-formation assay, flow cytometry, wound healing assay, transwell assays, and xenograft model were performed to analyze cell vitality, proliferation, cell cycle, migration, invasion, and tumorigenicity.
Results:
The levels of ZBTB7A were effectively reduced in shRNA-CNE2 and shRNA-5-8F. Their carcinogenicity was stronger separately than the abilities of NC-CNE2 and NC-5-8F. NC-CNE2 and shRNA-CNE2 were selected to establish the xenograft model because of their stronger tumorigenicity than NC-6-10B and shRNA-5-8F. The assay showed that shRNA-CNE2 had stronger tumorigenicity than NC-CNE2.
Conclusions:
The results demonstrated the reverse association between the expression of ZBTB7A and the tumorigenicity of NPC. We postulate that some oncogenic pathways, which are suppressed by ZBTB7A, will vicariously promote the proliferation and progression of NPC when ZBTB7A is decreased.
Introduction
Nasopharyngeal carcinoma (NPC) is an endemic disease in the Guangxi Province, South China (1-3). Recurrence and metastasis severely interfere with the survival quality of NPC patients. The development of distant metastasis shows that some regulative mechanisms remain unclear (4). The tumor metastasis related factors, such as Epstein-Barr virus (5), tumor metastatic genes, tumor metastatic suppressor genes (6-8), cell adhesion molecules (9), and noncoding RNAs (10-12), play important roles in regulating the metastatic mechanisms of NPC. These factors contribute to create a complex network of regulation, which can promote or suppress the metastasis of NPC (13-15).
ZBTB7A is also called Pokemon, FBI-1, OCZF, and LRF. ZBTB7A has a very important impact on human cancer, and acts in an oncogenic or oncosuppressive role (16, 17). For example, ZBTB7A mainly plays an oncogenic role in hepatocellular carcinoma (HCC). The proliferation and invasiveness of HCC cell lines are suppressed by the transient and stable knockdown of ZBTB7A (18, 19). ZBTB7A mainly plays an oncosuppressive role in melanoma through stable knockdown (20, 21).
It seems that ZBTB7A has been proved as a proto-oncogene in our early studies (22, 23). The previous results showed that the high protein expression of ZBTB7A was connected with NPC development in most of patients, compared to patients with chronic rhinitis (22). In contrast, the viability, migration, and invasion of CNE3 sublines were weakened by the transient knockdown of ZBTB7A (23). It is well known that the stable knockdown of ZBTB7A can provide an ideal model for studying metastatic mechanisms (20, 21). We constructed short-hair RNA (shRNA) plasmid and effectively knocked down the expression of ZBTB7A through transient transfection (24). In order to exactly clarify the role of ZBTB7A, we used the high-metastatic potential NPC cell lines, CNE2 and 5-8F, as research objects (25).
Methods
Cell culture
CNE2 and 5-8F cell lines were derived from NPC patients. CNE2 was preserved in the Research Center of Medical Sciences, The People’s Hospital of Guangxi Zhuang Autonomous Region (Nanning, Guangxi, China). 5-8F was obtained from the State Key Laboratory of Oncology in South China, Sun Yat-Sen University Cancer Center (Guangzhou, Guangdong, China). CNE2 and 5-8F were grown in RPMI Medium 1640 basic with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.).
Stable knockdown of ZBTB7A in NPC cell lines
CNE2 and 5-8F were seeded in 12-well plates at a density of 104 cells/well. Transfection was performed as 4 μg plasmid: 10 μL Lipofectamine® 3000 (Invitrogen) when the cells reached 50% confluence. They were transferred to 96-well plates by serial dilution after 48 hours (23). The cells transfected with plasmids were selected by G418. The best concentrations were 300 μg/mL (CNE2) and 250 μg/mL (5-8F). The concentration was reduced to 150 μg/mL and 125 μg/mL, respectively, after 10 days. The clone cells were cultured stably during 15 passages. The cells stably transfected with plasmids were called NC-CNE2, shRNA-CNE2, NC-5-8F, and shRNA-5-8F.
Total RNA extraction and real-time PCR
Total RNA of the transfected cells was extracted by TRIZOL (Invitrogen). RNA concentration was detected by Nanodrop 2000 (Thermo Fisher Scientific, Inc.), and 4 μg total RNA was analyzed by 1.2% agarose gel electrophoresis with 0.6 mol/L formaldehyde. The gel was photographed by Bio Imaging System Gene Genius (Syngene). An equal amount (4 μg) of total RNA was reverse transcribed using the RevertAid First Strand cDNA Synthesis Kit (Fermentas; Thermo Fisher Scientific, Inc.). For real-time PCR, the ZBTB7A (NM_015898.2) primers were: Forward, 5’-GCTTGGGCCGGTTGAATGTA-3’, Reverse, 5’-GGCTGTGAAGTTACCGTCGG-3’. GAPDH (NM_002046.4) was used as an internal reference control: Forward, 5’-CATGAGAAGTATGACAACAGCC-3’, Reverse, 5’-AGTCCTTCCACGATACCAAAGT-3’. The primers of ZBTB7A and GAPDH were synthesized by Sangon Biotech Co. The reaction condition referred to SuperReal PreMix Plus (SYBR Green) Kit (Tiangen). The reaction program consisted of an initial denaturation step at 95°C for 10 minutes, denaturation at 95°C for 15 seconds, and annealing at 60°C for 60 seconds for 40 cycles; and the dissociation stage at 95°C for 15 seconds, 60°C for 1 minute, 95°C for 15 seconds, and 60°C for 15 seconds. The reaction program was executed by 7500 real-time PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc.). The mRNA expression level of ZBTB7A was calculated by the 2 Delta-delta Ct method.
Protein extraction and Western-blot
Total protein was extracted from the transfected cells using the 99% radio immunoprecipitation assay lysis buffer and 1% protease inhibitor Phenylmethanesulfonyl fluoride (PMSF) (Beyotime). The final concentration of PMSF was 1%. The protein was quantified using the Bradford method. For Western-blot, equal amount protein samples (20 μg) were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto nitrocellulose membranes (Bio-Rad Laboratories, Inc.). The membranes were incubated with the appropriate antibodies (1:500 dilution, catalog no., ab70208, rabbit polyclonal; anti-ZBTB7A antibody from Abcam, 1:1200 dilution, catalog no., AA128, mouse monoclonal; anti-β-actin antibody from Beyotime) overnight at 4°C, then incubated with horseradish peroxidase-conjugated anti-rabbit (1:8,000 dilution, catalog no., ZB-2301; ZSGB-BIO) or anti-mouse (1:12,000 dilution, catalog no., A0216; Beyotime) for 2 hours at 37°C. The detection was performed by BeyoECL Plus (Beyotime).
MTT assay
CNE2 and 5-8F stably transfected cells were trypsinized and seeded in 96-well plates at a density of 1,500 cells/well and incubated with 0.5% of MTT (Sigma-Aldrich; Merck KGaA,) for 4 hours. The cells were lysed by dimethyl sulfoxide (DMSO, Sigma) and the absorbance was detected by GF-M3000 Elisa Analyzer (Caihong Analytical Instruments) at 490 nm.
Colorimetric focus-formation assay
CNE2 and 5-8F stably transfected cells were seeded in 6-well plates at a density of 100 cells/well, fixed by 100% methanol after 2 weeks, and dyed by 5% crystal violate (Amresco) in 100% methanol.
Cell cycle analysis by flow cytometry
CNE2 and 5-8F stably transfected cells were seeded in 6-well plates at a density of 106 cells/well. The cells were harvested and washed in cold phosphate-buffered saline (PBS) after 24 hours. Propidium iodide staining was carried out using Cycle Test Plus DNA Reagent Kit (BD Biosciences), according to the manufacturer’s protocol. After incubation for 10 minutes in the dark at room temperature, the cells were analyzed by FACSCalibur Flow Cytometry (BD Biosciences).
Wound healing assay
CNE2 and 5-8F stably transfected cells were seeded in 6-well plates at a density of 106 cells/well. A wound was created through manually scraping the cell monolayer with a 200 μL pipette tip after 24 hours. The cultures were washed 3 times with PBS to remove floating cells. The cells were cultured. Cell migration was observed at 2 time points (0 h and 24 h) in 5 randomly selected microscopic fields for each condition and each time point. Images were acquired with a IX71 fluorescence microscopy (Olympus; original magnification: ×40). The moving distance of the cells was calculated through measuring the wound width at different time points, then subtracting the wound width at time 0 hours.
Transwell migration and invasion assays
Transwell migration and invasion assays were performed in 6.5 mm transwell chambers of 24-well plates (pore size, 8 μm; Costar). 2 × 105 cells of CNE2 and 5-8F transfected cells were resuspended in 200 μL serum-free 1640 medium in the upper surface, and 800 μL medium containing 10% FBS was added to the lower surface. For in vitro invasion assays, the upper surface was coated by 70 μL dilution of Matrigel Basement Membrane Matrix (BD Biosciences); 17.5 μL matrigel was diluted by 52.5 μL serum-free 1640 medium. The dilution was incubated for 4 hours at 37°C. After 24 hours (migration assay) and 48 hours (invasion assay) of incubation, the nonmigratory and noninvasive cells on the upper surface of filters were completely removed by cotton swabs. The migratory and invasive cells on the lower surface of the filter were rinsed with PBS, then fixed with methanol and stained with 1% crystal violet. The migratory and invasive cells were quantitated by visual counting (BX51 microscopy, Olympus; original magnification: ×200). The mean of 5 individual fields in the center of the filter was obtained for each well.
Xenograft model
Based on the above assays, we selected NPC cell lines NC-CNE2 and shRNA-CNE2 to establish the tumor model because they had stronger tumorigenicity than NC-5-8F and NC-6-10B. Balb/c pure line mice were randomly grouped from the Guangxi Medical University Laboratory Animal Centre (8 mice/group). The mice were female, 4 weeks old, weighed 15 g, and were randomly divided into 2 groups. Every nude mouse was subcutaneously injected with 5 × 106 cells/200 L PBS in each group. The volume of primary tumors was measured and calculated by using the formula V = (π/6)(d1 × d2)3/2 for 27 days, after which the weight of the tumors was measured. The study was approved by the ethics committee of The People’s Hospital of Guangxi Zhuang Autonomous Region.
Statistical analysis
All assays were performed in 3 independent experiments, except for the animal assay, and all data were expressed as the mean ± standard deviation. Data analysis and drawing were performed by Sigma Plot 12.5 (SPSS Inc.). Statistical differences of 2 groups were evaluated with independent-samples T-test and analysis of variance (ANOVA) was used for multiple comparisons (SPSS 13.0). In all cases, p<0.05 was considered to be statistically significant.
Results
ZBTB7A levels of CNE2 and 5-8F stably transfected with shRNA plasmid decreased
CNE2 and 5-8F cells were stably transfected with shRNA coding plasmid targeting ZBTB7A. They were named shRNA-CNE2 and shRNA-5-8F. The cells stably transfected with empty plasmid were used as negative controls (NC) and named NC-CNE2 and NC-5-8F. The results indicated that mRNA and protein levels of ZBTB7A were efficiently reduced in shRNA-CNE2 and shRNA-5-8F (Fig. 1).
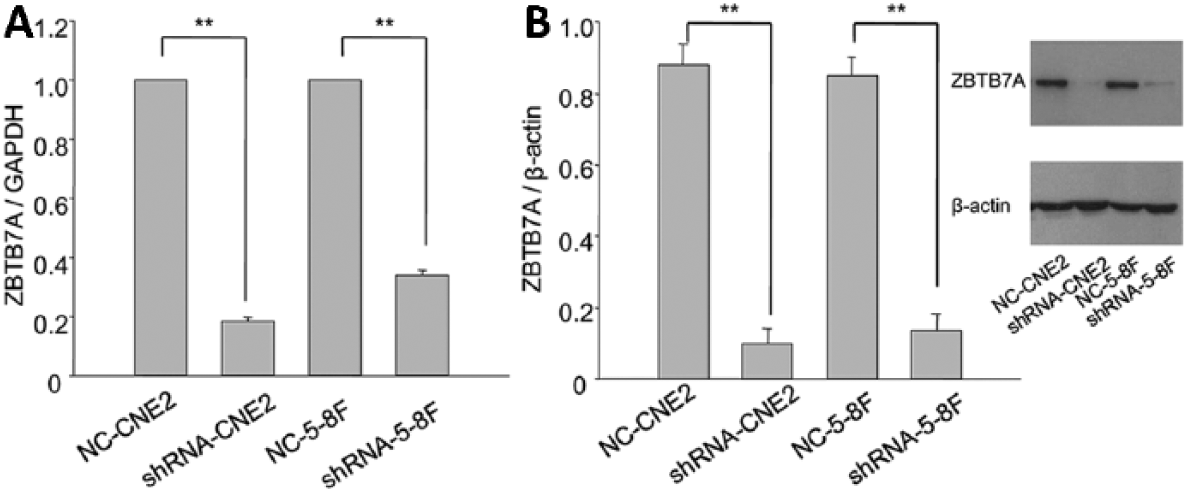
ZBTB7A levels of CNE2 and 5-8F stably transfected cells. (
The vitality and proliferation of shRNA-CNE2 and shRNA-5-8F strengthened
The vitality of shRNA-CNE2 and shRNA-5-8F was stronger than that of NC-CNE2 and NC-5-8F, respectively (Fig. 2A). The proliferation of shRNA-CNE2 and shRNA-5-8F was stronger than that of NC-CNE2 and NC-5-8F (Figs. 2B, C).
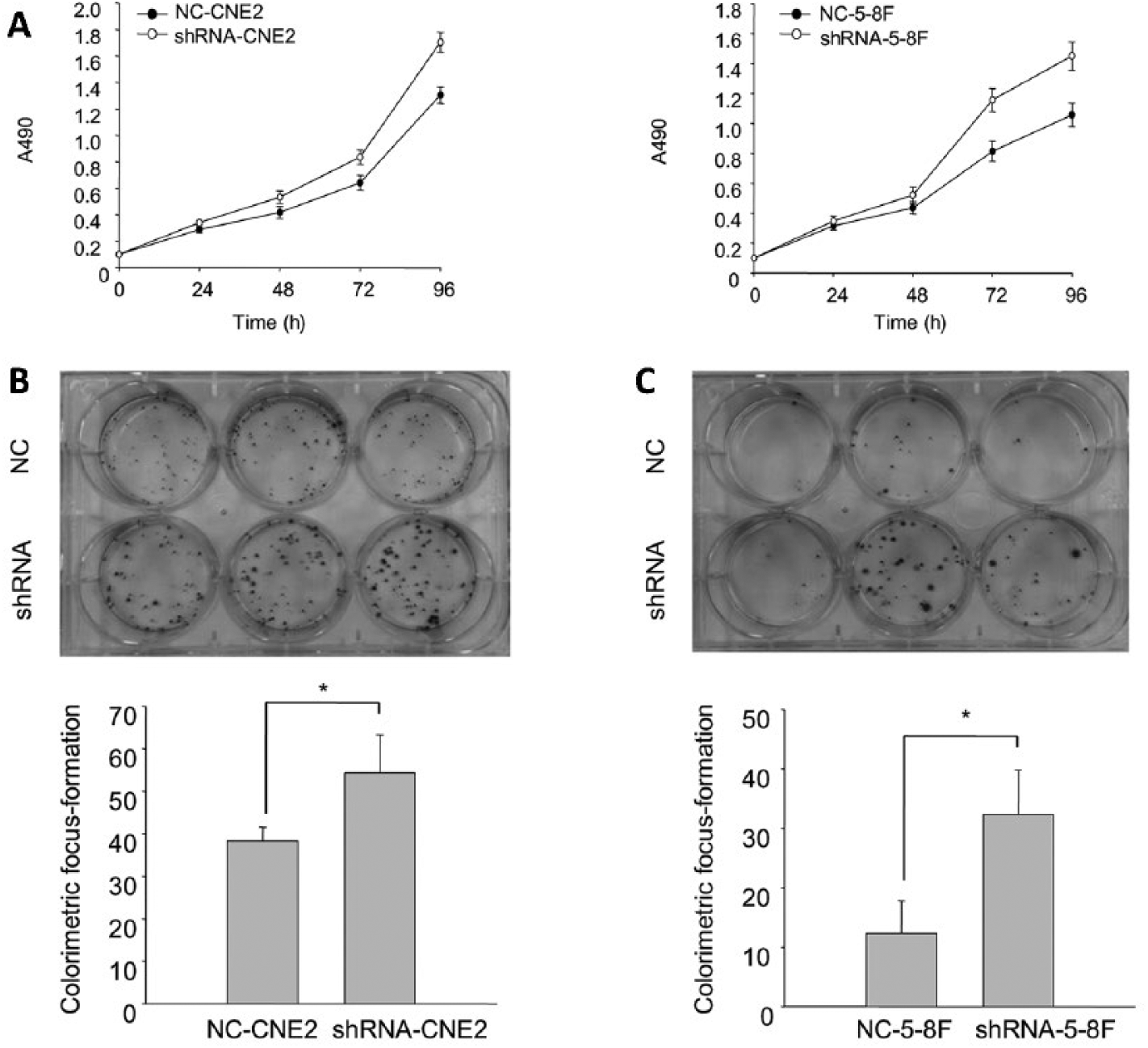
The vitality and proliferation of CNE2 and 5-8F stably transfected cells. (
The percentage of shRNA-CNE2 and shRNA-5-8F cells in G0-G1 phase decreased
Flow cytometry analysis showed that knockdown of ZBTB7A markedly decreased the percentage of shRNA-CNE2 and shRNA-5-8F cells in the G0-G1 phase, while it increased the percentage of the cells in the S phase (Fig. 3).

The cell cycle phases of CNE2 and 5-8F stably transfected cells. The cell cycles of NC-CNE2, shRNA-CNE2, NC-5-8F, and shRNA-5-8F were analyzed by FCM. **p<0.01. FCM = flow cytometry.
The migration and invasion of shRNA-CNE2 and shRNA-5-8F strengthened
For wound healing assays, the migratory rates of shRNA-CNE2 and shRNA-5-8F were higher than those of NC-CNE2 and NC-5-8F cells, respectively (Fig. 4A). For transwell assays, the abilities of the migration and invasion of shRNA-CNE2 and shRNA-5-8F were stronger than those of NC-CNE2 and NC-5-8F, respectively (Figs. 4B, C).
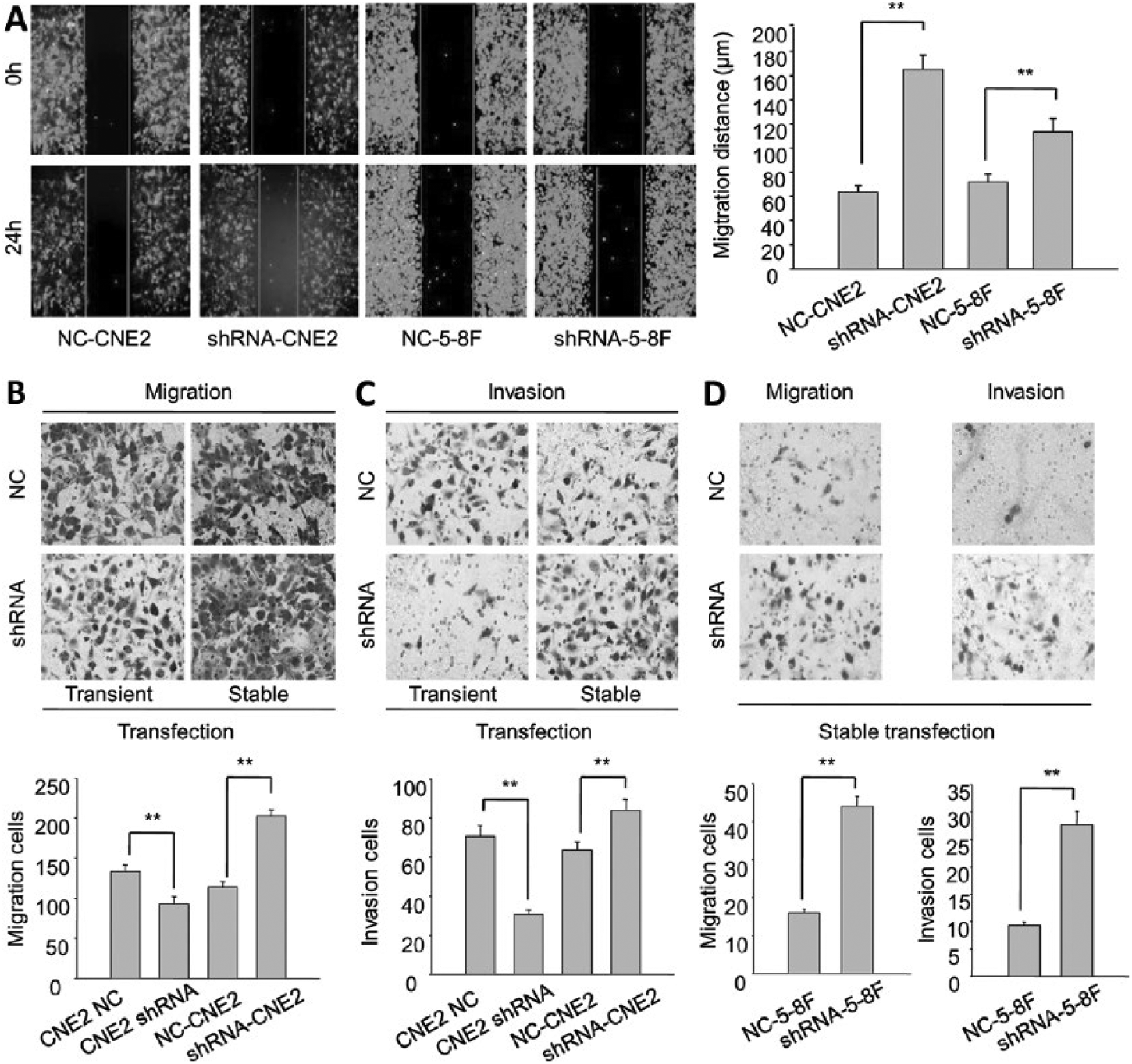
The migration and invasion of CNE2 and 5-8F stably transfected cells. (
The tumorigenicity of shRNA-CNE2 in the xenograft model was higher than NC-CNE2
On the basis of the obtained results, we demonstrated that the tumorigenicity of NC-CNE2 cells was higher than that of NC-5-8F cells. Therefore, we selected NC-CNE2 and shRNA-CNE2 to establish a xenograft model. The volume of tumors formed from a subcutaneous injection with shRNA-CNE2 was larger than that from NC-CNE2. The weight of tumors formed from a subcutaneous injection with shRNA-CNE2 was higher than that from NC-CNE2 (Fig. 5).
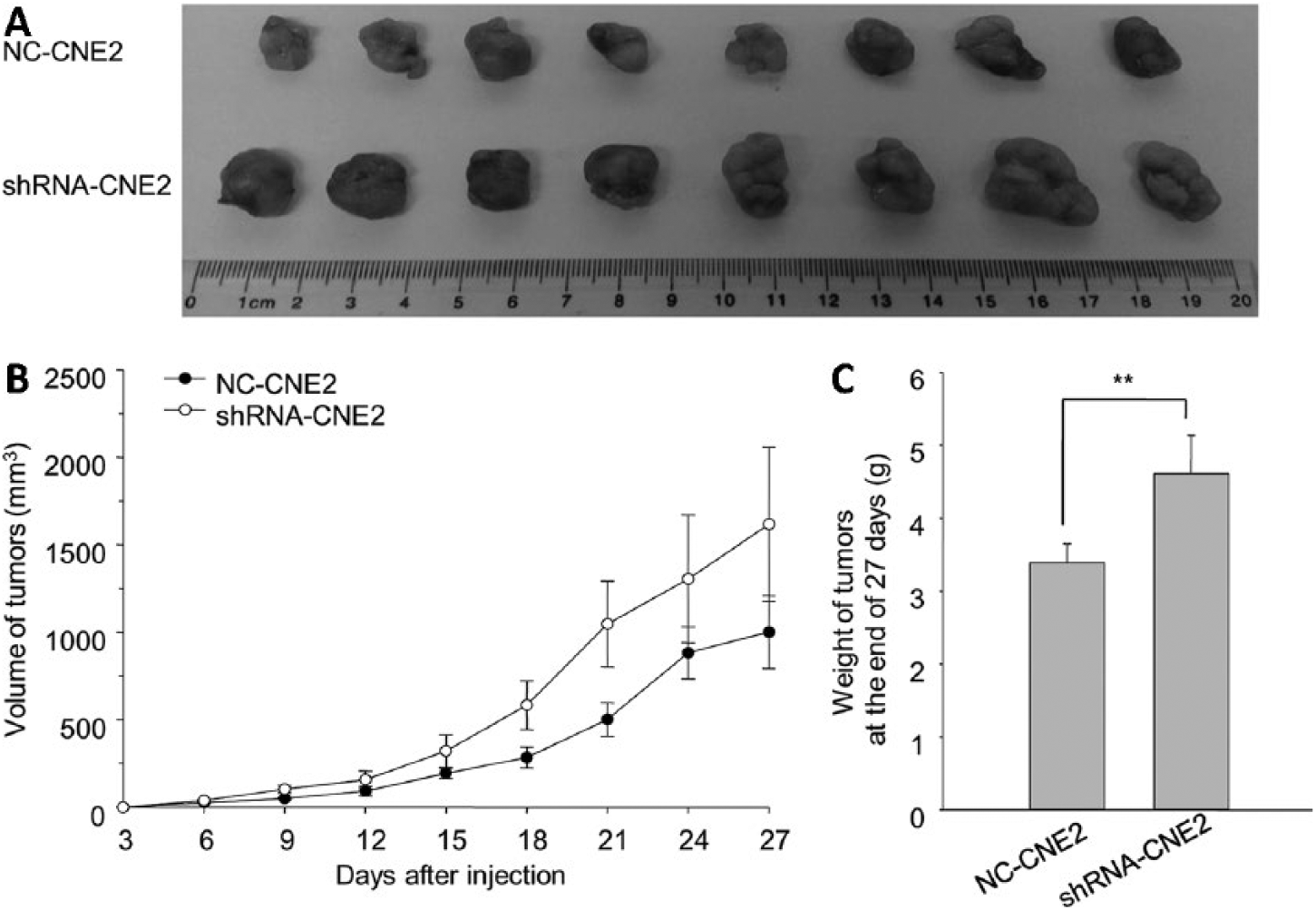
The comparison of xenograft model between NC-CNE2 and shRNA-CNE2. (
Discussion
Since the best ZBTB7A shRNA plasmid was constructed and screened (24), we detected the carcinogenic changes of NPC cell lines with ZBTB7A being knocked down. At the beginning, we found that the viability, proliferation, migration, and invasion of CNE2 was transiently transfected with ZBTB7A shRNA plasmid for 48 hours was weaker than in the control groups that were transiently transfected with an empty vector (Supplemental Figs. S1, S2, available online at www.tumorijournal.com), as were the CNE3 sublines (23). The results suggest that ZBTB7A is a proto-oncogene in NPC.
However, strikingly opposite results appeared when ZBTB7A was knocked down stably in CNE2 and 5-8F. The viability, proliferation, migration, and invasion of shRNA-CNE2 and shRNA-5-8F were separately stronger than NC-CNE2 and NC-5-8F. Fortunately, the tumor suppressor role of ZBTB7A has been reported by Harvard University (20, 21, 26, 27). The studies have a certain reference value for explaining our results. We hypothesize that ZBTB7A controls oncogenic and oncosuppressive pathways at the same time. If the level of ZBTB7A is notably increased, the proto-oncogene will play an important role in NPC. If the level of ZBTB7A is obviously decreased, some oncogenic pathways that are suppressed by ZBTB7A vicariously promote the proliferation and progression of NPC cells.
In order to confirm the hypothesis, we used mRNA expression profile chips to select differentially expressed mRNAs. Some of them are closely connected with metastatic cancer, especially NPC (data not shown). We hypothesize that they are probably regulated by ZBTB7A in NPC cells. It is necessary to speculate important pathways that are regulated by ZBTB7A. It is likely that part of them has an important influence on NPC progression. We have recently found that a low level of ZBTB7A came from a few NPC patients with advanced stage, but the regulatory mechanism of ZBTB7A is still unknown (data not shown). In order to clarify the association between tumor-related genes and NPC, we will keep on detecting the levels of the differentially expressed mRNAs of NPC tissues.
Supplemental Material
TMJ753301_Supplementary_Figure_S2 – Supplemental material for Stable knockdown of ZBTB7A promotes cell proliferation and progression in nasopharyngeal carcinoma
Supplemental material, TMJ753301_Supplementary_Figure_S2 for Stable knockdown of ZBTB7A promotes cell proliferation and progression in nasopharyngeal carcinoma by Fei Liu, Fengzhu Tang, Jiao Lan, Wei Jiao, Yongfeng Si, Wensheng Lu, Mingzheng Mo, Bing Li, Jinlong Lu, Jiazhang Wei, Ying Qin, Ruiping Xiao, Benjian Zhang,Yongli Wang, Weiming Xiong in Tumori Journal
Footnotes
Acknowledgements
The authors are very grateful to Professor Musheng Zeng at Sun Yat-Sen University Cancer Center for generously providing the 5-8F cell line.
Disclosures
Financial support: This study was supported by the Natural Science Foundation of Guangxi Province (2015GXNSFAA139166, 2016GXNSFBA380144, 2016GXNSFCB380003), Guangxi Scientific Research and Technology Development Project (14124003-1-3).
Conflict of interest: None of the authors has financial interested related to this study to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.