
Abstract
Sensory receptors were described and illustrated after they had been observed with the aid of microscopes. Most descriptions were made in the 19th century after the introduction of achromatic lenses in microscopes. In some senses (like vision), receptors were named according to their morphology whereas in others (like touch), they are known by the names of those who initially described them. Illustrations of the receptors from original sources are here combined with portraits of their originators.
Introduction
Cells were described soon after the first microscopes were focused on animal matter (see Lancaster, 2014). Hooke (1665) gave them their name and identified plant cells. The senses also came under similar early microscopic scrutiny. The application of microscopes to biological specimens was a huge leap assisting understanding, but the microscopes themselves were neither powerful nor free from optical aberrations; the techniques for preparing specimens for observation were also wanting (Turner, 1998). The gross features of the nervous system had been examined with the naked eye, but a new world was exposed by the microscope, and this world was examined by Malpighi (1665). As a consequence of the many biological structures he examined microscopically (both plant and animal), Malpighi is often regarded as the first histologist. In addition to chick embryos, where he provided one of the first accurate descriptions of development in the nervous system, he examined the brain (including the optic nerve), the skin (which now has a Malpighian layer) and the tongue. He applied to the interpretations of physiology the mechanistic concepts derived from the Galilean school; microscopic organization of living tissues was seen as based on the functioning of a multitude of minute machines. The type of microscope Malpighi used is not known (Motta, 1998) unlike Hooke (1665) who gave textual detail and illustrated his compound microscope; Hooke also described a more powerful simple microscope (Ford, 2007). It was with such a device that van Leeuwenhoek (1675) observed a variety of animal cells, including nerve fibres. Microscopes improved in design and in the quality of lenses within them during the 18th century (for a survey of early microscopes, see Mayall, 1886) but interest in focusing the instruments on sensory receptors was not at the forefront.
The microscopic world was transformed by the introduction of powerful achromatic instruments in the 1830s, and rapid advances were made thereafter (Harris, 1999; Schickore, 2007). Among those who combined the achromatic microscope with remarkable observational skills was Purkinje (1837). In 1832, Purkinje obtained an achromatic microscope (Chvátal, 2017) and directed it at the large cells in the cerebellum, thereby identifying the cells that bear his name. His microscopic observations were made before any adequate staining methods had been developed. Purkinje used alcohol to fix his preparations, and he made thin sections so that they could be examined microscopically. Purkinje’s laboratory at Breslau (present-day Vroclaw) has been described as the cradle of histology, and it was matched only by that in Berlin, established by Johannes Müller (Otis, 2007). The cell doctrine was most clearly articulated by Schleiden (1838) for plants and Schwann (1839) for animals and it is associated with their names. At the end of the century, Waldeyer (1891) extended the doctrine to nerves. He named the nerve cell body, its fibres and arborisations as a ‘Neurone’ and supported the theory that neurons were the fundamental structural and functional units of the nervous system. Some of these cell visionaries are shown in Figure 1.
Cell visionaries by Nicholas Wade. Upper left, cork cells as illustrated by Robert Hooke (1635–1703; Hooke, 1665) contain portraits of Matthias Schleiden (1804–1881) and Theodor Schwann (1810–1882). Upper right, Antonius van Leeuwenhoek (1632–1723) together with the simple microscope through which he viewed biological tissues. Lower left, Wilhelm von Waldeyer (1836–1921) combined with an illustration of neurons. Lower right, a portrait of Jan Evangelista Purkinje (1787–1869), a Plössl achromatic microscope of the type he used and the illustration of the large cerebellar cells (derived from Purkinje, 1837) that bear his name.
Among the microscopic structures that were isolated and described after the cell doctrine had been enunciated were specialized sensory cells, called receptors, and they could be related to the stimuli that excited them. Those located in well-defined sense organs were named on the basis of their morphology (rods, cones, hair cells, etc.), whereas the receptors in or beneath the surface of the skin were generally named after those who first described them (e.g. Golgi tendon organs, Krause end-bulbs, Meissner corpuscles, Merkel discs, Pacinian corpuscles and Ruffini cylinders). It is clear that there are many specialised sensory cells. They will be examined here with regard to the retina, inner ear, papillae on the tongue, olfactory cells and receptors in the skin, muscles and tendons. Illustrations of cell microanatomies as illustrated by those who described them will be combined with portraits of the scientists who drew attention to them. I call these ‘perceptual portraits’ (see Wade, 1995, 2006, 2016, 2017).
Retina
The concept that there were specialised receptors arrived relatively late on the sensory scene. In the 17th century, when Descartes (1637/1902) was discussing the eye, he considered that the retina consisted of nerve endings and that their dimensions defined the limits of visual resolution (see Wade, 2004). As these were of a particular size, he argued that no object smaller than a fibre ending could be resolved. This relationship was maintained even when the microanatomy of the retinal cells was better understood. Treviranus (1837, Figure 2) presented drawings based on vertical and horizontal microscopic sections of cells in the visual systems of many species. His diagram of the crow’s retina indicated a wider variation in retinal structure than had previously been represented, and the layers within it are clearly shown. The papillae were directed towards the incoming light rather than away from it. In this regard, Treviranus was reflecting Descartes’ idea that the terminations of the optic nerves were the receptive elements in the retina, and that they were directed towards the lens. The years following 1840 saw rapid advances in fixing, sectioning and staining microscopic preparations (Finger, 1994; Harris, 1999; Schickore, 2007). Nonetheless, Treviranus described and illustrated cylindrical cells in the retinas of a variety of animals, and opened the way for others to examine the microscopic structure of the retina in more detail. As Polyak (1957) remarked: ‘The work of Treviranus, though erroneous in almost every point, was beneficial because it stimulated an immediate series of investigations’ (p. 48). Treviranus was a comparative zoologist who did much to establish biology as an independent discipline within Germany, and to provide support for the cell doctrine. The correct anatomical orientation of the retinal elements was described shortly after Treviranus by Bidder (1839); the terminations of the optic nerve structures were directed towards the choroid rather than the lens. A decade later, Bowman (1847/1849) provided a diagram of the retina, which distinguished between what he called rods and bulbs (Figure 2). Bowman described the constituents of the retina in the following way: ‘The elements peculiar to the retina are … of two kinds – Columnar particles, or rods, arranged vertically in a single series; and Bulbous particles, interspersed at regular intervals among the former’ (p. 80). He drew attention to the disagreements about the interpretations of the terminal structures of the retina, and to the difficulties of obtaining good specimens for microscopical study. Bowman described differences in the numbers of rods and bulbs in different species, but no generalization was drawn from this.
The retinas of Treviranus, Bowman, Kölliker and Schultze by Nicholas Wade. Upper left, Gottfried Reinhold Treviranus (1776–1837) is shown in his diagram of the crow retina in his posthumously published volume on the inner structure of the retina (derived from Treviranus, 1837). Upper right, William Bowman (1816–1892) is portrayed within the diagram of the human retina derived from Bowman (1847/1849). Lower left, Alfred Kölliker (1817–1905) can be seen in his representation of the retina (derived from Kölliker, 1854). Lower right, Max Schultze (1825–1874) and his illustration of retinal structure, flanked by his diagrams of an isolated rod and cone (derived from Schultze, 1866).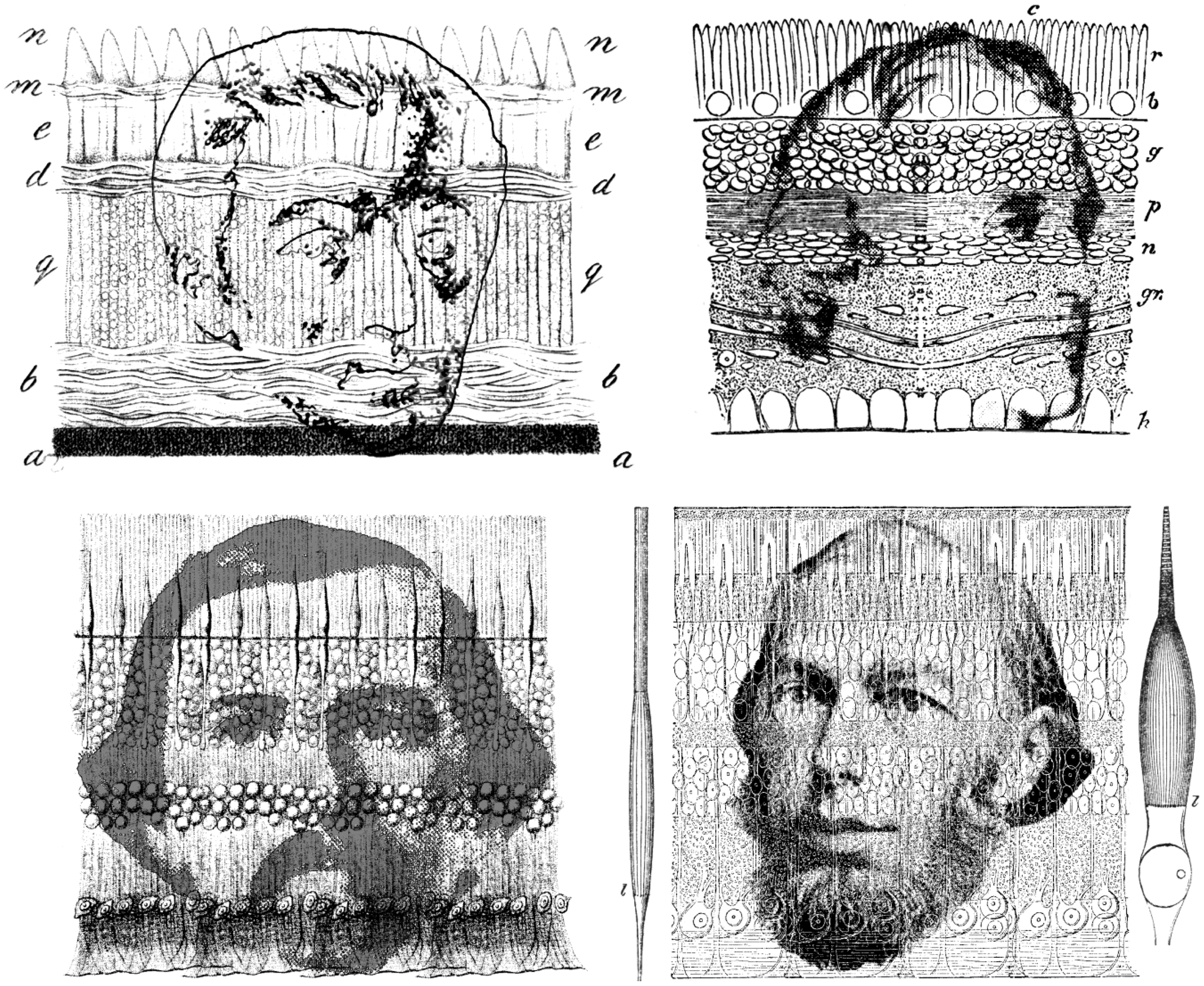
Helmholtz drew upon the burgeoning histological research that was emerging from German laboratories. The authorities on retinal structure were Kölliker (Figure 2) and his collaborator Heinrich Müller. The figure Helmholtz (1867) used to illustrate retinal structure in the first volume of his Handbuch der Physiologischen Optik was from Kölliker (1850), but he changed it in the second edition (Helmholtz, 1896) to that by Schultze (Figure 2), together with Schultze’s (1866) diagram of a single rod and cone. The numerical ordering of the layers in the retina was reversed, too (see Wade, 2007). For Kölliker’s diagram, the sequence started with the rod and cone layer; for Schultze’s diagram, the number of layers was extended to 10, and the sequence terminated with the choroid. The English translation of Helmholtz (1924) confounds the two accounts; the text is taken from the first edition, but Schultze’s diagram from the second edition replaces that of Kölliker. The third German edition (and therefore the English translation) is based on the text of Helmholtz’s first edition, and so contains this conflation.
Schultze succeeded Helmholtz in the chair of anatomy at Bonn in 1859, when Helmholtz moved to Heidelberg. In the first volume of his Handbuch, Helmholtz (1924) was able to state that ‘The retina is composed partly of the microscopical components of the nervous system (nerve fibres, ganglion cells and nuclei), and partly of certain characteristic elements, the so-called rods (bacilli) and cones (coni)’ (p. 24). Only cones were present in the fovea. Schultze (1866) also examined the complement of rods and cones in a variety of animals, and was able to suggest that rods and cones function under different levels of illumination – duplicity theory.
Bowman (1847/1849) referred to ‘rods’ and ‘bulbs’ as the two cell types in the retina but Hannover’s (1844) ‘cônes’ was the name for the latter that was generally adopted. In German, they were called Zapfen, and this is the term used by Helmholtz (1867).
Schultze’s (1866) suggestion of different functions for rods and cones instigated a search for differences in their chemical compositions that could account for them. Heinrich Müller (1851, Figure 3) observed the colours of the retinas of frogs: ‘The rods of frogs appear somewhat reddish, where they lie in a sufficient density over one another, and one can see single rods alternatively colourless or coloured, depending upon whether they are lying or upright’ (p. 236). Müller’s discovery of the coloured retinal receptors was not actively pursued for another two decades when his pioneering observation was acknowledged by Boll (1876, Figure 3). Boll (1877/1977) referred to the colour of the dark-adapted retina as ‘intense purple-red’ but he revised this in his second, longer publication to ‘visual red’ and this reddish colour became paler when exposed to light (see Baumann, 1977; Wade, 2008a). Kühne (1879/1977, Figure 3) extended Boll’s work but referred to the unbleached retinal chemical as ‘visual purple’ (see Wade, 2008b). Kühne extracted rhodopsin from the rods of frogs and rabbits and showed that the rate of bleaching was dependent not only on the intensity of light but also on its wavelength. The visual purple was confined to rods and was not seen in the foveas of humans. Most significantly, Kühne established the ‘visual cycle’: Visual purple in the rods is bleached by light to form visual yellow which in turn is transformed to visual white.
The red retinas of Müller, Boll and Kühne by Nicholas Wade. Left, Heinrich Müller (1820–1864) described the red retina and also investigated the Purkinje tree. Centre, a portrait of Franz Boll (1849–1879) is contained within a motif of retinal rods (modified from a detail of an illustration in Boll, 1877/1977). Right, the visage of Willy Kühne (1837–1900) is combined with diagrams of the retinal rods of dark adapted frogs (derived from a figure in Kühne, 1879/1977).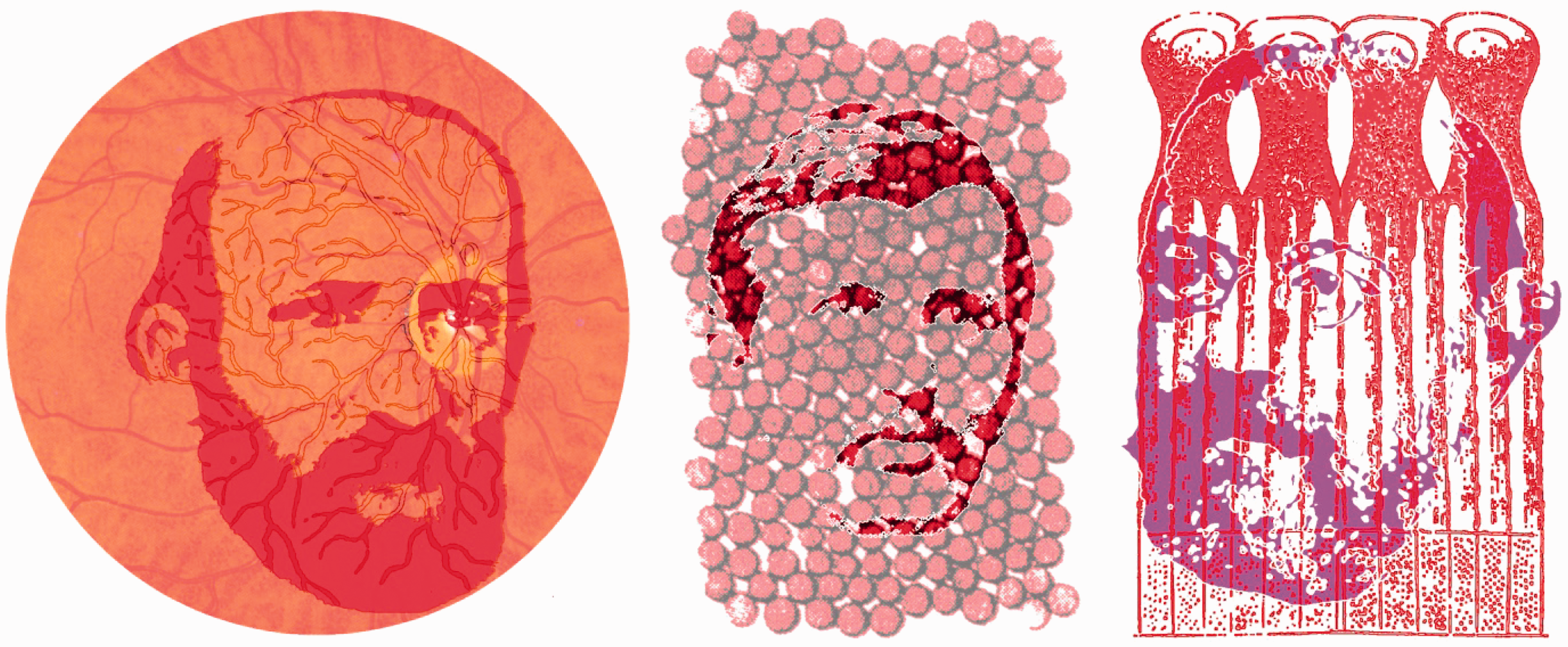
Inner Ear
The location of the inner ear within the bony labyrinth made it difficult to dissect and delayed investigation into its structure. In one of the earliest books dedicated to the ear and hearing, Du Verney (1683) lamented the state of studies of that sense:
Of all the Organs assign’d to the Use of Animals, we have the least Knowledge of those of the Senses; but there is none more obscure than that of Hearing: the Minuteness and Delicacy of the Parts which compose it, being inclos’d by other Parts, (which by reason of their Hardness, are scarcely penetrable) render the Enquiries into them more difficult, and their Structure something so intricate, that there is as much Trouble in explaining, as there is in discovering them. (1737, p. vii)
Despite these difficulties, Scarpa (1789; Figure 4) provided detailed diagrams of the cochlear and vestibular system. This paved the way for his fellow countryman, Corti (1851; Figure 4), to provide some detail of the microanatomy of the cochlea.
The inner ears of Scarpa, Corti, Deiters, Retzius, Held and Wersäll by Nicholas Wade. Upper left, Antonio Scarpa (1752–1832) produced detailed grey-scale representations of the cochlea and vestibular apparatus as well as outline drawings, both of which are shown (derived from Scarpa, 1789). Upper right, Alfonso Corti (1822–1876) can be seen within his coloured illustration of the structure and hair cells on the basilar membrane of the cochlea (derived from Corti, 1851); the plate was folded in the journal which accounts for the differences in the intensity of the background. Centre left, Otto Deiters (1834–1863) illustrated the cells (f in the illustration) supporting the hair cells (derived from O. Deiters, 1859). Centre right, one of the diagrams by Gustaf Retzius (1842–1919) of the structure of the organ of Corti (derived from Retzius, 1884) together with his portrait. Lower left, a portrait of Hans Held (1866–1942) and figures from Held (1909) illustrating cells in the saccule of rabbits and pigeons. Lower right, Jan Wersäll (1930–2017) and his schematic illustration of Type I and Type II cells from the ampulla of guinea pig (derived from Wersäll, 1956).
Scarpa added greatly to knowledge of the anatomy of the senses as well as of the brain (Grzybowski & Sak, 2013). He made important discoveries on the anatomy of the internal ear and of the vestibular system (as attested by Scarpa’s ganglion). He described accurately the innervation of the heart, correcting the commonly held error that the heart lacked nerves. He was also a great surgeon and he developed new surgical approaches particularly in the treatment of urinary bladder stones. His skills as an artist are evident in the anatomical drawings that are produced from copper plates in his books. Scarpa was a friend of the Corti family and as a medical student in Pavia, Corti was greatly influenced by Scarpa’s anatomical studies. After graduating in medicine from Vienna, Corti studied histology under Kölliker in Würzburg where he worked on the retina before turning to the cochlea (Kley, 1986). There are many named features of structures in the inner ear (Mudry, 2001) but that of Corti stands out. His involvement in auditory research was restricted to the early 1850s, but its impact was immense. Corti (1850) published a brief report of his research which included some illustrations; the article following his in Archiv für Anatomie, Physiologie und wissenschaftliche Medicin was by Helmholtz (1850) on measures of the speed of nerve transmission. Corti’s (1851) longer article in the following year gave the coloured illustration with which his portrait is combined in Figure 4. Corti distinguished between the inner and outer hair cells on the basilar membrane as well as many other anatomical features of the inner ear, and Helmholtz (1865) drew extensively on Corti’s studies to develop his resonance theory of hearing.
The nerve fibres supplying the inner and outer hair cells were traced by Schultze (1858) but it was his colleague in Bonn, O. Deiters (1859, 1860), who explored the structures of the inner ear in more detail (Figure 4). He indicated how the inner and outer hair cells were arranged in arcs and he also described the supporting cells for the hair cells, with which his name is associated (see V. S. Deiters & Guillery, 2013). Retzius (1884) provided elegant illustrations of the cell structures on the organ of Corti (Figure 4). Like many anatomists before him, his illustrations were initially produced by an artist but dissatisfaction with this procedure resulted in him making his own drawings. Retzius spent most of his academic life at the Karolinska Institute in Stockholm (Grant, 2011).
The inner ear also comprises three semicircular canals and two otolith organs (utricle and saccule) the gross anatomies of which were illustrated by Scarpa (1789) and Retzius (1884). The nonacoustic parts of the inner ear (semicircular canals and otolith organs) have been neglected relative to the cochlear because their function remained unknown long after the role of the cochlear in audition had been appreciated. Indeed, the orthogonal semicircular canals were thought to be involved in auditory localization until their link to vertigo was established (see Wade, 2000). The semicircular canals serve to detect angular accelerations of the head and the otolith organs register the orientation of the head to gravity (a linear acceleration). Thus the inclusion of the vestibular system in the theatre of the senses came rather late. The anatomy of the semicircular canals was well established (see van de Water, 2012) before the function they served was clarified independently by Mach, Breuer, and Crum Brown in the 1870s. The accelerations are detected by bending hair cells in the ampullae of the semicircular canals or beneath the otolith layers of the utricle and saccule. These were elegantly illustrated by Held (1909; Figure 4) together with many representations of cells in the organ of Corti. In the illustration of the hair cells from the saccule of a pigeon (Held, 1909, Figure 33) the epithelial surface is referred to as the cupula and the otoliths are not represented. The distinction between two types of hair cell was to await the examination of the inner ear by means of electronmicroscopes (Wersäll, 1956); he called the flask-shaped hair cells Type I and the cylindrical cells Type II (Figure 4). Wersäll worked in the Karolinska Institute where Retzius had conducted his histological investigation of the inner ear.
Tongue
Malpighi (1665, Figure 5) described and illustrated the papillae on the tongue. In his illustration, he divided the tongue into five sections and noted the distribution of the papillae, particularly in the posterior section. His diagram of the microscopic features of the tongue is inset at the upper left. Waller (1847, Figure 5) examined the tongue of frog and distinguished between two types of papillae. He carried out similar studies on human tongue and described conical or filiform papillae and fungiform papillae (Waller, 1849). The detailed cellular structure of mammalian taste buds was provided independently by Lovén (1867, 1868) and Schwalbe (1867) although neither of them mentioned Malpighi. Lovén initially presented his account in Swedish and this was acknowledged by Schwalbe (1868). They showed that the papillae contained clusters of cells around their sides. Schwalbe called the them ‘taste cups’ whereas Lovén referred to them as ‘taste bulbs’ or ‘taste onions’. Titchener (1915) called them ‘taste-buds’ or ‘taste-beakers’. Schwalbe was a student of Max Schultze and Lovén was a professor in Stockholm; they are both shown in Figure 5, contained within an illustration based on Lovén’s (1867) representation of a circumvallate papilla taken from Schwalbe’s (1887) text book.
The tastes of Malpighi, Waller, Lovén and Schwalbe by Nicholas Wade. Upper left: Marcello Malpighi (1628–1694) is shown with his illustrations of the papillae on the tongue (derived from Malpighi, 1665). Upper right: Augustus Waller (1816–1870) is shown within his illustration of a fungiform papilla from a frog (derived from Waller, 1847). Lower, the diagram of a circumvallate papilla is from Schwalbe (1887) and it is based on an earlier engraving by Lovén (1867). Christian Lovén (1835–1904) is embedded on the left and Gustav Schwalbe (1844–1916) is on the right.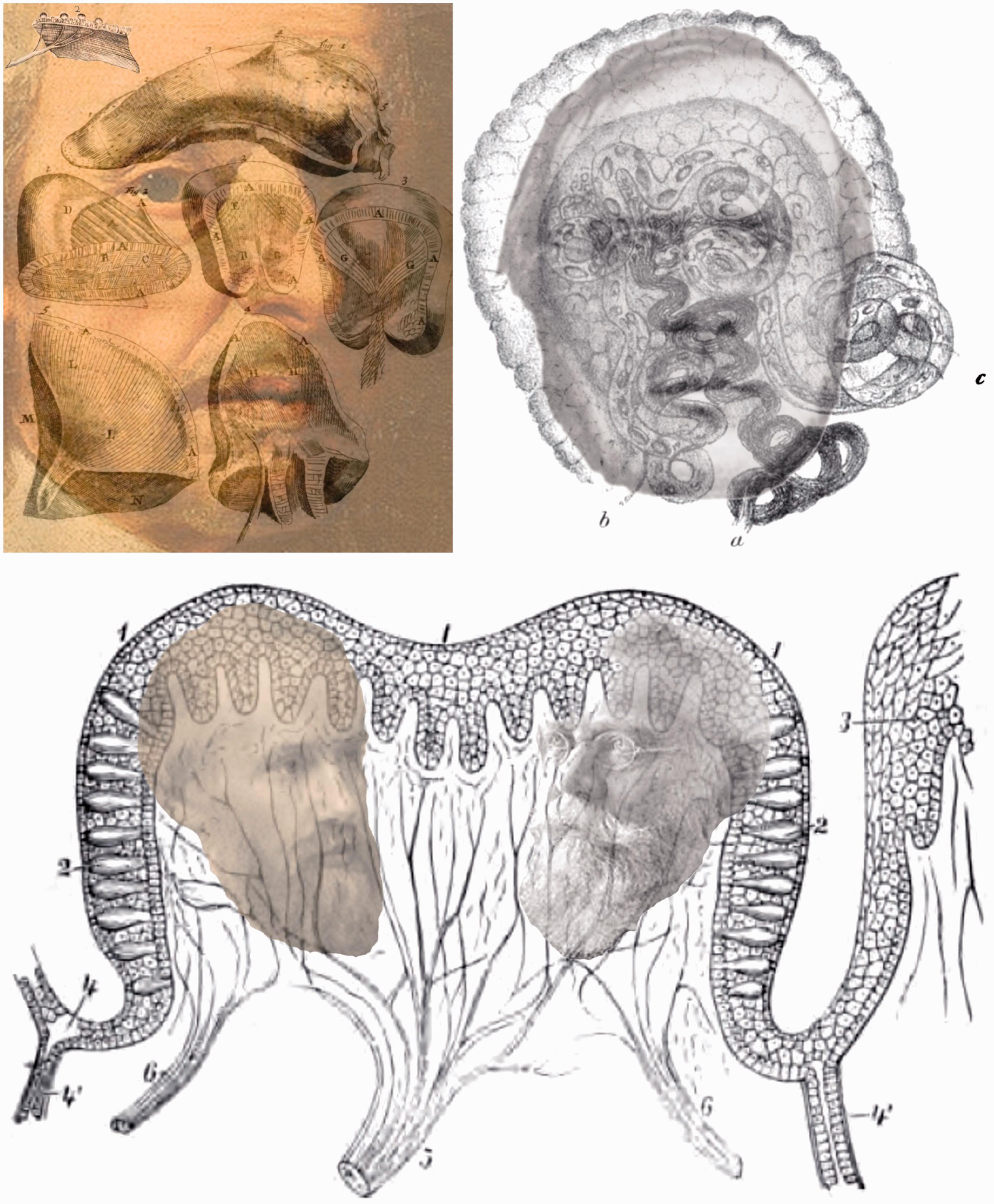
Nose
The olfactory bulb was clearly described and illustrated by Scarpa (1789) but the detail of the olfactory receptors awaited microscopic investigations (Doty, 2015). The observations and illustrations by Eckhard (1855), Ecker (1855, 1857) and Schultze (1856, 1862) did much to clarify the structure of olfactory receptors and the pathways of nerves from them (Figure 6). Eckhard was an anatomist and physiologist at Giessen where he examined the olfactory cells in frogs distinguishing between two types, cylindrical and fusiform. Ecker was an anatomist at Freiburg and studied cells in the olfactory systems of frogs and humans. Both Eckhard (1862) and Ecker (1864, 1869) wrote textbooks on anatomy. Schultze examined the sensory cells within the nose before he investigated those in the eye and his research on olfactory receptors is chronicled by Zippel (1993).
The olfactory cilia of Ecker, Eckhard and Schultze by Nicholas Wade. Left: A portrait of Alexander Ecker (1816–1887) is combined with a diagram of epithelial and ciliary olfactory cells of the frog (derived from Ecker, 1864). Centre: Conrad Eckhard (1822–1905) is shown with olfactory cells of humans (derived from Eckhard, 1862). Right: Max Schultze (1825–1874) with the olfactory cells from owl, pike, frog and human (derived from Schultze, 1856).
Skin, Muscles and Joints
In contrast to the restricted range of receptors in the eye, ear, mouth and nose, the skin provided histologists with a startling variety of specialised cells. In chronological sequence, they were described by Pacini (1840), Meissner (1853), Krause (1860), Merkel (1875), Golgi (1880) and Ruffini (1894) and their names continue to be associated with the receptor cells. The anatomists are shown in Figure 7 together with representations of the cells bearing their names.
Explorers of the skin by Nicholas Wade. From left to right, Filippo Pacini (1812–1883), Georg Meissner (1829–1905), Wilhelm Krause (1833–1910), Friedrich Merkel (1845–1919), Camillo Golgi (1843–1926) and Angelo Ruffini (1864–1929) combined with illustrations of their eponymous cells.
Pacini first observed the onion-shaped cells that bear his name in 1831 during a dissection class as a medical student (Bentivoglio & Pacini, 1995; Henle & Kölliker, 1844) but he did not publish an account of them until 1840 and his illustration was reprinted in his 1889 study. The cells are large and had been seen as early as 1741 by Abraham Vater in the skin of human fingertips. He called them ‘papillae nerveae’ and they were examined in greater detail by his student Johann Gottlob Lehmann in the same year (see Vater, 1747); they are also referred to as Vater–Pacinian bodies (Neumeister, 1845). Meissner initially described his microscopic observations of human skin together with his professor at Göttingen (Wagner & Meissner, 1852) and then published them in his own name the following year; they are occasionally called Meissner–Wagner corpuscles (Nafe, 1934). The touch corpuscles were considered to respond to deep pressure. Krause’s (1861) end-bulbs or corpuscles were described in 1860 and his portrait is combined with an illustration of a touch cell from the conjunctiva of a human eye. Merkel (1875) illustrated a variety of specialized skin receptors including the ‘touch cell’ with which his name became attached 3 years later. His atlas of the skin and sense organs (Merkel, 1917) contained the schematic diagram of a Merkel cell from a human lip that was used in Figure 7. Golgi (1880) observed two types of cells in the region of the tendon. One was somewhat like the cells described by Pacini and Krause which Golgi referred to it as a ‘tactile body’. Mazzoni (1890) showed that they are present in the skin and they are now referred to Golgi–Mazzoni corpuscles. The portrait of Golgi is combined with an illustration of the cells from Merkel (1917). Ruffini is shown in a duplicate and reflection of the representation of cylindrical cells in his 1894 study; the publication also drew attention to the Golgi–Mazzoni subcutaneous receptors.
Cutaneous sensory ‘spots’ specifically responsive to touch (pressure) and pain, as well as warmth and cold, had been described by Ritter (1801) and were isolated later in the century, using more sensitive and specific apparatus. A division of the skin senses into three separate systems (one to register temperature, a second for pressure and a third for touch) was proposed by Natanson (1844; see Norrsell, Finger, and Lajonchere, 1999). Three sets of independent studies were reported in the 1880s: Blix (1882, 1884) followed by Goldscheider (1884) and Donaldson (1885). All three argued for a separate temperature sense and this was implicitly supported by von Frey (1894) who linked the sensations of touch, warmth, cold and pain to specific skin receptors. He proposed that Meissner corpuscles were involved in touch perception, Ruffini cylinders for warmth, Krause end-bulbs for cold and free nerve endings for pain (see Melzack & Wall, 1962). His theory survived despite mounting evidence against it (Sinclair, 1967). Blix, Goldscheider and von Frey are shown in Figure 8.
Cutaneous sensory spots of Blix, Goldscheider and von Frey by Nicholas Wade. Left: Magnus Blix (1849–1904) is shown in mappings of sensory spots for touch (black), cold (green) and warm (red) on the back of the hand; the patterns are reproduced from Blix (1882). Centre: Alfred Goldscheider (1858–1935) can be seen in an illustration of cold (blue) and warm (red) sensitive spots from the fingertips (from Goldscheider, 1885). Right, a portrait of Max von Frey (1852–1932) is combined with a pattern pressure spots from hairy and hairless areas of the human calf (from Von Frey, 1896).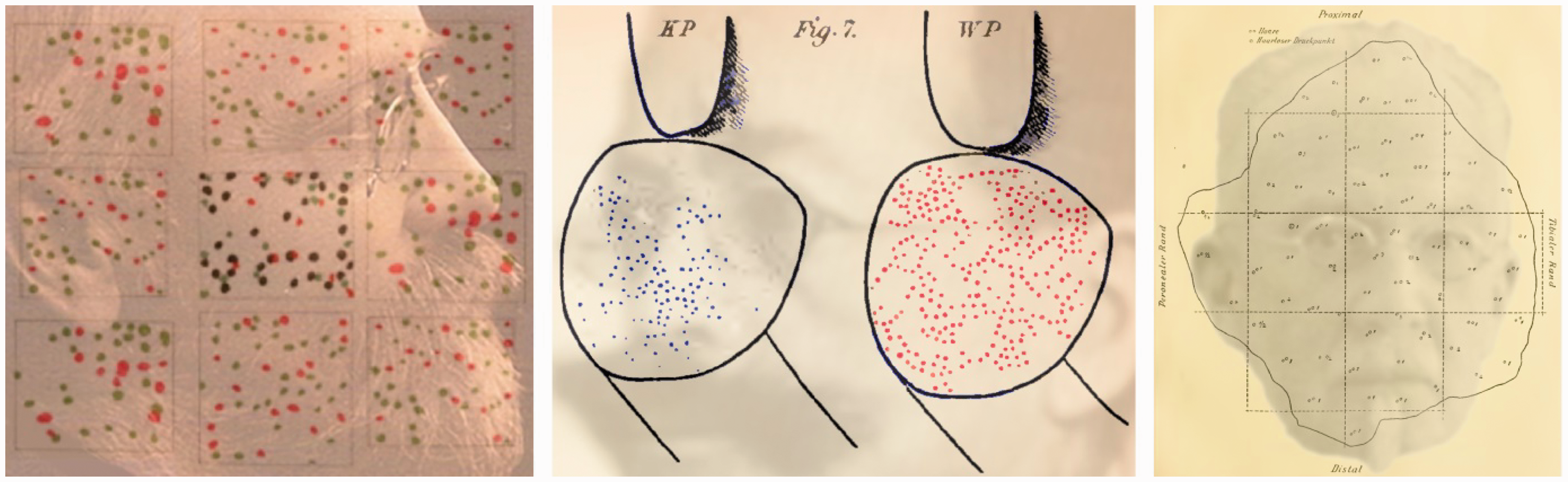
Touch and kinaesthesis are often considered in combination (see Rose & Mountcastle, 1959): Coordinated behaviour is based upon the integration of sensory information from the surface and beneath the skin. The principal receptors were considered to be in the muscles or their tendons which respond to the stretching of muscles or the tension in tendons as well as in the joint capsules. Hassall (1851) illustrated muscle spindles but they were described in greater detail by Ruffini (1898) who distinguished between three different types. Specialised receptors were also found in the tendons by Golgi (1880) and they continue to be known as Golgi tendon organs. The joint receptors are considered to be of four types similar to those which have been mentioned earlier: free nerve endings, Golgi endings, Ruffini cylinders and Pacinian corpuscles. The muscle and tendon receptors are shown in Figure 9 together with their discoverers.
Kinaesthetes by Nicholas Wade. Left: Arthur Hill Hassall (1817–1894) in an illustration of muscle spindles in the tongue (derived from Hassall, 1851). Centre: Angelo Ruffini in his representation of muscle spindles (Ruffini, 1898). Right: Camillo Golgi combined with his illustration of nerves supplying tendon organs (derived from Golgi, 1880).
Hassall (1849), a general practitioner from London, published one of the first books devoted to microscopic anatomy; it consisted of two volumes that were lavishly illustrated with many of the figures in colour. It was translated into German (Hassall, 1852) and an American edition was published in 1851. Additional plates of illustrations were produced for the latter and it was one of these that contained the initial indications of muscle spindles; the German translation did not contain it. Ruffini (1898) acknowledged that Hassall’s (1851) ‘description and interpretation, although brief, show clearly he had before him “muscle-spindles”’ (p. 190). Weismann (1861) provided more detail of the organization of the muscle spindles within the ‘Weismann bundles’ and Kühne (1863) gave them the name ‘muscle spindles’. When Golgi (1880) investigated the receptors in the tendons that bear his name he was able to use his revolutionary silver staining method (see Golgi, 1873; Mazzarello, 2010; Wade & Piccolino, 2006).
Conclusions
The scientists portrayed in this early history of receptors were visionaries in two senses. They used their observational skills to detect and describe the receptors in a variety of senses. They were also visionaries because they encapsulated their discoveries with schematic diagrams that could be appreciated with a greater precision than their written descriptions. Some, like Scarpa, Retzius and Golgi, displayed their artistry in the representations they published. Moreover, most of the discoveries were made before staining methods assisted in isolating the neural structures when observed under the microscope.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.