
Abstract
Deltamethrin (DEL) and thiacloprid (THIA) are commonly used synthetic insecticides in agriculture either separately or in combination. There is limited information in human cells for the effects of the mixture of DEL + THIA on oxidative stress. Therefore, the present study was designed to examine the effects of the mixture on cell proliferation and oxidative stress in human lung fibroblast cells. Human telomerase reverse transcriptase (hTERT)-expressing human lung fibroblasts, WTHBF-6 cells, were treated with 2.5 + 37.5, 5 + 75, 12.5 + 187.5, and 25 +375 µM concentrations of DEL + THIA for the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide assay and 5 + 75, 12.5 + 187.5, and 25 + 375 µM for lipid peroxidation and reduced glutathione (GSH) assays for 24, 48, and 72 h in the absence and presence of metabolizing fractions of the mammalian liver (S9 mixture). Both the mixture of DEL + THIA and their metabolites significantly reduced cell viability and induced cytotoxicity in WTHBF-6 cells, especially at higher concentrations. The mixture of DEL + THIA significantly decreased GSH levels at the highest concentration for all treatment times and at the highest two concentrations (12.5 + 187.5 and 25 + 375 µM) for 72 h in the presence of S9 mixture. The highest concentration of DEL + THIA mixture caused a significant increase in malondialdehyde (MDA) level at 72 h in the absence of S9 mixture. There were also significant increases in MDA levels at the highest concentration for 48-h and all concentrations of DEL + THIA for 72-h treatment in WTHBF-6 cell cultures with S9. These data showed that the mixture of DEL + THIA and their metabolites can induce cytotoxicity and oxidative stress in human lung fibroblasts.
Introduction
Pyrethroid and neonicotinoid insecticides are widely used in controlling insect pests on major crops and in animal health because they have high activity against insects (Şekeroğlu et al., 2013, 2014). Occupationally or incidentally, humans are exposed not only to single pesticides but also to pesticide mixtures (Şekeroğlu et al., 2014). Deltamethrin (DEL) is an alpha-cyano pyrethroid insecticide with strong, neurotoxic insecticidal activity. Thiacloprid (THIA), a member of the neonicotinoid group of insecticides, also shows neurotoxic activity acting as an agonist of the nicotinic acetylcholine receptor (Aydın, 2011; Şekeroğlu et al., 2013, 2014). They are widely used separately or as a mixture for the control of insect pests (Şekeroğlu et al., 2013, 2014). Proteus, a commercial mixture of DEL + THIA, is a powerful and wide-spectrum insecticide (Şekeroğlu et al., 2013). It is used for a wide variety of crops such as potato, cabbage, pea, cauliflower, many vegetables (Bayer Crop Science, 2020a), hazelnut, peach, apple, olive, melon (Bayer Crop Science, 2020b), and onion (Bayer Crop Science, 2020c). The mixture is mixed with water and sprayed to cover all parts of the plants (Bayer Crop Science, 2020c). Therefore, people can be exposed to it by inhalation.
Pesticides may show toxicity due to the induction of oxidative stress because some pesticides can increase reactive oxygen species (ROS) levels and lipid peroxidation (LPO), alter homeostatic and regulatory processes, or reduce antioxidant defense (Aydın, 2011; Lushchak et al., 2018; Öğüt et al., 2011; Saoudi et al., 2019). If there is an increase in ROS levels, they may react with important cellular macromolecules and cause enzyme inactivation, LPO, DNA damage, and genotoxicity (Lushchak et al., 2018). Previous studies have reported that exposure to combinations of some pesticides can cause more genotoxic effects than exposure to a single pesticide (Şekeroğlu et al., 2013). Some pesticides can adversely affect the function of various organs and lead to the development of organ toxicity, immunotoxicity, and carcinogenicity (Aydın, 2011; Şekeroğlu et al., 2013, 2014).
Pesticides in combinations have shown cumulative and/or synergistic effects (Şekeroğlu et al., 2013). Since some pesticides in mixtures can interact with each other, living organisms, and the environment in multiple ways, exposure to them can cause problems for the environment, ecosystems, and human health. Although there are many studies showing that DEL or THIA can induce cytotoxicity and oxidative stress, toxicity studies of the mixture of DEL + THIA are very limited. A literature review has shown that there are only two studies about the effects of DEL + THIA mixture on cytotoxicity and oxidative stress. The mixture of DEL + THIA caused oxidative stress by decreasing the levels of antioxidant enzymes in lymphoid organs and plasma in rats (Aydın, 2011). It has also been reported that the mixture of DEL + THIA can synergistically induce cytotoxicity and genotoxicity in rat bone marrow cells (Şekeroğlu et al., 2013).
It has been reported that exposure to some pesticides may increase the risk of lung cancer among licensed pesticide applicators (Bonner et al., 2017). There is evidence that exposure to various pesticides can disturb a number of biological pathways, including oxidative stress and immunotoxicity that have been linked with carcinogenesis (Alavanja and Bonner, 2018). However, the potential effects of DEL + THIA mixture, which can also be taken into the body by inhalation, on cell viability and oxidative stress in human lungs have not been studied. Therefore, the present study aimed to investigate the effect of DEL + THIA mixture on cytotoxicity and oxidative stress in human lung cells in the absence and presence of metabolizing fractions of the mammalian liver (S9 mixture).
Tumors consist of malignant cancer cells and stromal cells that constitute the tumor microenvironment. The tumor stroma mainly consists of the basement membrane, fibroblasts, extracellular matrix, and immune cells. Cancer-associated fibroblasts (CAFs) are one of the major components in the tumor stroma, and their main precursors are normal fibroblasts (Bremnes et al., 2011; Wang et al., 2017). ROS produced by tumor epithelial cells or stromal cells such as fibroblasts can contribute to tumor growth and metastasis (Costa et al., 2014). Although recent studies indicated that CAFs are involved in tumor growth, little is known about their role in lung cancer (Bremnes et al., 2011; Wang et al., 2017). Therefore, here we used human lung fibroblasts (WTHBF-6 cells) with an extended life span and characteristics of normal diploid cells similar to primary human bronchial fibroblasts.
Reconstituting telomerase activity in a variety of cell types by expression of hTERT offers the possibility to discover the normal properties of primary human cells without causing genetic instability and confounding effects of cellular senescence. These immortal cells are more valuable to the study of the normal functioning of various processes during cancer development (Lee et al., 2004; Wise et al., 2004). Thus, hTERT-expressing human lung fibroblasts were selected for our experiments.
Materials and methods
Chemicals
Dulbecco’s minimal essential medium and Ham’s F-12 (DMEM/F12), phosphate-buffered saline, penicillin–streptomycin, trypsin/ethylenediaminetetraacetic acid (EDTA), sodium pyruvate, and
Cell culture
Human telomerase reverse transcriptase (hTERT)-expressing human lung fibroblasts, WTHBF-6 cells, were a kind gift from Dr John P Wise (University of Louisville, Louisville, Kentucky, USA). This cell line exhibits a diploid karyotype, normal growth parameters, and an extended life span (Xie et al., 2015). The cells were cultured in DMEM/F12 medium supplemented with 15% Cosmic calf serum, 2 mM
Cell viability assay and concentration selection
Since a commercial mixture of DEL + THIA (Proteus) contains DEL 20 g L−1 and THIA 150 g L−1, the concentrations were prepared as THIA at seven and a half times the concentration of DEL in all experiments. DEL and THIA concentrations used in the present study were chosen as the result of a preliminary study using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay. WTHBF-6 cells were seeded in a 96-well plate (1 × 104 cells each well). The cells were treated with 10 different concentrations of the mixtures of DEL + THIA (0.5 + 7.5, 1.25 + 18.75, 2.5 + 37.5, 5 + 75, 12.5 + 187.5, 25 + 375, 50 + 750, 100 + 1500, 150 + 2250, 200 + 3000 µM) dissolved in dimethyl sulfoxide (DMSO; 0.1%) for 24, 48, and 72 h. Cells treated with DMSO (0.1%) were used as control. Cultures were incubated for 4 h after 20 µL of MTT solution was added to each well. Then, 100 µL of DMSO was added to each well, followed by measuring optical density at 570 nm using a microplate reader (Biotek Elx800, Winooski, Vermont, USA). Logarithmic regressions were performed to determine the half maximal inhibitory concentration (IC50). All experiments were performed in triplicate, and the results were expressed as percentage inhibition relative to control cells.
According to the preliminary findings, DEL + THA reduced the cell viability values to <IC50 value after 150 + 250 µM, 50 + 750 µM, and 25+375 µM concentrations for 24-h, 72-h, and 120-h treatments, respectively. We used 2.5 + 37.5, 5 + 75, 12.5 + 187.5, and 25+375 µM concentrations for further MTT assays and 5 + 75, 12.5 + 187.5, and 25 + 375 µM for malondialdehyde (MDA) and reduced glutathione (GSH) assays in the absence and presence of S9 mixture.
LPO and GSH assays
WTHBF-6 cells were incubated with DEL + THIA for 24, 48, and 72 h. Control cells were also treated with DMSO (0.1%). After treatment, the media were then removed, and cells were washed three times with 3 mL ice-cold phosphate-buffered saline. Cells were treated with cold lysis buffer (50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% Triton X, 5 mM disodium EDTA) on ice for 10 min. The cells were scraped off and the cell suspension was transferred into a centrifuge tube on ice. The cell suspensions were centrifuged at 15,000 × g for 10 min at 4°C. Supernatants containing cell extracts were used for the estimation of both LPO and GSH contents.
LPO was quantified as MDA according to the method of Ohkawa et al. (1979). Cell extract (0.1 mL) and 1.9 mL of 0.1 M sodium phosphate buffer (pH 7.4) were mixed and incubated in a water bath at 37°C for 1 h, followed by the addition of 5% trichloroacetic acid (TCA) and 1% thiobarbituric acid. The solution was heated in a water bath at 95°C for 60 min, and absorbance was measured at 532 nm. MDA concentration was calculated using an extinction coefficient of 1.56 × 105 M−1 cm−1 and expressed in nmoles of MDA per hour per milligram protein.
GSH content was determined by the method of Ellman (1959). The mixture of 0.1 mL cell extract and 0.9 mL of 5% TCA was centrifuged at 2300 × g for 15 min at 4°C. Then 1.5 mL of 0.01% DTNB and 0.5 mL of supernatant were mixed, and absorbance was measured at 412 nm. GSH concentration was calculated using the molar extinction coefficient of 1.36 × 104 M−1 cm−1 and expressed in nmoles of GSH per milligram protein.
All experiments were performed in the absence or presence of S9 mixture. Human liver S9 microsomal fraction and an NADPH generation system were used for metabolic activation. The S9 mixture was prepared in 1× Tris buffer, pH 7.6. The final concentration of the S9 mixture in the culture medium was 1%.
Statistical analysis
Each experiment was repeated three times. All data were expressed as the mean ± standard error (SE). Statistical analyses were performed using SPSS version 14.0 (SPSS Inc., Chicago, Illinois). The data were analyzed by analysis of variance, followed by paired Student’s t-test. The value of p < 0.05 was considered statistically significant.
Results
Cell viability
The effects of DEL + THIA mixture on cell viability in WTHBF-6 cells are shown in Figure 1. MTT results showed that DEL + THIA significantly reduced the cell viability in a time- and concentration-dependent manner (p < 0.05). The mixture of DEL + THIA has greater cytotoxic effects on WTHBF-6 cells, especially at higher concentrations. These decreases in cell viability were significant at 100 + 1500 µM, 150 + 2250 µM, and 200 + 3000 µM for 48 h and 72 h and 50 + 750 µM for 72 h when compared to the control (p < 0.05).
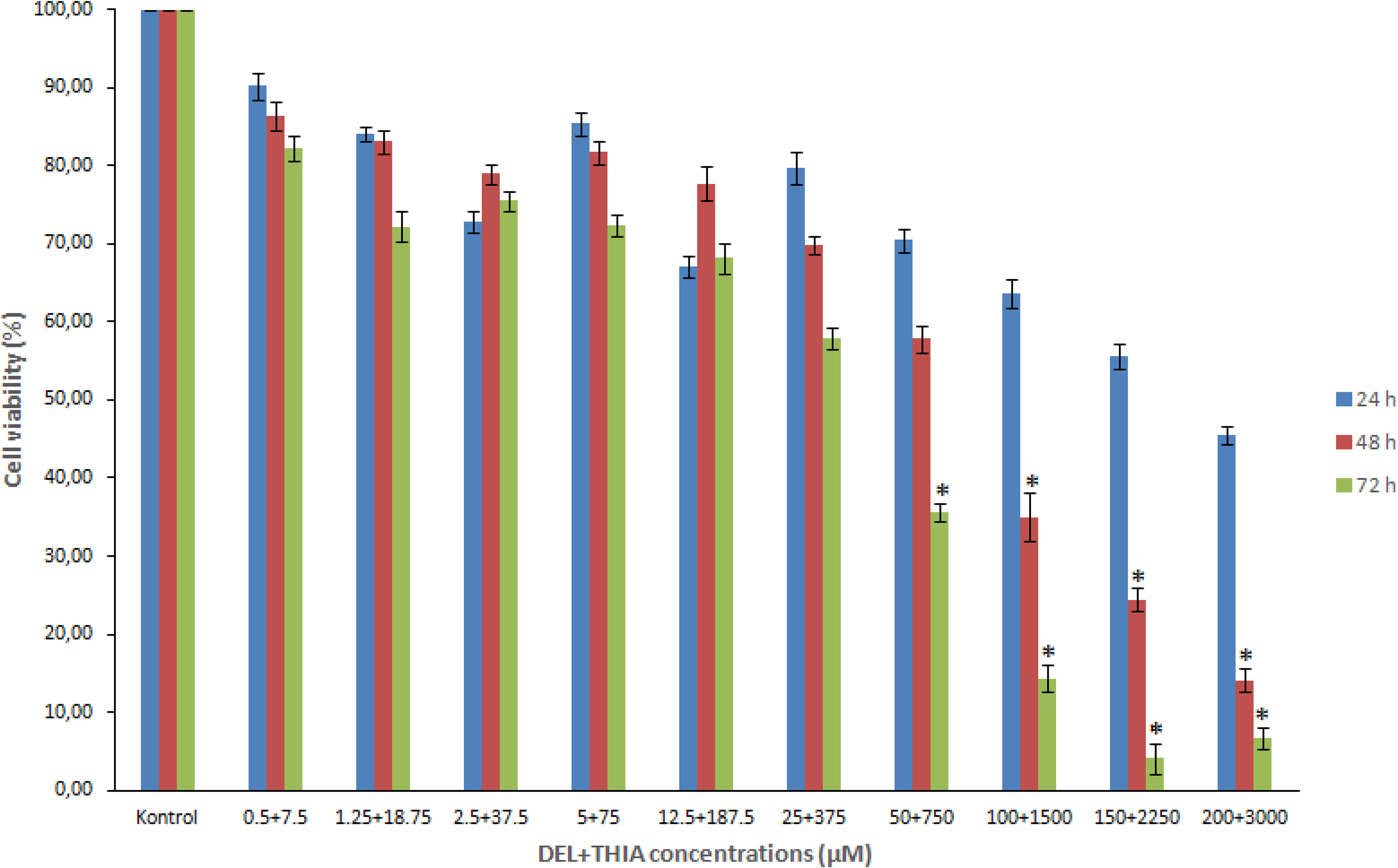
Effects of DEL + THIA on cell viability of WTHBF-6 cells. DEL + THIA significantly reduced the cell viability after 100 + 1500 µM for 48 h and 50 + 750 µM for 72 h. Data are expressed as mean ± SE for n = 3. *Significant against control (DMSO 0.1%) at p < 0.05. DEL: deltamethrin; THIA: thiacloprid; SE: standard error..
DEL + THIA had no effect on cell viability for 24 h in the absence or presence of S9 (Figure 2(a)). Concentration-dependent decreases were observed in cultures treated with DEL + THIA for 48 and 72 h in the absence and presence of S9 mixture (p < 0.05). Cell viability was maximally decreased at the highest two concentrations (12.5 + 187.5 and 25 + 375 µM) in the absence of S9 mixture, but only at the highest concentration (25 + 375 µM) in the presence of S9 mixture for the 48-h treatment (p < 0.05) (Figure 2(b)). Although decreases in cell viability were significant at all concentrations except for the lowest concentration for 72-h treatment in culture without S9 mixture, all DEL + THIA concentrations reduced cell viability in cultures with S9 mixture for 72-h treatment (p < 0.05) (Figure 2(c)). These results showed that both the mixture of DEL + THIA and their metabolites can significantly reduce the cell viability and induce cytotoxicity in human lung fibroblasts, especially at higher concentrations.
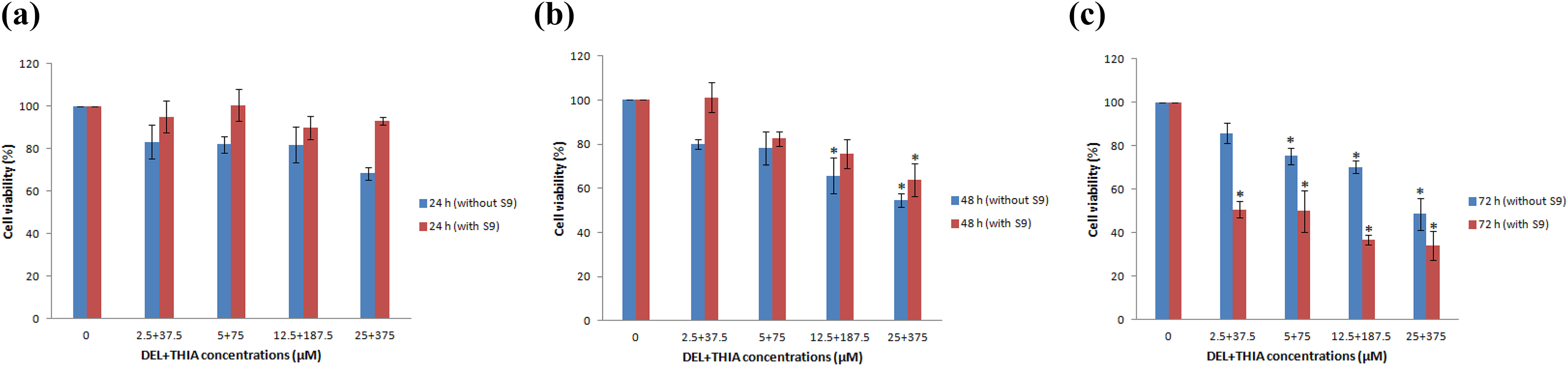
Effects of DEL + THIA tested four concentrations on cell viability of WTHBF-6 cells after 24-, 72- and 72-h treatments in the absence and presence of S9 mixture. DEL + THIA did not significantly reduce the cell viability for 24 h in the presence of S9 mixture (a). Cell viability significantly decreased at the highest two concentrations (12.5 + 187.5 and 25 + 375 µM) in the absence of S9 mixture, but only at the highest concentration (25 + 375 µM) in the presence of S9 mixture for 48-h treatment (b). All DEL + THIA concentrations significantly reduced the cell viability for 72-h treatment except for the lowest concentration in culture without S9 mixture (c). Data are expressed as mean ± SE for n= 3. *Significant against control (DMSO 0.1%) at p < 0.05. DEL: deltamethrin; THIA: thiacloprid; SE: standard error.
LPO and GSH
The DEL + THIA mixture decreased GSH levels at the highest concentration (25 + 375 µM) for 48 and 72 h in the absence of S9 mixture compared to the control group (Figure 3(a)). However, the mixture decreased GSH levels at the highest concentration for all treatment times and the highest two concentrations (12.5 + 187.5 and 25 + 375 µM) for 72 h in the presence of S9 mixture compared to the control group (Figure 3(b)). A significant increase in MDA level was found at the highest concentration of DEL + THIA mixture for 72-h treatment in cultures without S9 mixture compared to the control group (Figure 4(a)). There were significant increases in MDA levels at the highest concentration for 48 h and all concentrations of DEL + THIA for 72-h treatment in cultures with S9 mixture compared to the control group (Figure 4(b)).
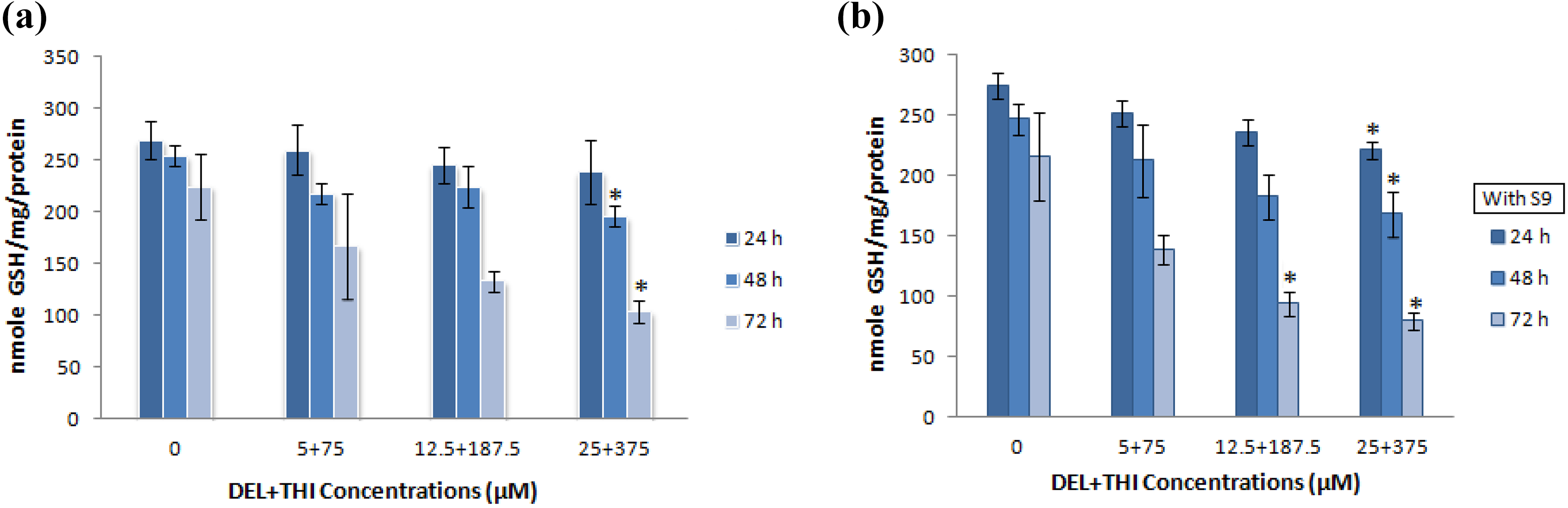
Effects of DEL + THIA on GSH levels in WTHBF-6 cells after 24-, 72- and 72-h treatments in the absence and presence of S9 mixture. The mixture of DEL +THIA significantly decreased GSH levels at the highest concentration (25 + 375 µM) for 48 and 72 h in the absence of S9 mixture (a). Significantly decreased GSH levels were found at the highest concentration for all treatment times and at the highest two concentrations (12.5 + 187.5 and 25 + 375 µM) for 72 h in the presence of S9 mixture (b). Data are expressed as mean ± SE for n = 3. *Significant against control (DMSO 0.1%) at p < 0.05. DEL: deltamethrin; THIA: thiacloprid; GSH: reduced glutathione; SE: standard error.
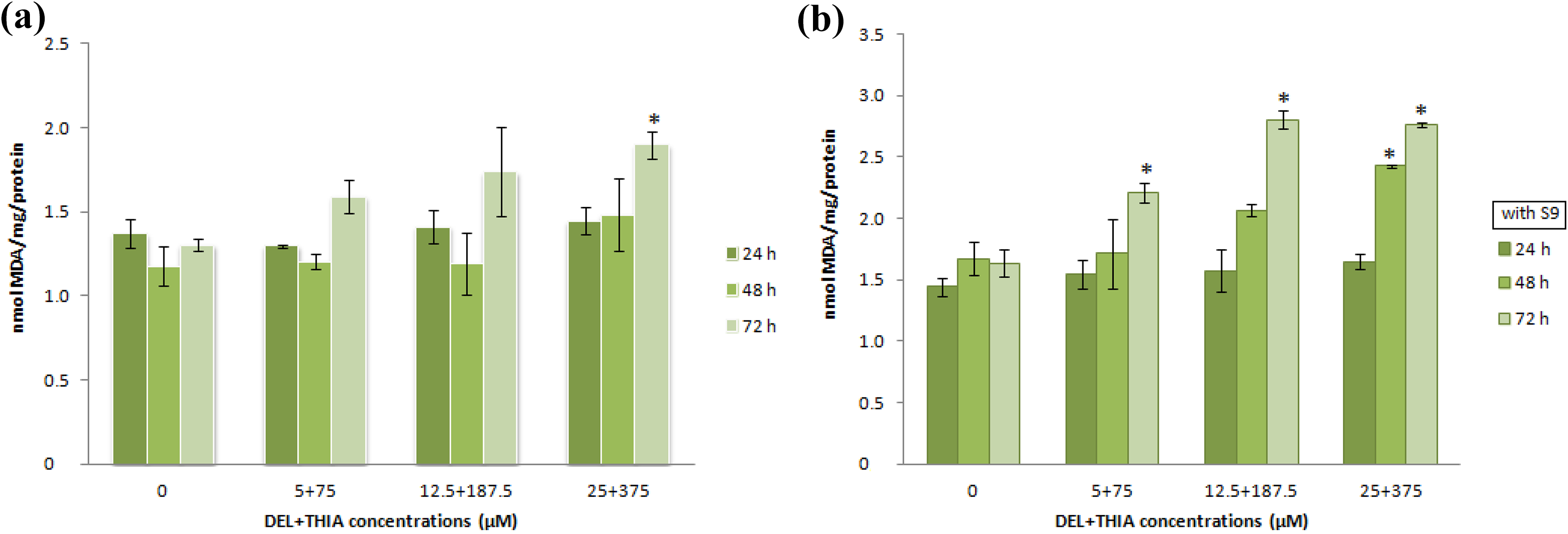
Effects of DEL + THIA on MDA levels in WTHBF-6 cells after 24-, 72- and 72-h treatments in the absence and presence of S9 mixture. A significant increase in MDA level was found at the highest concentration of DEL + THIA mixture for 72-h treatment in cultures without S9 mixture (a). There were significant increases in MDA levels at the highest concentration for 48 h and all concentrations for 72-h treatment in cultures with S9 mixture (b). Data are expressed as mean ± SE for n = 3. *Significant against control (DMSO 0.1%) at p < 0.05. DEL: deltamethrin; THIA: thiacloprid; MDA: malondialdehyde; SE: standard error.
Discussion
The effects of DEL on cytotoxicity and oxidative stress have been demonstrated in previous studies. Decreased proliferation was found in mouse fibroblast cells treated with a commercial formulation of DEL (50 and 500 µM) for 72 h (Baeza-Squiban et al., 1987). Significantly increased MDA levels were found in male rats treated with DEL (1 mg kg−1day−1 for 21 days) (El-Gohary et al., 1999). DEL decreased the activity of antioxidant enzymes and increased thiobarbiturate levels (TBARSs) in mouse liver exposed to 5.6 mg kg−1 day−1 for 15 days (Rehman et al., 2006) and rat liver exposed to 15 mg kg−1 day−1 for 30 days (Manna et al., 2005). DEL (1.28 mg kg− day−1) significantly induced LPO by increasing TBARS in rat plasma (Yousef et al., 2006). Significantly increased plasma MDA concentrations were found in female rats treated with DEL (0.003, 0.03, and 0.3 mg kg−1 day−1) for 45 days (Chargui et al., 2012). DEL significantly decreased the cell viability at 10–1000 μM (Romero et al., 2012) and 50–250 μM (Ko et al., 2016) in human neuroblastoma cells. DEL (15, 30, and 60 mg kg−1 day−1) induced oxidative stress by decreasing glutathione S-transferase (GST) and increasing MDA levels in rat liver for 30 days treatment (Çavuşoğlu et al., 2016). Increased oxidative stress activities were found in rats treated with DEL (7.2 mg kg−1 day−1) for 30 days (Ncir et al., 2015). Decreased cell viability, ROS, and GSH levels in spleen and thymus were observed in mice treated with a single dose of DEL (5 mg kg−1, oral for 7 days) (Kumar et al., 2018). DEL (2.56 mg kg−1 day−1) for 28 days increased LPO and decreased GSH in rats (Mekircha et al., 2018).
Some studies have also shown that THIA can induce cytotoxicity and oxidative stress. Increased cytotoxicity (2.0 × 10−1 M) and DNA damage (0.6 × 10−1 to 1.4 × 10−1) were observed in human peripheral blood lymphocytes exposed to Calypso, a commercial formulation of THIA (Calderón-Segura et al., 2012). THIA (120–480 µg mL−1) reduced the proliferation index in cultured bovine peripheral lymphocytes (Drazovska et al., 2013). THIA (75, 150, and 300 µg mL−1) induced cytotoxicity in human peripheral blood lymphocytes in the absence and presence of S9 mixture (Kocaman et al., 2014). A decrease in the expression of GST was observed in bovine blood cultures treated with THIA (30–480 µg mL−1) for 24 h (Galdíkova et al., 2015). There was a significant decrease in GST and an increase in MDA activity in rat liver treated with THIA (22.5 mg kg−1 day−1) for 30 days (Hendawi et al., 2016). THIA (30, 60, 120, 240 and 480 µg mL−1) induced cytotoxic/cytostatic effects in bovine blood cultures (Galdíkova et al., 2019). Increases in LPO and protein oxidation and decreases in GSH levels were found in THIA-treated (22.5 mg kg−1 day−1) rats for 30 days (Kammoun et al., 2017). THIA (0.5–4 mM) decreased the viability of human neuroblastoma and human hepatocellular carcinoma cells after 24- and 48-h treatments (Şenyıldız et al., 2018). THIA (22.5 mg kg−1) for 30 days induced nephrotoxicity and hepatotoxicity by increasing MDA and decreasing GSH activities in the liver and kidney of rats (Kammoun et al., 2019). Oxidant-based DNA damage and decreased cell viability and proliferation were detected in bovine peripheral lymphocytes treated with THIA (10, 30, 60, 120, and 240 µg mL−1) for 4 h (Schwarzbacherová et al., 2019). Significantly GSH peroxidase (GPx) levels were found in mice brain treated with THIA (50, 100, and 200 mg kg−1) for 3 weeks (Akkoç et al., 2020).
There are only two reports on the toxic effects of the mixture of DEL + THIA. The acute and subacute in vivo toxic effects of DEL or THIA, or a mixture of these insecticides was investigated in plasma, polymorphonuclear leukocytes, and lymphoid organs such as spleen, thymus, and bone marrow of rats treated orally with THIA (single acute dose of 112.5 mg kg−1 and a subacute dose of 22.5 mg kg−1 day−1 for 30 days) or DEL (single acute dose of 15 mg kg−1 and a subacute dose of 3 mg kg−1 day−1 for 30 days), or combined doses of these pesticides. All treatments induced oxidative stress in rats by significantly decreasing GSH levels in all lymphoid organs and increasing LPO in all lymphoid organs and plasma (Aydın, 2011). Similar to these results, the results of the present study also demonstrated that the mixture of DEL + THIA can induce oxidative stress in lung cells by decreasing GSH levels and increasing MDA levels. The treatment of the mixture of DEL + THIA for 24 h or 30 days induced cytotoxicity and genotoxicity in rat bone marrow cells (Şekeroğlu et al., 2013). It has also been reported that the combination of these pesticides showed a synergistic effect in rat bone marrow cells. Consistent with previous results, our in vitro results have shown that the mixture of DEL + THIA can induce cytotoxicity and oxidative stress in human lung cells despite differences in concentrations, cells, organisms, and treatment times. Besides, in vivo toxicity of DEL + THIA mixture may result from oxidative stress.
The results of the present study were also correlated with some previous studies in which the effects of DEL or THIA in combination with other insecticides have been investigated. Increased oxidative damage due to high levels of ROS and LPO was detected in rat liver treated with chlorpyrifos and/or DEL at low (5 mg kg−1 day−1) and high (35 mg kg−1 day−1) doses for 16 weeks (Tuzmen et al., 2008). The mixture of DEL (0.003, 0.03, and 0.3 mg kg−1 day−1) and permethrin (0.013, 0.13, and 1.3 mg kg−1 day−1) for 30, 45, and 60 days decreased superoxide dismutase, GPx, and catalase activities in rats (Chargui et al., 2010). Decreased GST and increased MDA levels were found in rat liver treated with DEL (1.28 mg kg−1 day−1), fluoride (44.5 mg L−1 day−1), and their combination for 28 days (Dubey et al., 2013). DEL (1.02, 2.56, and 6.40 mg kg−1), dichlorvos (0.64, 1.64, and 4.00 mg kg−1), and their mixtures increased MDA and protein carbonyl levels in rat liver for 90 days (Xu et al., 2015). Acute exposure to a combination of DEL and chlorpyrifos (DEL 5 mg kg−1, chlorpyrifos 1 mg kg−1 for 10 days) induced brain oxidative damage by adversely altering LPO and antioxidant parameters (GPx and GSH) in rats (Saoudi et al., 2019). Increased toxicity and oxidative stress were found in zebra fish treated with beta-cypermethrin and THI (Wang et al., 2020). Both our results and the results of previous studies support the hypothesis that some pesticide combinations including DEL or THIA can induce oxidative damage and toxicity.
Chronic oxidative stress has serious effects on tumor initiation, growth, and metastasis. ROS can be produced by tumor epithelial cells or various stromal cell types such as fibroblasts. ROS act as important signaling molecules not only in cancer cells but also in surrounding stromal components, through diffusion of ROS compounds. Ultimately, ROS can mediate cell motility and invasive properties of tumor cells, contribute to extracellular matrix remodeling, enhance neoangiogenesis, and induce metabolic reprogramming of tumor and stromal cells (Costa et al., 2014). Because DEL + THIA mixture increased oxidative damage in human lung fibroblasts in the present study, they may promote tumor development and growth through the abovementioned alterations.
In conclusion, the results of the study demonstrated that DEL + THIA mixture and their metabolites can induce cytotoxicity by decreasing cell viability and oxidative stress by decreasing GSH levels and increasing MDA levels in human lung fibroblasts. Since oxidative stress can play an important role in carcinogenesis and some genetic, metabolic, and cellular responses, more detailed in vivo and in vitro studies should be performed to better understand the potential effects of DEL + THIA mixture in lung cancer progression.
Footnotes
Acknowledgements
We thank Dr John P Wise for the gift of cells.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by a grant from Scientific Research Funding of Ordu University (Turkey) with the project number: AR-1669.